
Genetic Foundation of Male Spur Length and Its Correlation with Female Egg Production in Chickens
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
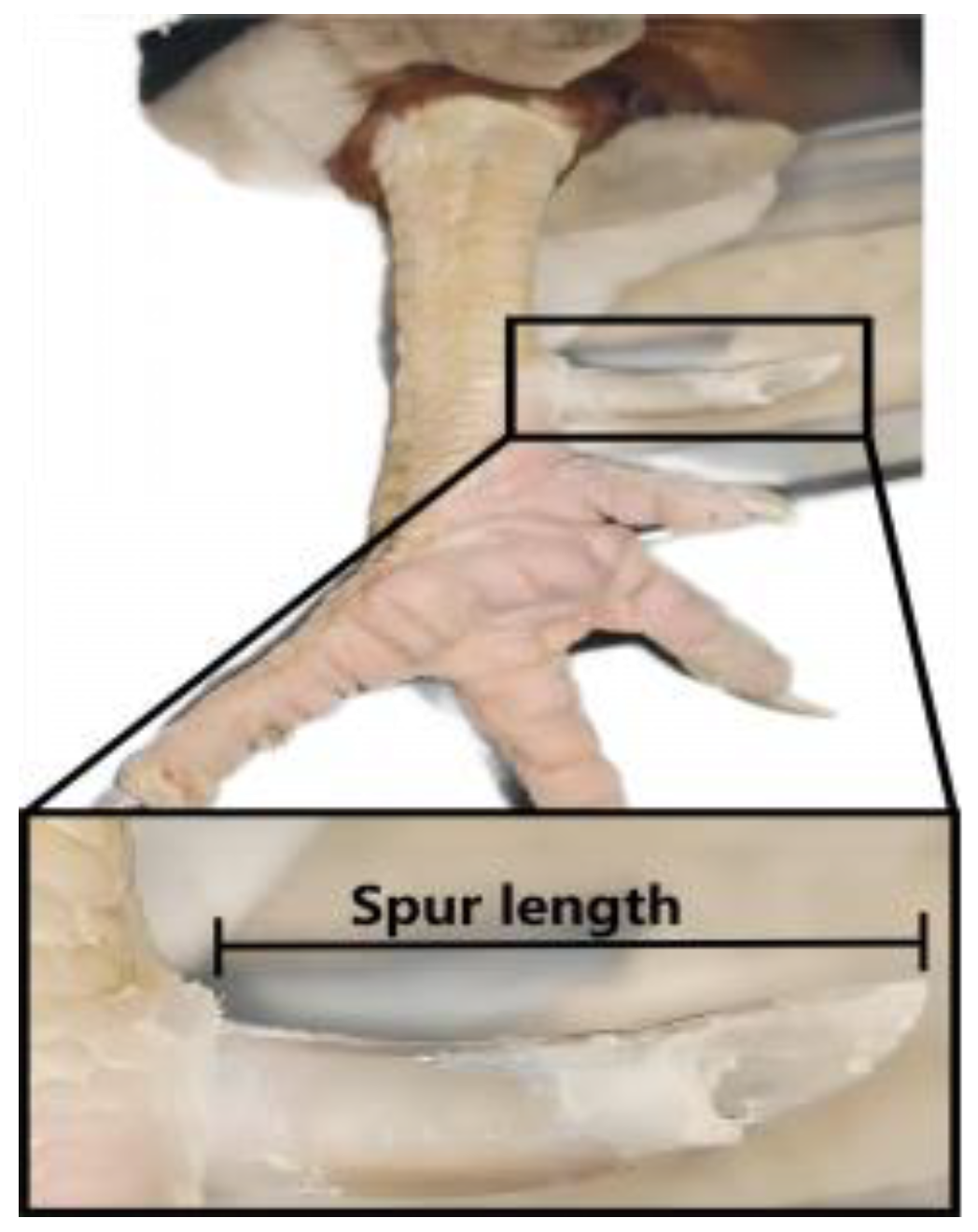
2.2. Animal and Chicken Phenotypic Measurement
2.3. Genetic Parameter Estimation
2.4. Genomic Analysis
3. Results
3.1. Heritability and Correlation Analysis
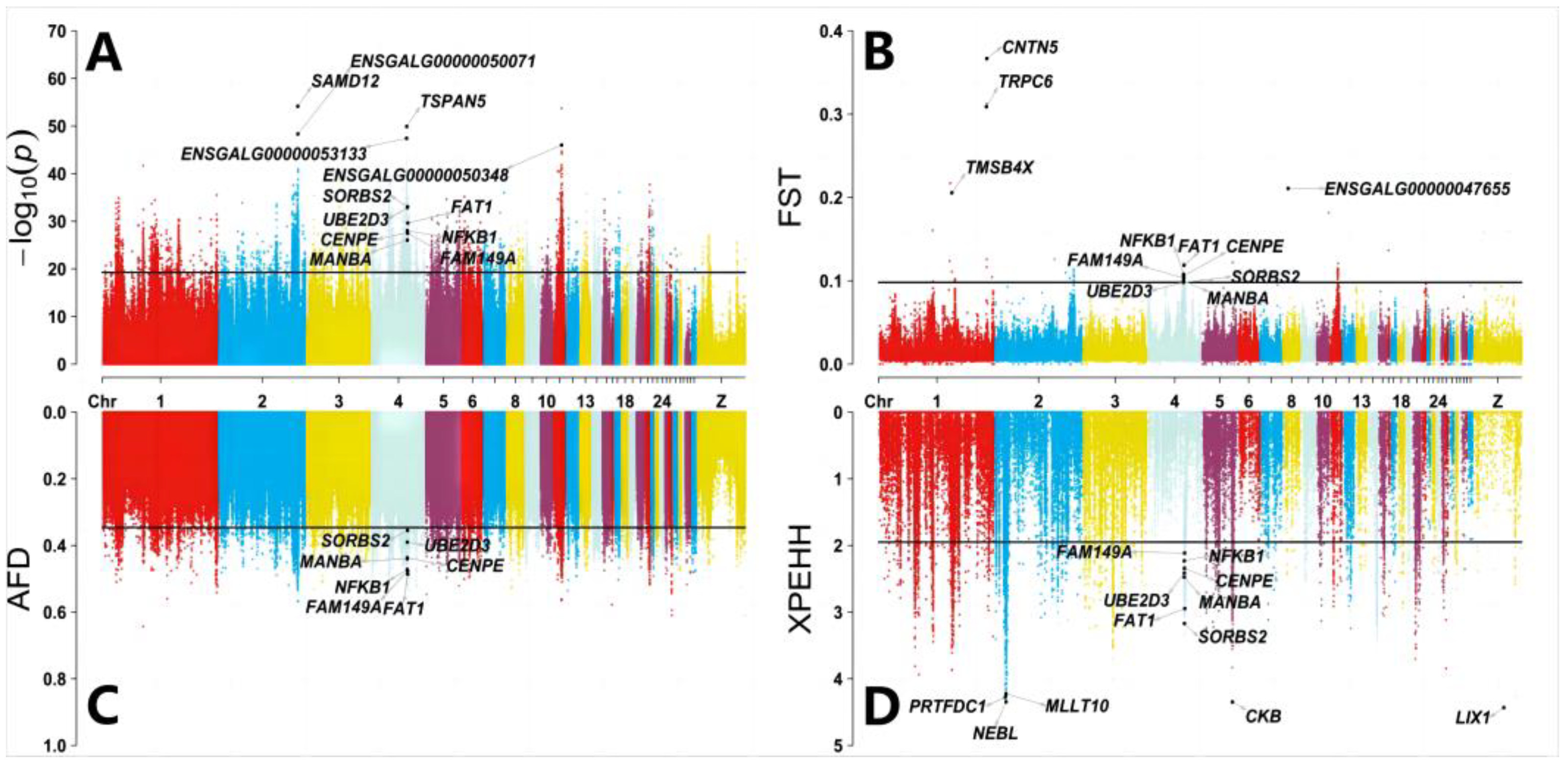
3.2. Pool-GWAS and Genome-Wide Selection Signature Analysis
4. Discussion
4.1. Genetic Parameter Estimates
4.2. Gene Function Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, N. A gene for color differences between sexes. Science 2020, 368, 1185–1186. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Lee, J.; Lee, B.; Lee, K. Research Note: Muscle hypertrophy is associated with reversed sexual dimorphism in body size of quail. Poult. Sci. 2024, 103, 103263. [Google Scholar] [CrossRef] [PubMed]
- Roulin, A.; Jensen, H. Sex-linked inheritance, genetic correlations and sexual dimorphism in three melanin-based colour traits in the barn owl. J. Evol. Biol. 2015, 28, 655–666. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Liu, Q.; Wang, P.; Liu, B.; Sun, W.; Pan, B. Sex dimorphism in the deutonymphs of Dermanyssus gallinae (De Geer, 1778) based on geometric morphometrics. Exp. Parasitol. 2023, 249, 108530. [Google Scholar] [CrossRef] [PubMed]
- Shogren, E.H.; Anciães, M.; Barske, J.; Cestari, C.; DuVal, E.H.; Gaiotti, M.G.; Johnson, E.I.; Kimball, R.T.; Marini, M.A.; Ryder, T.B.; et al. Dancing drives evolution of sexual size dimorphism in manakins. Proc. Biol. Sci. 2022, 289, 20212540. [Google Scholar] [CrossRef] [PubMed]
- Langmore, N.E. Female birdsong. Curr. Biol. 2020, 30, R789–R790. [Google Scholar] [CrossRef]
- Halgrain, M.; Bernardet, N.; Hennequet-Antier, C.; Réhault-Godbert, S. Sex-specific transcriptome of the chicken chorioallantoic membrane. Genomics 2023, 116, 110754. [Google Scholar] [CrossRef] [PubMed]
- Washburn, K.W.; Smyth, J.R., Jr. Inheritance of auxiliary spur in the domestic fowl. Poult. Sci. 1971, 50, 385–388. [Google Scholar] [CrossRef]
- Christmas, R.B.; Harms, R.H. Observation of Spurs in Four Strains of White Leghorn Hens as Affected by Season of Maturity and Dietary Nutrient Level. Poult. Sci. 1982, 61, 1037–1040. [Google Scholar] [CrossRef]
