
Metabolism and Nutrition of L-Glutamate and L-Glutamine in Ruminants
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction

{kind=link}
| AAs | AAs in Feeds | AAs in Ruminal | Free AAs in | AAs in Skeletal | AAs in the Whole | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| (g/100 g AAs) | Bacterial Proteins | Plasma (µmol/L) | Muscle Proteins | Body | ||||||
| (g/100 g AAs) b | (g/100 g AAs) b | (mg/g AA) b | ||||||||
| Sheep | Cattle | Sheep | Cattle | Sheep | Cattle | Sheep | Cattle | Sheep | Cattle d | |
| Ala | 6.53 | 8.02 | 6.74 | 6.72 | 182 | 181 | 5.52 | 5.55 | 66.5 | 71.2 |
| Arg | 5.91 | 5.18 | 5.03 | 5.01 | 190 | 121 | 6.58 | 6.57 | 68.0 | 70.2 |
| Asn | 5.13 | 4.71 | 5.34 | 5.36 | 33 | 31 | 4.16 | 4.18 | 35.8 | 36.7 |
| Asp | 5.83 | 6.58 | 6.74 | 6.75 | 11 | 5.4 | 5.15 | 5.16 | 43.7 | 44.8 |
| Cys c | 1.87 | 1.61 | 1.48 | 1.49 | 114 | 132 | 1.38 | 1.35 | 14.6 | 13.4 |
| Gln | 9.02 | 5.95 | 5.11 | 5.13 | 372 | 286 | 5.66 | 5.64 | 50.9 | 49.1 |
| Glu | 7.85 | 10.8 | 8.02 | 7.99 | 61 | 52 | 9.35 | 9.32 | 83.2 | 80.2 |
| Gly | 4.90 | 4.93 | 5.06 | 5.07 | 511 | 347 | 4.17 | 4.18 | 113 | 113.3 |
| His | 2.18 | 2.28 | 2.05 | 2.07 | 62 | 67 | 3.94 | 3.95 | 21.2 | 25.3 |
| Ile | 4.20 | 4.47 | 5.53 | 5.51 | 62 | 100 | 5.13 | 5.15 | 36.0 | 28.1 |
| Leu | 8.32 | 8.67 | 7.67 | 7.66 | 107 | 148 | 8.34 | 8.33 | 69.4 | 69.3 |
| Lys | 4.98 | 4.66 | 7.70 | 7.70 | 94 | 104 | 9.02 | 9.03 | 61.0 | 64.6 |
| Met | 1.63 | 1.79 | 2.42 | 2.40 | 24 | 27 | 3.18 | 3.17 | 19.0 | 18.8 |
| Phe | 4.90 | 5.30 | 5.13 | 5.16 | 36 | 51 | 4.19 | 4.18 | 34.6 | 36.5 |
| Pro | 7.93 | 4.9 | 3.67 | 3.66 | 156 | 184 | 4.06 | 4.08 | 85.5 | 81.5 |
| Ser | 5.05 | 4.56 | 4.65 | 4.62 | 75 | 67 | 4.38 | 4.41 | 45.2 | 44.0 |
| Thr | 3.81 | 4.78 | 5.52 | 5.57 | 60 | 62 | 4.61 | 4.59 | 36.8 | 40.3 |
| Trp | 1.24 | 1.53 | 1.39 | 1.38 | 39 | 49 | 1.26 | 1.25 | 11.4 | 11.4 |
| Tyr | 3.73 | 3.42 | 4.65 | 4.63 | 61 | 70 | 3.76 | 3.75 | 27.0 | 25.3 |
| Val | 4.98 | 5.91 | 6.08 | 6.11 | 128 | 224 | 5.96 | 5.94 | 42.6 | 39.3 |
| Hyp | ND | ND | ND | ND | 41 | 45 | 0.20 | 0.21 | 34.8 | 36.7 |
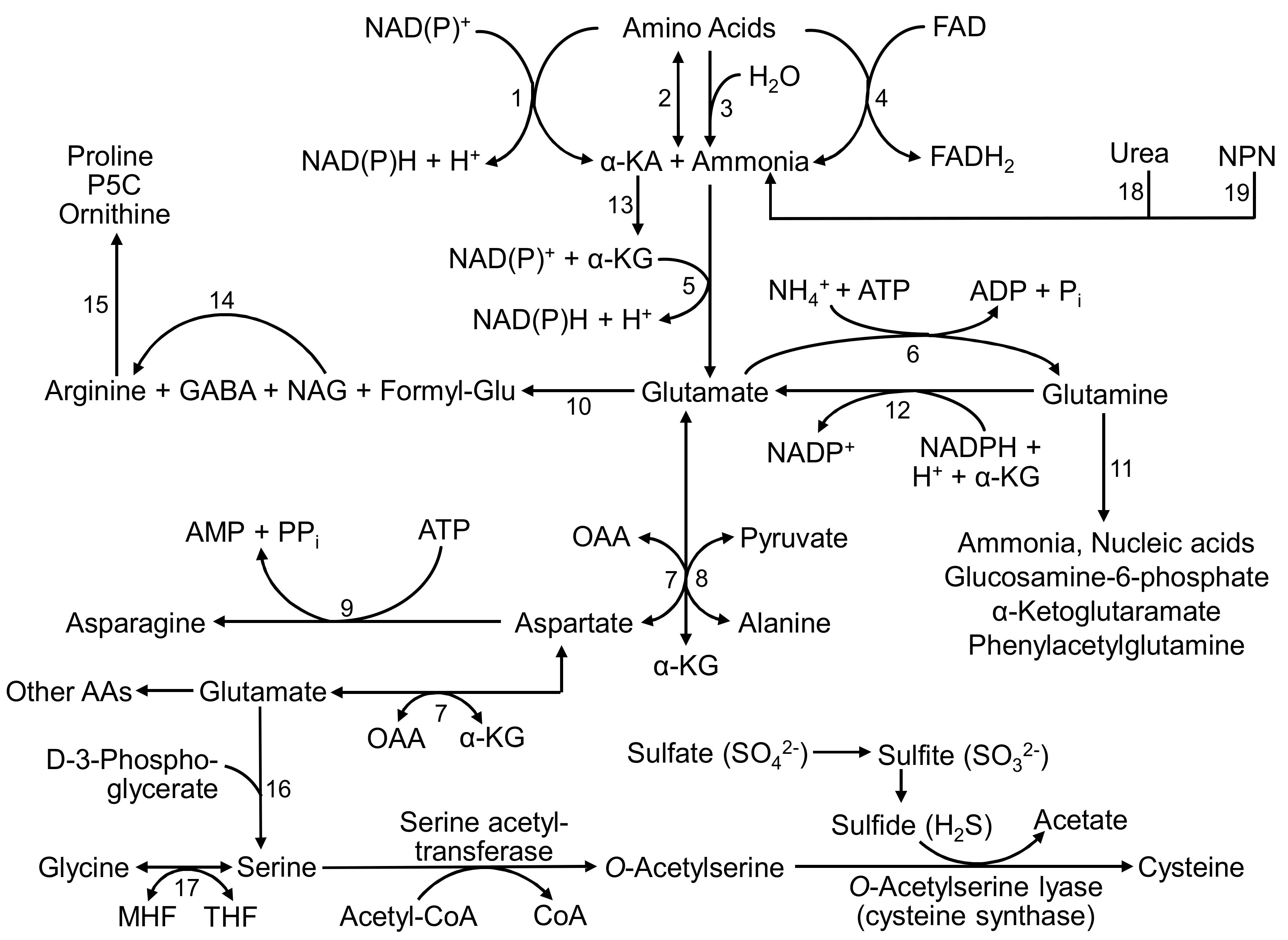
2. Glu and Gln Metabolism in Ruminants
2.1. Glu and Gln Synthesis in Ruminants
2.1.1. Glu and Gln Synthesis in the Rumen
2.1.2. Glu and Gln Synthesis in Extra-Ruminal Tissues
2.2. Glu and Gln Catabolism in Ruminants
2.2.1. Glu and Gln Catabolism in the Rumen
2.2.2. Glu Catabolism in Extra-Ruminal Tissues
2.2.3. Gln Catabolism in Extra-Ruminal Tissues
2.3. Developmental Changes in Glu and Gln Metabolism in Postnatal Ruminants
3. Glu and Gln Nutrition in Ruminants
3.1. Glu Nutrition in Ruminants
3.1.1. Glu Nutrition and Feed Intake in Calves
3.1.2. Glu Nutrition and Feed Intake in Lambs
3.1.3. Glu Nutrition and Feed Intake in Adult Sheep
3.1.4. Glu Nutrition in Rams
3.1.5. Glu Nutrition in Female Goats
3.1.6. Glu Nutrition in Beef Cattle
3.1.7. Glu Nutrition in Dairy Cows
3.1.8. Safety of Glu Supplementation in Ruminants
3.2. Gln Nutrition in Ruminants
3.2.1. Gln Nutrition in Healthy Calves