- Fairfull, R.W.; Gowe, R.S. Genotypic and phenotypic parameters of spur incidence and length in White Leghorn hens. Poult. Sci. 1986, 65, 1995–2001. [Google Scholar] [CrossRef]
- Campo, J.L.; Gil, M.G.; Dávila, S.G.; Muñoz, I. Effect of lighting stress on fluctuating asymmetry, heterophil-to-lymphocyte ratio, and tonic immobility duration in eleven breeds of chickens. Poult. Sci. 2007, 86, 37–45. [Google Scholar] [CrossRef]
- Morris, K.R.; Hirst, C.E.; Major, A.T.; Ezaz, T.; Ford, M.; Bibby, S.; Doran, T.J.; Smith, C.A. Gonadal and Endocrine Analysis of a Gynandromorphic Chicken. Endocrinology 2018, 159, 3492–3502. [Google Scholar] [CrossRef]
- Valdez, M.B., Jr.; Mizutani, M.; Kinoshita, K.; Fujiwara, A.; Yazawa, H.; Shimada, K.; Namikawa, T.; Yamagata, T. Differential development of sex-related characters of chickens from the GSP and PNP/DO inbred lines after left ovariectomy. J. Reprod. Dev. 2010, 56, 154–161. [Google Scholar] [CrossRef]
- Blomqvist, A.; Berg, C.; Holm, L.; Brandt, I.; Ridderstrale, Y.; Brunstrom, B. Defective Reproductive Organ Morphology and Function in Domestic Rooster Embryonically Exposed to o,p′-DDT or Ethynylestradiol1. Biol. Reprod. 2006, 74, 481–486. [Google Scholar] [CrossRef]
- Ohlsson, T.; Smith, H.G.; Råberg, L.; Hasselquist, D. Pheasant sexual ornaments reflect nutritional conditions during early growth. Proc. Biol. Sci. 2002, 269, 21–27. [Google Scholar] [CrossRef]
- von Schantz, T.; Wittzell, H.; Göransson, G.; Grahn, M.; Persson, K. MHC genotype and male ornamentation: Genetic evidence for the Hamilton-Zuk model. Proc. Biol. Sci. 1996, 263, 265–271. [Google Scholar] [CrossRef]
- Göransson, G.; von Schantz, T.; Fröberg, I.; Helgee, A.; Wittzell, H. Male characteristics, viability and harem size in the pheasant, Phasianus colchicus. Anim. Behav. 1990, 40, 89–104. [Google Scholar] [CrossRef]
- Grahn, M.; Von Schantz, T. Fashion and Age in Pheasants: Age Differences in Mate Choice. Proc. Biol. Sci. 1994, 255, 237–241. [Google Scholar]
- Porter, C.K.; Romero, F.G.; Adams, D.C.; Bowie, R.C.K.; Riddell, E.A. Adaptive and non-adaptive convergent evolution in feather reflectance of California Channel Islands songbirds. Proc. Biol. Sci. 2023, 290, 20231914. [Google Scholar] [CrossRef]