3.2.2. Gln Nutrition in Calves with Diarrhea
3.2.3. Gln Nutrition in Beef Cattle
3.2.4. Gln Nutrition in Dairy Cows
3.2.5. Gln Nutrition and Alleviation of Infections by Internal Parasites in Sheep
3.2.6. Gln Nutrition and Alleviation of an Acid-Base Imbalance in Gestating Sheep
3.2.7. Gln Supplementation to Improve Ruminal Function in Lambs
3.2.8. Gln Supplementation to Alleviate Heat Stress in Goats and Lambs
3.2.9. Gln Nutrition in Female Goats
3.2.10. Gln Nutrition in Rams
3.2.11. Safety of Gln Supplementation in Ruminants
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, G. Amino Acids: Biochemistry and Nutrition; CRC Press: Boca Raton, FL, USA, 2022. [Google Scholar]
- Hugentobler, S.A.; Diskin, M.G.; Leese, H.J.; Humpherson, P.G.; Watson, T.; Sreenan, J.M.; Morris, D.G. Amino acids in oviduct and uterine fluid and blood plasma during the estrous cycle in the bovine. Mol. Reprod. Dev. 2007, 74, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.L.; Matsumura, M.; Obitsu, T.; Sugino, T. Phytol supplementation alters plasma concentrations of formate, amino acids, and lipid metabolites in sheep. Animal 2021, 15, 100174. [Google Scholar] [CrossRef] [PubMed]
- Gilbreath, K.R.; Bazer, F.W.; Satterfield, M.C.; Wu, G. Amino acid nutrition and reproductive performance in ruminants. Adv. Exp. Med. Biol. 2021, 1285, 43–61. [Google Scholar] [PubMed]
- Wu, G.; Bazer, F.W.; Dai, Z.L.; Li, D.F.; Wang, J.J.; Wu, Z.L. Amino acid nutrition in animals: Protein synthesis and beyond. Annu. Rev. Anim. Biosci. 2014, 2, 387–417. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.P.; Hewitt, D. The amino acid requirements of the preruminant calf. Br. J. Nutr. 1979, 41, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Okine, E.K.; Glimm, D.R.; Thompson, J.R.; Kennelly, J.J. Influence of stage of lactation on glucose and glutamine metabolism in isolated enterocytes from dairy cattle. Metabolism 1995, 44, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.Q.; Wu, G. L-Glutamate nutrition and metabolism in swine. Amino Acids 2018, 50, 1497–1510. [Google Scholar] [CrossRef] [PubMed]
- Runyan, C.A.; Downey-Slinker, E.D.; Ridpath, J.F.; Hairgrove, T.B.; Sawyer, J.E.; Herring, A.D. Feed intake and weight changes in Bos indicus-Bos taurus crossbred steers following bovine viral diarrhea virus type 1b challenge under production conditions. Pathogens 2017, 6, 66. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, C.M. The Welfare of Sheep; Springer: New York, NY, USA, 2008. [Google Scholar]
- Koeln, L.L.; Schlagheck, T.G.; Webb, K.E., Jr. Amino acid flux across the gastrointestinal tract and liver of calves. J. Dairy Sci. 1993, 76, 2275–2285. [Google Scholar] [CrossRef]
- Pelaez, R.; Phillips, D.D.; Walker, D.M. Amino acid supplementation of isolated soybean protein in milk replacers for preruminant lambs. Adv. Exp. Med. Biol. 1978, 105, 443–452. [Google Scholar] [PubMed]
- Kepro. Amino Acid Oral. 2017. Available online: https://www.kepro.nl/products/amino-acid-oral-2 (accessed on 5 November 2017).