- Madsen, P.; Jensen, J. A user’s guide to DMU. A Package for Analysing Multivariate Mixed Models Version; Danish Institute of Agricultural Sciences: Tjele, Denmark, 2013; Volume 6, pp. 1–33. [Google Scholar]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. GigaScience 2021, 10, 4. [Google Scholar] [CrossRef]
- Kofler, R.; Pandey, R.V.; Schlötterer, C. PoPoolation2: Identifying differentiation between populations using sequencing of pooled DNA samples (Pool-Seq). Bioinformatics 2011, 27, 3435–3436. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Bastide, H.; Betancourt, A.; Nolte, V.; Tobler, R.; Stöbe, P.; Futschik, A.; Schlötterer, C. A genome-wide, fine-scale map of natural pigmentation variation in Drosophila melanogaster. PLoS Genet. 2013, 9, e1003534. [Google Scholar] [CrossRef]
- Szpiech, Z.A.; Hernandez, R.D. selscan: An efficient multithreaded program to perform EHH-based scans for positive selection. Mol. Biol. Evol. 2014, 31, 2824–2827. [Google Scholar] [CrossRef]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef]
- Régnier, V.; Vagnarelli, P.; Fukagawa, T.; Zerjal, T.; Burns, E.; Trouche, D.; Earnshaw, W.; Brown, W. CENP-A is required for accurate chromosome segregation and sustained kinetochore association of BubR1. Mol. Cell. Biol. 2005, 25, 3967–3981. [Google Scholar] [CrossRef]
- Saffery, R.; Earle, E.; Irvine, D.V.; Kalitsis, P.; Choo, K.H. Conservation of centromere protein in vertebrates. Chromosome Res. 1999, 7, 261–265. [Google Scholar] [CrossRef]
- Suzuki, A.; Hori, T.; Nishino, T.; Usukura, J.; Miyagi, A.; Morikawa, K.; Fukagawa, T. Spindle microtubules generate tension-dependent changes in the distribution of inner kinetochore proteins. J. Cell Biol. 2011, 193, 125–140. [Google Scholar] [CrossRef]
- Samejima, I.; Spanos, C.; Alves Fde, L.; Hori, T.; Perpelescu, M.; Zou, J.; Rappsilber, J.; Fukagawa, T.; Earnshaw, W.C. Whole-proteome genetic analysis of dependencies in assembly of a vertebrate kinetochore. J. Cell Biol. 2015, 211, 1141–1156. [Google Scholar] [CrossRef]
- Álvarez-Hernán, G.; Garrido-Jiménez, S.; Román, Á.C.; Carvajal-González, J.M.; Francisco-Morcillo, J. Distribution of planar cell polarity proteins in the developing avian retina. Exp. Eye Res. 2021, 209, 108681. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhai, B.; Yuan, P.; Fan, S.; Jin, W.; Li, W.; Sun, G.; Tian, Y.; Liu, X.; Kang, X.; et al. MiR-29b-1-5p regulates the proliferation and differentiation of chicken primary myoblasts and analysis of its effective targets. Poult. Sci. 2022, 101, 101557. [Google Scholar] [CrossRef] [PubMed]
- Oikkonen, J.; Onkamo, P.; Järvelä, I.; Kanduri, C. Convergent evidence for the molecular basis of musical traits. Sci. Rep. 2016, 6, 39707. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Hu, G.; Cao, H.; Guo, X. Quercetin Ameliorates Lipopolysaccharide-Induced Duodenal Inflammation through Modulating Autophagy, Programmed Cell Death and Intestinal Mucosal Barrier Function in Chicken Embryos. Animals 2022, 12, 3524. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, Y.H.; Teng, P.; Pazdro, R.; Kim, W.K. Long Bone Mineral Loss, Bone Microstructural Changes and Oxidative Stress After Eimeria Challenge in Broilers. Front. Physiol. 2022, 13, 945740. [Google Scholar] [CrossRef] [PubMed]
- Swaggerty, C.L.; Byrd, J.A., 2nd; Arsenault, R.J.; Perry, F.; Johnson, C.N.; Genovese, K.J.; He, H.; Kogut, M.H.; Piva, A.; Grilli, E. A blend of microencapsulated organic acids and botanicals reduces necrotic enteritis via specific signaling pathways in broilers. Poult. Sci. 2022, 101, 101753. [Google Scholar] [CrossRef]
- Palamidi, I.; Mountzouris, K.C. Diet supplementation with an organic acids-based formulation affects gut microbiota and expression of gut barrier genes in broilers. Anim. Nutr. 2018, 4, 367–377. [Google Scholar] [CrossRef]
- Ge, C.; Yu, M.; Petitte, J.N.; Zhang, C. Epidermal growth factor-induced proliferation of chicken primordial germ cells: Involvement of calcium/protein kinase C and NFKB1. Biol. Reprod. 2009, 80, 528–536. [Google Scholar] [CrossRef]
- Mariani, P.; Barrow, P.A.; Cheng, H.H.; Groenen, M.M.; Negrini, R.; Bumstead, N. Localization to chicken chromosome 5 of a novel locus determining salmonellosis resistance. Immunogenetics 2001, 53, 786–791. [Google Scholar] [CrossRef]
- Polansky, O.; Seidlerova, Z.; Faldynova, M.; Sisak, F.; Rychlik, I. Protein expression in the liver and blood serum in chickens in response to Salmonella Enteritidis infection. Vet. Immunol. Immunopathol. 2018, 205, 10–16. [Google Scholar] [CrossRef]