- Gilbreath, K.R.; Nawaratna, G.I.; Wickersham, T.A.; Satterfield, M.C.; Bazer, F.W.; Wu, G. Ruminal microbes of adult steers do not degrade extracellular L-citrulline and have a limited ability to metabolize extra-cellular L-glutamate. J. Anim. Sci. 2019, 97, 3611–3616. [Google Scholar] [CrossRef] [PubMed]
- Kung, L.M., Jr.; Rode, L.M. Amino acid metabolism in ruminants. Anim. Feed Sci. Technol. 1996, 59, 167–172. [Google Scholar] [CrossRef]
- Lewis, T.R.; Emery, R.S. Metabolism of amino acids in the bovine rumen. J. Dairy Sci. 1962, 45, 1487–1492. [Google Scholar] [CrossRef]
- Bergen, W.G. Amino acids in beef cattle nutrition and production. Adv. Exp. Med. Biol. 2021, 1285, 29–42. [Google Scholar] [PubMed]
- Cao, Y.; Yao, J.; Sun, X.; Liu, S.; Martin, G.B. Amino acids in the nutrition and production of sheep and goats. Adv. Exp. Med. Biol. 2021, 1285, 63–79. [Google Scholar] [PubMed]
- Gilbreath, K.R.; Nawaratna, G.I.; Wickersham, T.A.; Satterfield, M.C.; Bazer, F.W.; Wu, G. Metabolic studies reveal that ruminal microbes of adult steers do not degrade rumen-protected or unprotected L-citrulline. J. Anim. Sci. 2020, 98, skz370. [Google Scholar] [CrossRef] [PubMed]
- Gilbreath, K.R.; Bazer, F.W.; Satterfield, M.C.; Cleere, J.J.; Wu, G. Ruminal microbes of adult sheep do not degrade extracellular L-citrulline. J. Anim. Sci. 2020, 98, skaa164. [Google Scholar] [CrossRef] [PubMed]
- Tagari, H.; Bergman, E.N. Intestinal disappearance and portal blood appearance of amino acids in sheep. J. Nutr. 1978, 108, 790–803. [Google Scholar] [CrossRef]
- El-Kadi, S.W.; Baldwin, R.L., 6th; McLeod, K.R.; Sunny, N.E.; Bequette, B.J. Glutamate is the major anaplerotic substrate in the tricarboxylic acid cycle of isolated rumen epithelial and duodenal mucosal cells from beef cattle. J. Nutr. 2009, 139, 869–875. [Google Scholar] [CrossRef]
- Lapierre, H.; Bernier, J.F.; Dubreuil, P.; Reynolds, C.K.; Farmer, C.; Ouellet, D.R.; Lobley, G.E. The effect of feed intake level on splanchnic metabolism in growing beef steers. J. Anim. Sci. 2000, 78, 1084–1099. [Google Scholar] [CrossRef]
- Wu, G.; Li, P. The “ideal protein” concept is not ideal in animal nutrition. Exp. Biol. Med. 2022, 247, 1191–1201. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Fiorotto, M.L.; Reeds, P.J. Amino acid compositions of body and milk protein change during the suckling period in rats. J. Nutr. 1993, 123, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wu, G. Composition of amino acids and related nitrogenous nutrients in feedstuffs for animal diets. Amino Acids 2020, 52, 523–542. [Google Scholar] [CrossRef] [PubMed]
- Alumot, E.; Bruckental, I.; Tadmor, A.; Kennit, C.; Holstein, P. Effect of proline on arginine uptake and nitrogen metabolism of lactating goats. J. Dairy. Sci. 1983, 66, 1243–1247. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Nguyen, H.V.; Garciaa-Bravo, R.; Fiorotto, M.L.; Jackson, E.M.; Lewis, D.S.; Lee, D.R.; Reeds, P.J. Amino acid composition of human milk is not unique. J. Nutr. 1994, 124, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Mepham, T.B. Amino acid utilization by lactating mammary gland. J. Dairy Sci. 1982, 65, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Principles of Animal Nutrition; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Baetz, A.L.; Hubbert, W.T.; Graham, C.K. Developmental changes of free amino acids in bovine fetal fluids with gestational age and the interrelationships between the amino acid concentrations in the fluid compartments. J. Reprod. Fertil. 1975, 44, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.; Spencer, T.E.; Bazer, F.W.; Wu, G. Developmental changes of amino acids in ovine fetal fluids. Biol. Reprod. 2003, 68, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.; Wu, G.; Bazer, F.W.; Spencer, T.E. Developmental changes in polyamine levels and synthesis in the ovine conceptus. Biol. Reprod. 2003, 69, 1626–1634. [Google Scholar] [CrossRef] [PubMed]
- Satterfield, M.C.; Gao, H.J.; Li, X.L.; Wu, G.; Johnson, G.A.; Spencer, T.E.; Bazer, F.W. Select nutrients and their associated transporters are increased in the ovine uterus following early progesterone administration. Biol. Reprod. 2010, 82, 224–231. [Google Scholar] [CrossRef]
- Satterfield, M.C.; Dunlap, K.A.; Keisler, D.H.; Bazer, F.W.; Wu, G. Arginine nutrition and fetal brown adipose tissue development in diet-induced obese sheep. Amino Acids 2012, 43, 1593–1603. [Google Scholar] [CrossRef] [PubMed]
- Satterfield, M.C.; Dunlap, K.A.; Keisler, D.H.; Bazer, F.W.; Wu, G. Arginine nutrition and fetal brown adipose tissue development in nutrient-restricted sheep. Amino Acids 2013, 45, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.-K.; Nejad, J.G.; Lee, Y.-S.; Kong, H.-S. Effects of L-glutamine supplementation on degradation rate and rumen fermentation characteristics in vitro. Anim Biosci. 2021, 35, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Malheiros, J.M.; Correia, B.S.B.; Ceribeli, C.; Cardoso, D.R.; Colnago, L.A.; Junior, S.B.; Reecy, J.M.; Mourão, G.B.; Coutinho, L.L.; Palhares, J.C.P.; et al. Comparative untargeted metabolome analysis of ruminal fuid and feces of Nelore steers (Bos indicus). Sci. Rep. 2021, 11, 12752. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Jia, Y.; Li, Q.; Burris, W.R.; Bridges, P.J.; Matthews, J.C. Hepatic glutamate transport and glutamine synthesis capacities are decreased in finished vs. growing beef steers, concomitant with increased GTRAP3-18 content. Amino Acids 2018, 50, 513–525. [Google Scholar] [CrossRef]