- Bottje, W.G.; Lassiter, K.; Dridi, S.; Hudson, N.; Kong, B.W. Enhanced expression of proteins involved in energy production and transfer in breast muscle of pedigree male broilers exhibiting high feed efficiency. Poult. Sci. 2017, 96, 2454–2458. [Google Scholar] [CrossRef] [PubMed]
- Swindell, E.C.; Moeller, C.; Thaller, C.; Eichele, G. Cloning and expression analysis of chicken Lix1, a founding member of a novel gene family. Mech. Dev. 2001, 109, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Scheibinger, M.; Ellwanger, D.C.; Krey, J.F.; Choi, D.; Kelly, R.T.; Heller, S.; Barr-Gillespie, P.G. Single-cell proteomics reveals changes in expression during hair-cell development. eLife 2019, 8, e50777. [Google Scholar] [CrossRef] [PubMed]
- Qiao, N.; Pan, J.; Kang, Z.; Liu, G.; Tang, Z.; Li, Y. Effect of a background Ca(2+) entry pathway mediated by TRPC1 on myocardial damage of broilers with induced ascites syndrome. Avian Pathol. 2019, 48, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Guillemette, J.; Caron, A.Z.; Regimbald-Dumas, Y.; Arguin, G.; Mignery, G.A.; Boulay, G.; Guillemette, G. Expression of a truncated form of inositol 1,4,5-trisphosphate receptor type III in the cytosol of DT40 triple inositol 1,4,5-trisphosphate receptor-knockout cells. Cell Calcium 2005, 37, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Quigley, G.D.; Juhn, M. A Comparison of Spur Growth in the Cock, Slip, and Capon*. Poult. Sci. 1951, 30, 900–901. [Google Scholar] [CrossRef]
- Li, X.; Wang, Z.; Yang, B. Identification of the hub genes linked to zearalenone-induced hepatotoxicity in broiler chickens. Environ. Res. 2024, 246, 118094. [Google Scholar] [CrossRef]
- Giles, T.; van Limbergen, T.; Sakkas, P.; Belkhiri, A.; Maes, D.; Kyriazakis, I.; Mendez, J.; Barrow, P.; Foster, N. Differential gene response to coccidiosis in modern fast growing and slow growing broiler genotypes. Vet. Parasitol. 2019, 268, 1–8. [Google Scholar] [CrossRef]
- Reppert, S.M.; Weaver, D.R.; Cassone, V.M.; Godson, C.; Kolakowski, L.F., Jr. Melatonin receptors are for the birds: Molecular analysis of two receptor subtypes differentially expressed in chick brain. Neuron 1995, 15, 1003–1015. [Google Scholar] [CrossRef]