- Miles, E.D.; McBride, B.W.; Jia, Y.; Liao, S.F.; Boling, J.A.; Bridges, P.J.; Matthews, J.C. Glutamine synthetase and alanine transaminase expression are decreased in livers of aged vs. young beef cows and GS can be upregulated by 17β-estradiol implants. J. Anim. Sci. 2015, 93, 4500–4509. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.M.; Costa, T.C.; Mendes, T.A.O.; Dutra, L.L.; Silva, D.N.L.; Araújo, R.D.; Serão, N.V.L.; Rennó, L.N.; Silva, Y.F.R.S.; Detmann, E.; et al. Can the post-ruminal urea release impact liver metabolism, and nutritional status of beef cows at late gestation? PLoS ONE 2023, 18, e0293216. [Google Scholar] [CrossRef] [PubMed]
- Blake, J.S.; Salter, D.N.; Smith, R.H. Incorporation of nitrogen into rumen bacterial fractions of steers given protein- and urea-containing diets. Ammonia assimilation into intracellular bacterial amino acids. Br. J. Nutr. 1983, 50, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.W.; Bauman, D.E. Regulation of amino acid metabolism in dairy and beef cattle. In Proceedings of the 21st Annual Southwest Nutrition & Management Conference, Tempe, AZ, USA, 23–24 February 2006; pp. 34–44. [Google Scholar]
- Stoll, B.; Burrin, D.G. Measuring splanchnic amino acid metabolism in vivo using stable isotopic tracers. J. Anim. Sci. 2006, 84 (Suppl. S13), E60–E72. [Google Scholar] [CrossRef]
- Li, P.; He, W.L.; Wu, G. Composition of amino acids in foodstuffs for humans and animals. Adv. Exp. Med. Biol. 2021, 1332, 189–210. [Google Scholar] [PubMed]
- Mahgoub, O.; Lodge, G.A. A comparative study on growth, body composition and carcass tissue distribution in Omani sheep and goats. J. Agric. Sci. 1998, 131, 329–339. [Google Scholar] [CrossRef]
- Odongo, N.E.; Greenwood, S.L.; Or-Rashid, M.M.; Radford, D.; Alzahal, O.; Shoveller, A.K.; Lindinger, M.I.; Matthews, J.C.; McBride, B.W. Effects of nutritionally induced metabolic acidosis with or without glutamine infusion on acid-base balance, plasma amino acids, and plasma nonesterified fatty acids in sheep. J. Anim. Sci. 2009, 87, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Greene, L.W. Glutamine and glucose metabolism in bovine blood lymphocytes. Comp. Biochem. Physiol. 1992, 103B, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Wijayasinghe, M.S.; Milligan, L.P.; Thompson, J.R. In vitro degradation of leucine in muscle, adipose tissue, liver, and kidney of fed and starved sheep. Biosci. Rep. 1983, 3, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Eisemann, J.H.; Huntington, G.B.; Catherman, D.R. Patterns of nutrient interchange and oxygen use among portal-drained viscera, liver, and hindquarters of beef steers from 235 to 525 kg body weight. J. Anim. Sci. 1996, 74, 1812–1831. [Google Scholar] [CrossRef] [PubMed]
- Nieto, R.; Obitsu, T.; Fernández-Quintela, A.; Bremner, D.; Milne, E.; Calder, A.G.; Lobley, G.E. Glutamine metabolism in ovine splanchnic tissues: Effects of infusion of ammonium bicarbonate or amino acids into the abomasum. Br. J. Nutr. 2002, 87, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Huntington, G.B.; Reynolds, C.K. Oxygen consumption and metabolite flux of bovine portal-drained viscera and liver. J. Nutr. 1987, 117, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Ballard, F.J.; Filsell, O.H.; Jarrett, I.G. Amino acid uptake and output by the sheep hind limb. Metabolism 1976, 25, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.; Teng, C.; Timmerman, M.; Meschia, G.; Battaglia, F.C. Production and utilization of amino acids by ovine placenta in vivo. Am. J. Physiol. 1998, 274, E13–E22. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.C.; Huang, J.; Rentfrow, G. High-affinity glutamate transporter and glutamine synthetase content in longissimus dorsi and adipose tissues of growing Angus steers differs among suckling, weanling, backgrounding, and finishing production stages. J. Anim. Sci. 2016, 94, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Pell, J.M.; Jeacock, M.K.; Shepherd, D.A.L. Glutamate and glutamine metabolism in the ovine placenta. J. Agric. Sci. 1983, 101, 275–281. [Google Scholar] [CrossRef]
- Bergman, E.N.; Kaufman, C.F.; Wolff, J.E.; Williams, H.H. Renal metabolism of amino acids and ammonia in fed and fasted pregnant sheep. Am. J. Physiol. 1974, 226, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, R.N.; Bergman, E.N. Glutamine metabolism, interorgan transport and glucogenicity in the sheep. Am. J. Physiol. 1978, 234, E197–E203. [Google Scholar] [CrossRef] [PubMed]
- Eisemann, J.H.; Huntington, G.B. Metabolite flux across portal-drained viscera, liver, and hindquarters of hyperinsulinemic, euglycemic beef steers. J. Anim. Sci. 1994, 72, 2919–2929. [Google Scholar] [CrossRef]
- Lei, J.; Feng, D.Y.; Zhang, Y.L.; Zhao, F.Q.; Wu, Z.L.; San Gabriel, A.; Fujishima, Y.; Uneyama, H.; Wu, G. Nutritional and regulatory role of branched-chain amino acids in lactation. Front. Biosci. 2012, 17, 2725–2739. [Google Scholar] [CrossRef] [PubMed]
- Lobley, G.E.; Milano, G.D. Regulation of hepatic nitrogen metabolism in ruminants. Proc. Nutr. Soc. 1997, 56, 547–563. [Google Scholar] [CrossRef]
- Li, P.; Knabe, D.A.; Kim, S.W.; Lynch, C.J.; Hutson, S.M.; Wu, G. Lactating porcine mammary tissue catabolizes branched-chain amino acids for glutamine and aspartate synthesis. J. Nutr. 2009, 139, 502–1509. [Google Scholar] [CrossRef] [PubMed]
- Lassala, A.; Bazer, F.W.; Cudd, T.A.; Datta, S.; Keisler, D.H.; Satterfield, M.C.; Spencer, T.E.; Wu, G. Parenteral administration of L-arginine prevents fetal growth restriction in undernourished ewes. J. Nutr. 2010, 140, 1242–1248. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, R.N.; Hoover, W.H.; Sniffen, C.J. Gluconeogenesis from amino acids in mature wether sheep. J. Nutr. 1973, 103, 1587–1593. [Google Scholar] [CrossRef] [PubMed]
- Tanha, T.; Amanlou, H.; Chamani, M.; Ebrahimnezhad, Y.; Salamatdost, R.; Maheri, N.; Fathi, M. Impact of glutamine on glutathione peroxidase activity (GPX) and total antioxidant status (TAS) during transition period in Holstein dairy cows. J. Cell Anim. Biol. 2011, 5, 206–214. [Google Scholar]
- Loor, J.J.; Lopreiato, V.; Palombo, V.; D’Andrea, M. Physiological impact of amino acids during heat stress in ruminants. Anim. Front. 2023, 13, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Caroprese, M.; Albenzio, M.; Marino, R.; Santillo, A.; Sevi, A. Immune response and milk production of dairy cows fed graded levels of rumen-protected glutamine. Res. Vet. Sci. 2011, 93, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Ocheja, O.B.; Ayo, J.O.; Aluwong, T.; Minka, N.S. Effects of L-glutamine on rectal temperature and some markers of oxidative stress in Red Sokoto goats during the hot-dry season. Trop. Anim. Health Prod. 2017, 49, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Hoskin, S.O.; Lobley, G.E.; Coop, R.L.; Jackson, F. The effect of cysteine and glutamine supplementation on sheep infected with Trichostrongylus colubriformis. Proc. N. Z. Soc. Anim. Prod. 2002, 62, 72–76. [Google Scholar]
- Jafari, A.; Emmanuel, D.G.V.; Christopherson, R.J.; Thompson, J.R.; Murdoch, G.K.; Woodward, J.; Field, C.J.; Ametaj, B.N. Parenteral administration of glutamine modulates acute phase response in postparturient dairy cows. J. Dairy Sci. 2006, 89, 4660–4668. [Google Scholar] [CrossRef]
- McNeil, C.J.; Hoskin, S.O.; Bremner, D.M.; Holtrop, G.; Lobley, G.E. Whole-body and splanchnic amino acid metabolism in sheep during an acute endotoxin challenge. Br. J. Nutr. 2016, 116, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Sawant, O.B.; Wu, G.; Washburn, S.E. Maternal L-glutamine supplementation prevents prenatal alcohol exposure-induced fetal growth restriction in ewes. Amino Acids 2015, 47, 1183–1192. [Google Scholar] [CrossRef]
- Wang, S.; Wang, F.; Kong, F.; Cao, Z.; Wang, W.; Yang, H.; Wang, Y.; Bi, Y.; Li, S. Effect of supplementing different levels of L-glutamine on Holstein calves during weaning. Antioxidants 2022, 11, 542. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Satterfield, M.C.; Gilbreath, K.R.; Posey, E.A.; Sun, Y.X. L-Arginine nutrition and metabolism in ruminants. Adv. Exp. Med. Biol. 2022, 1354, 177–206. [Google Scholar] [PubMed]
- National Research Council (NRC). Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids; National Academy Press: Washington, DC, USA, 2007. [Google Scholar]
- Brake, D.W.; Titgemeyer, E.C.; Anderson, D.E. Duodenal supply of glutamate and casein both improve intestinal starch digestion in cattle but by apparently different mechanisms. J. Anim. Sci. 2014, 92, 4057–4067. [Google Scholar] [CrossRef] [PubMed]
- Howell, J.A.; Matthews, A.D.; Swanson, K.C.; Harmon, D.L.; Matthews, J.C. Molecular identification of high-affinity glutamate transporters in sheep and cattle forestomach, intestine, liver, kidney, and pancreas. J. Anim. Sci. 2001, 79, 1329–1336. [Google Scholar] [CrossRef] [PubMed]
- Huntington, G.B.; Prior, R.L. Net absorption of amino acids by portal-drained viscera and hind half of beef cattle fed a high concentrate diet. J. Anim. Sci. 1985, 60, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.E.; Bergman, E.N. Metabolism and interconversions of five plasma amino acids by tissues of the sheep. Am. J. Physiol. 1972, 223, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Oba, M.; Baldwin, R.L., 4th; Bequette, B.J. Oxidation of glucose, glutamate, and glutamine by isolated ovine enterocytes in vitro is decreased by the presence of other metabolic fuels. J. Anim. Sci. 2004, 82, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Sultana, H.; Kitano, A.; Wadud, S.; Takahashi, T.; Morita, T.; Onodera, R. Synthesis of citrulline from ornithine by the small intestinal mucosa of cattle. Anim. Sci. J. 2003, 74, 283–287. [Google Scholar] [CrossRef]
- Dillon, E.L.; Wu, G. Cortisol enhances citrulline synthesis from proline in enterocytes of suckling piglets. Amino Acids 2021, 53, 1957–1966. [Google Scholar] [CrossRef] [PubMed]
- McCoard, S.A.; Pacheco, D. The signifcance of N-carbamoylglutamate in ruminant production. J. Anim. Sci. Biotechnol. 2023, 14, 48. [Google Scholar] [CrossRef] [PubMed]
- Black, A.L.; Anand, R.S.; Bruss, M.L.; Brown, C.A.; Nakagiri, J.A. Partitioning of amino acids in lactating cows: Oxidation to carbon dioxide. J. Nutr. 1990, 120, 700–710. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Johnson, G.A.; Knabe, D.A.; Burghardt, R.C.; Spencer, T.E.; Li, X.L.; Wang, J.J. Important roles for L-glutamine in swine nutrition and production. J. Anim. Sci. 2011, 89, 2017–2030. [Google Scholar] [CrossRef] [PubMed]
- Burrin, D.G.; Ferrell, C.L.; Eisemann, J.H.; Britton, R.A. Level of nutrition and splanchnic metabolite flux in young lambs. J. Anim. Sci. 1991, 69, 1082–1091. [Google Scholar] [CrossRef]
- Doepel, C.L.; Lobley, G.E.; Bernier, J.F.; Dubreuil, P.; Lapierre, H. Effect of glutamine supplementation on splanchnic metabolism in lactating dairy cows. J. Dairy Sci. 2007, 90, 4325–4333. [Google Scholar] [CrossRef] [PubMed]
- Nappert, G.; Zello, G.A.; Naylor, J.M. Intestinal metabolism of glutamine and potential use of glutamine as a therapeutic agent in diarrhoeic calves. J. Am. Vet. Med. Assoc. 1997, 211, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Egan, A.R.; Black, A.L. Glutamic acid metabolism in the lactating dairy cow. J. Nutr. 1968, 96, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Egan, A.R.; Moller, F.; Black, A.L. Metabolism of glutamic acid, valine and arginine by the lactating goat. J. Nutr. 1970, 100, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, R.N.; Bergman, E.N. Transport of amino acids in whole blood and plasma of sheep. Am. J. Physiol. 1980, 239, E242–E247. [Google Scholar] [PubMed]