- Nie, C.; Zhang, Z.; Zheng, J.; Sun, H.; Ning, Z.; Xu, G.; Yang, N.; Qu, L. Genome-wide association study revealed genomic regions related to white/red earlobe color trait in the Rhode Island Red chickens. BMC Genet. 2016, 17, 115. [Google Scholar] [CrossRef]
- Liu, H.; Li, X.; Zhang, K.; Lv, X.; Zhang, Q.; Chen, P.; Wang, Y.; Zhao, J. Integrated multi-omics reveals the beneficial role of chlorogenic acid in improving the growth performance and immune function of immunologically stressed broilers. Anim. Nutr. 2023, 14, 383–402. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Trait | Right Spur Length | Left Spur Length | BW18 1 | SL18 2 | AFE 3 | BWA 4 | FEW 5 |
|---|---|---|---|---|---|---|---|
| Right spur length | 0.72 | ||||||
| Left spur length | 0.844 ** | 0.67 | |||||
| BW18 | 0.224 ** | 0.173 ** | - | ||||
| SL18 | 0.178 ** | 0.190 ** | 0.495 ** | - | |||
| AFE | 0.101 ** | 0.100 ** | −0.037 | 0.021 | - | ||
| BWA | 0.069 * | 0.069 * | 0.191 ** | 0.107 ** | 0.241 ** | - | |
| FEW | −0.008 | 0.013 | −0.023 | 0.027 | 0.587 ** | 0.268 ** | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, A.; Zhao, X.; Zhao, X.; Wang, G.; Zhang, X.; Ren, X.; Zhang, Y.; Cheng, X.; Yu, X.; Wang, H.; et al. Genetic Foundation of Male Spur Length and Its Correlation with Female Egg Production in Chickens. Animals 2024, 14, 1780. https://doi.org/10.3390/ani14121780
Chen A, Zhao X, Zhao X, Wang G, Zhang X, Ren X, Zhang Y, Cheng X, Yu X, Wang H, et al. Genetic Foundation of Male Spur Length and Its Correlation with Female Egg Production in Chickens. Animals. 2024; 14(12):1780. https://doi.org/10.3390/ani14121780
Chicago/Turabian StyleChen, Anqi, Xiaoyu Zhao, Xiurong Zhao, Gang Wang, Xinye Zhang, Xufang Ren, Yalan Zhang, Xue Cheng, Xiaofan Yu, Huie Wang, and et al. 2024. "Genetic Foundation of Male Spur Length and Its Correlation with Female Egg Production in Chickens" Animals 14, no. 12: 1780. https://doi.org/10.3390/ani14121780
APA StyleChen, A., Zhao, X., Zhao, X., Wang, G., Zhang, X., Ren, X., Zhang, Y., Cheng, X., Yu, X., Wang, H., Guo, M., Jiang, X., Mei, X., Wei, G., Wang, X., Jiang, R., Guo, X., Ning, Z., & Qu, L. (2024). Genetic Foundation of Male Spur Length and Its Correlation with Female Egg Production in Chickens. Animals, 14(12), 1780. https://doi.org/10.3390/ani14121780