- Bergman, E.N.; Heitmann, R.N. Metabolism of amino acids by the gut, liver, kidneys, and peripheral tissues. Fed. Proc. 1978, 37, 1228–1232. [Google Scholar] [PubMed]
- Bröer, S. Amino acid transport across mammalian intestinal and renal epithelia. Physiol. Rev. 2008, 88, 249–286. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Liao, S.F.; Son, K.W.; Greenwood, S.L.; McBride, B.W.; Boling, J.A.; Matthews, J.C. Metabolic acidosis in sheep alters expression of renal and skeletal muscle amino acid enzymes and transporters. J. Anim. Sci. 2010, 88, 707–717. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.; Kristensen, N.B. Effect of abomasal glucose infusion on splanchnic amino acid metabolism in periparturient dairy cows. J. Dairy Sci. 2009, 92, 3306–3318. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.K. Splanchnic metabolism of amino acids in ruminants. In Ruminant Physiology; Sejrsen, K., Hvelplund, T., Nielsen, M.O., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2006; pp. 225–248. [Google Scholar]
- Plaizier, J.C.; Walton, J.P.; McBride, B.W. Effect of post-ruminal infusion of glutamine on plasma amino acids, milk yield and composition in lactating dairy cows. Can. J. Anim. Sci. 2001, 81, 229–235. [Google Scholar] [CrossRef]
- Meijer, G.A.L.; de Visser, H.; van der Meulen, J.; van der Koelen, C.J.; Klop, A. Effect of glutamine or propionate infused into the abomasum on milk yield, milk composition, nitrogen retention and net flux of amino acids across the udder of high yielding dairy cows. In Protein metabolism and nutrition. Proceedings 7th International Symposium, Vale de Santarém, Portugal, 24–27 May 1995; Nunes, A.F., Portugal, A.V., Costa, J.P., Ribeiro, J.R., Eds.; Estacoa Zootechnica National: Santerm, Portugal, 1995; pp. 157–160. [Google Scholar]
- Meijer, G.A.L.; Van der Meulen, J.; Bakker, J.G.M.; Van der Koelen, C.J.; Van Vuuren, A.M. Free amino acids in plasma and muscle of high yielding dairy cows in early lactation. J. Dairy Sci. 1995, 78, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- del Castillo, J.R.; Súlbaran-Carrasco, M.C.; Burguillos, L. Glutamine transport in isolated epithelial intestinal cells. Identification of a Na+-dependent transport mechanism, highly specific for glutamine. Eur. J. Physiol. 2002, 445, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Brockman, R.P.; Bergman, E.N. Effect of glucagon on plasma alanine and glutamine metabolism and hepatic gluconeogenesis in sheep. Am. J. Physiol. 1975, 228, 1628–1633. [Google Scholar] [CrossRef] [PubMed]
- Welbourne, T.C. Interorgan glutamine flow in metabolic acidosis. Am. J. Physiol. 1987, 253, F1069–F1076. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.; Curthoys, N.P. Glutamine metabolism: Role in acid-base balance. Biochem. Mol. Biol. Educ. 2004, 32, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Freetly, H.C.; Ferrell, C.L.; Archibeque, S.L. Net flux of amino acids across splanchnic tissues of ewes during abomasal protein and glucose infusion. In Proceedings of Energy and Protein Metabolism and Nutrition; EAAP Publication No. 124; Wageningen Academic: Wageningen, The Netherlands, 2007; pp. 337–338. [Google Scholar]
- Obitsu, T.; Bremner, D.; Milne, E.; Lobley, G.E. Effect of abomasal glucose infusion on alanine metabolism and urea production in sheep. Br. J. Nutr. 2000, 84, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.; Kristensen, N.B. Effects of glucogenic and ketogenic feeding strategies on splanchnic glucose and amino acid metabolism in postpartum transition Holstein cows. J. Dairy Sci. 2012, 95, 5946–5960. [Google Scholar] [CrossRef] [PubMed]
- Pell, J.M.; Tooley, J.; Jeacock, M.K.; Shepherd, D.A.L. Glutamate and glutamine metabolism in tissues of developing lambs. J. Agric. Sci. 1983, 101, 265–273. [Google Scholar] [CrossRef]
- Masters, C.J.; Horgan, D.J. Glutamate transaminase activity in sheep tissues, and the response to prolonged protein depletion. Aust. J. Biol. Sci. 1962, 15, 690–699. [Google Scholar] [CrossRef]
- Minich, D.M.; Brown, B.I. A review of dietary (phyto)nutrients for glutathione support. Nutrients 2019, 11, 2073. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Burghardt, R.C.; Wu, G.; Johnson, G.A.; Spencer, T.E.; Bazer, F.W. Select nutrients in the ovine uterine lumen: IX. Differential effects of arginine, leucine, glutamine and glucose on interferon tau, orinithine decarboxylase and nitric oxide synthase in the ovine conceptus. Biol. Reprod. 2011, 84, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Sawant, O.B.; Meng, C.; Wu, G.; Washburn, S.E. Prenatal alcohol exposure and maternal glutamine supplementation alter the mTOR signaling pathway in ovine fetal cerebellum and skeletal muscle. Alcohol 2020, 89, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Bucak, M.N.; Turner, P.B.; Sarıozkan, S.; Ulutas, P.A. Comparison of the effects of glutamine and an amino acid solution on post-thawed ram sperm parameters, lipid peroxidation and anti-oxidant activities. Small Rumin. Res. 2009, 81, 13–17. [Google Scholar] [CrossRef]
- Sangeeta, S.; Arangasamy, A.; Kulkarni, S.; Selvaraju, S. Role of amino acids as additives on sperm motility, plasma membrane integrity and lipid peroxidation levels at pre-freeze and post-thawed ram semen. Anim. Reprod. Sci. 2015, 161, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Topraggaleh, T.R.; Shahverdi, A.; Rastegarnia, A.; Ebrahimi, B.; Shafiepour, V.; Sharbatoghli, M.; Esmaeili, V.; Janzamin, E. Effect of cysteine and glutamine added to extender on post-thaw sperm functional parameters of buffalo bull. Andrologia 2014, 46, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.J.; Wu, G.; Spencer, T.E.; Johnson, G.A.; Li, X.L.; Bazer, F.W. Select nutrients in the ovine uterine lumen: I. Amino acids, glucose and ions in uterine lumenal flushings of cyclic and pregnant ewes. Biol. Reprod. 2009, 80, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.A.; Seo, H.; Bazer, F.W.; Wu, G.; Kramer, A.C.; McLendon, B.A.; Cain, J.W. Metabolic pathways utilized by the porcine conceptus, uterus and placenta. Mol. Reprod. Dev. 2023, 90, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Kramer, A.C.; McLendon, B.A.; Cain, J.W.; Burghardt, R.C.; Wu, G.; Bazer, F.W.; Johnson, G.A. Elongating porcine coneptuses utilize glutaminolysis as an anaplerotic pathway to maintain the TCA cycle. Biol. Reprod. 2022, 107, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Széll, A.Z. The effect of glutamine on the development of sheep embryos in vitro. Theriogenology 1995, 44, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Meijer, G.A.L.; van der Meulen, J.; van Vuuren, A.M. Glutamine is a potentially limiting amino acid for milk production in dairy sows: A hypothesis. Metabolism 1993, 42, 356–364. [Google Scholar] [CrossRef]
- Turner, A.E.; Rees, G.; Barrett, D.C.; Reyher, K.K. Does inclusion of glutamine in oral rehydration solutions improve recovery from mild to moderate diarrhoea in preweaned calves? Vet. Rec. 2016, 179, 283–284. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (NRC). Nutrient Requirements of Dairy Cattle, 7th ed.; National Academy Press: Washington, DC, USA, 2001. [Google Scholar]
- Oltjen, R.R.; Robbins, J.D.; Davis, R.E. Studies involving the use of glutamic acid in ruminant nutrition. J. Anim. Sci. 1964, 23, 767–770. [Google Scholar] [CrossRef]
- Waldern, D.E.; Van Dyk, R.D. Effect of monosodium glutamate in starter rations on feed consumption and performance of early weaned calves. J. Dairy Sci. 1971, 54, 262–265. [Google Scholar] [CrossRef]
- Galgan, M.W.; Russell, T.S. Use of Dyna-Ferm and monosodium glutamate in rations for lambs. Wash. Agric. Exp. Stat. Bull. 1968, 695, 1–3. [Google Scholar]
- Colucci, P.E.; Grovum, W.L. Factors affecting the voluntary intake of food by sheep 6. The effect of monosodium glutamate on the palatability of straw diets by sham-fed and normal animals. Br. J. Nutr. 1993, 69, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Meza-Herrera, C.A. Puberty, kisspeptin and glutamate: A ceaseless golden braid. In Advances in Medicine and Biology; Benhardt, L.V., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2012; Volume 52, pp. 97–124. [Google Scholar]
- Torres-Moreno, M.; Meza-Herrera, C.A.; González-Bulnes, A.; López-Medrano, J.I.; Mellado-Bosque, M.; Wurzinger, M.; Trejo-Calzada, R. Effect of exogenous glutamate supply on the onset of puberty in goats: I. Serum levels of insulin. Trop. Subtrop. Agroecosystems 2009, 11, 193–196. [Google Scholar]
- Nakanishi, Y.; Iwasaki, M.; Manda, M. Taste responses of cows to Umami (monosodium glutamate and disodium 5′-inosinate). Anim. Sci. Technol. 1996, 67, 561–566. [Google Scholar]
- Li, C.; Zhang, J.; Li, Y.; Zhao, X.; Liang, H.; Li, K.; Qu, M.; Qiu, Q.; Ouyang, K. Glutamate supplementation improves growth performance, rumen fermentation, and serum metabolites in heat-stressed Hu sheep. Front. Nutr. 2022, 9, 851386. [Google Scholar] [CrossRef] [PubMed]
- Hisadomi, S.; Haruno, A.; Fujieda, T.; Sugino, T.; Oba, M. Rumen-protected glutamate supplementation in dairy cows. J. Dairy Sci. 2022, 105, 3129–3141. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.C.S.; Alves, J.P.M.; Fernandes, C.C.L.; Silva, M.R.L.; Conde, A.J.H.; Teixeira, D.Í.A.; Rondina, D. Use of monosodium-glutamate as a novel dietary supplement strategy for ovarian stimulation in goats. Anim. Reprod. 2023, 20, e20230094. [Google Scholar] [CrossRef] [PubMed]
- Naylor, J.M.; Leibel, T.; Middleton, D.M. Effect of glutamine or glycine containing oral electrolyte solutions on mucosal morphology, clinical and biochemical findings, in calves with viral induced diarrhea. Can. J. Vet. Res. 1997, 61, 43–48. [Google Scholar] [PubMed]
- Brooks, H.W.; White, D.G.; Hall, G.A.; Wagstaff, A.J.; Michell, A.R. Evaluation of a glutamine-containing oral rehydration solution for the treatment of calf diarrhoea using an Escherichia coli model. Vet. J. 1997, 153, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Brooks, H.W.; Hall, G.A.; Wagstaff, A.J.; Michell, A.R. Detrimental effects on villus form during conventional oral rehydration therapy for diarrhoea in calves; alleviation by a nutrient oral rehydration solution containing glutamine. Vet. J. 1998, 155, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Pal, B.; Pachauri, S.P. Effect of oral rehydration in neonatal calves treated for diarrhoea induced with Escherichia coli (0101:K99) infection. Indian J. Vet. Res. 2008, 17, 19–26. [Google Scholar]
- Jin, L.; Dong, G.; Lei, C.; Zhou, J.; Zhang, S. Effects of dietary supplementation of glutamine and mannan oligosaccharides on plasma endotoxin and acute phase protein concentrations and nutrient digestibility in finishing steers. J. Appl. Anim. Res. 2014, 42, 160–165. [Google Scholar] [CrossRef]
- Caroprese, M.; Albenzio, M.; Marino, R.; Santillo, A.; Sevi, A. Dietary glutamine enhances immune responses of dairy cows under high ambient temperature. J. Dairy Sci. 2012, 96, 3002–3011. [Google Scholar] [CrossRef]
- Washburn, S.E.; Sawant, O.B.; Lunde, E.R.; Wu, G.; Cudd, T.A. Acute alcohol exposure, acidemia or glutamine administration impacts amino acid homeostasis in ovine maternal and fetal plasma. Amino Acids 2014, 45, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Nemati, M.; Menatian, S.; Joz Ghasemi, S.; Hooshmandfar, R.; Taheri, M.; Saifi, T. Effect of protected-glutamine supplementation on performance, milk composition and some blood metabolites in fresh Holstein cows. Iran. J. Vet. Res. 2018, 19, 225–228. [Google Scholar] [PubMed]
- Feyz, M.; Teimouri Yansari, A.; Chashnidel, Y.; Dirandeh, E. Effect of protein levels and rumen protected glutamine supplementation on blood metabolites, thyroid hormones, and redox status of heat stressed fattening lambs. Iran. J. Appl. Anim. Sci. 2021, 11, 557–565. [Google Scholar]
- Wu, Q.; Xing, Z.; Liao, J.; Zhu, L.; Zhang, R.; Wang, S.; Wang, C.; Ma, Y.; Wang, Y. Effects of glutamine on rumen digestive enzymes and the barrier function of the ruminal epithelium in Hu lambs fed a high-concentrate finishing diet. Animals 2022, 12, 3418. [Google Scholar] [CrossRef] [PubMed]
- Mohamadzadeh, H.; Yansari, A.T.; Dirandeh, E. Effects of protein levels and glutamine supplementation on hematological parameters, some inflammatory and immune indicators of Afshari fattening male lambs under heat stress condition. J. Rumin. Res. 2023, 11, 91–110. [Google Scholar]
- van Keulen, P.; Khan, M.A.; Dijkstra, J.; Knol, F.; McCoard, S.A. Effect of arginine or glutamine supplementation and milk feeding allowance on small intestine development in calves. J. Dairy Sci. 2020, 103, 4754–4764. [Google Scholar] [PubMed]
- Hu, Z.Y.; Su, H.W.; Li, S.L.; Cao, Z.J. Effects of parenteral administration of glutamine on autophagy of liver cell and immune responses in weaned calves. J. Anim. Physiol. Anim. Nutr. 2013, 97, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Wickramasinghe, J.; Appuhamy, R. Effects of L-glutamine supplementation on growth, starter intake and health of early-weaned dairy Heifer calves. In Iowa State University Animal Industry Report; Iowa State University: Ames, IA, USA, 2019; pp. 1–5. [Google Scholar]
- Henson, J.N.; Bogdonoff, P.D.; Thrasher, G.W. Levels of monosodium glutamate in pig starter preference. J. Anim. Sci. 1962, 21, 999–1000. [Google Scholar]
- Torii, K.; Cagan, R.H. Biochemical studies of taste sensation. IX. Enhancement of L-[3H]glutamate binding to bovine taste papillae by 5′-ribonucleotides. Biochim. Biophys. Acta 1980, 627, 313–323. [Google Scholar] [PubMed]
- Ahangarani, M.A.; Bach, A.; Bassols, A.; Vidal, M.; Valent, D.; Ruiz-Herrera, S.; Terré, M. Performance, intestinal permeability, and metabolic profile of calves fed a milk replacer supplemented with glutamic acid. J. Dairy Sci. 2020, 103, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Grovum, W.L.; Chapman, H.W. Factors affecting the voluntary intake of food by sheep. 4. The effect of additives representing the primary tastes on sham intakes by oesophageal-fistulated sheep. Br. J. Nutr. 1988, 59, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-X. Temperature control of spermatogenesis and prospect of male contraception. Front. Biosci. (Schol. Ed.) 2010, 2, 730–755. [Google Scholar] [PubMed]
- Meza-Herrera, C.A.; Calderón-Leyva, G.; Soto-Sanchez, M.J.; Serradilla, J.M.; García-Martinez, A.; Mellado, M.; Veliz-Deras, F.G. Glutamate supply positively affects serum cholesterol concentrations without increases in total protein and urea around the onset of puberty in goats. Anim. Reprod. Sci. 2014, 147, 106–111. [Google Scholar] [PubMed]
- Meza-Herrera, C.A.; González-Velázquez, A.; Veliz-Deras, F.G.; Rodríguez-Martínez, R.; Arellano-Rodriguez, G.; Serradilla, J.M.; García-Martínez, A.; Avendaño-Reyes, L.; Macías-Cruz, U. Short-term glutamate administration positively affects the number of antral follicles and the ovulation rate in cyclic adult goats. Reprod. Biol. 2014, 14, 298–301. [Google Scholar] [CrossRef] [PubMed]
- Meza-Herrera, C.A.; Vergara-Hernández, H.P.; Paleta-Ochoa, A.; Álvarez-Ruíz, A.R.; Veliz-Deras, F.G.; Arellano Rodriguez, G.; Rosales-Nieto, C.A.; Macias-Cruz, U.; Rodriguez-Martinez, R.; Carrillo, E. Glutamate supply reactivates ovarian function while increases serum insulin and triiodothyronine concentrations in criollo x saanen-alpine yearlings’ goats during the anestrous season. Animals 2020, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- Luna-García, L.A.; Meza-Herrera, C.A.; Pérez-Marín, C.C.; Corona, R.; Luna-Orozco, J.R.; Véliz-Deras, F.G.; Delgado-Gonzalez, R.; Rodriguez-Venegas, R.; Rosales-Nieto, C.A.; Bustamante-Andrade, J.A.; et al. Goats as valuable animal model to test the targeted glutamate supplementation upon antral follicle number, ovulation rate, and LH-pulsatility. Biology 2022, 11, 1015. [Google Scholar] [CrossRef] [PubMed]
- Smith, Q.R. Transport of Glutamate and other amino acids at the blood-brain barrier. J. Nutr. 2000, 130, 1016S–1022S. [Google Scholar] [CrossRef] [PubMed]
- Acharya, S.; Petzel, E.A.; Hales, K.E.; Underwood, K.R.; Swanson, K.C.; Bailey, E.A.; Cammack, K.M.; Brake, D.W. Effects of long-term postgastric infusion of casein or glutamic acid on small intestinal starch digestion and energy balance in cattle. J. Anim. Sci. 2023, 101, skac329. [Google Scholar] [PubMed]
- Padunglerk, A.; Prasanpanich, S.; Kongmun, P. Use of monosodium glutamate by-product in cow diet on performance of lactating dairy cows. Anim. Sci. J. 2017, 88, 86–93. [Google Scholar] [CrossRef]
- Maeng, W.J.; Van Nevel, C.J.; Baldwin, R.L.; Morris, J.G. Rumen microbial growth rates and yields: Effect of amino acids and protein. J. Dairy Sci. 1976, 59, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Kajikawa, H.; Mitsumori, M.; Ohmomo, S. Stimulatory and inhibitory effects of protein amino acids on growth rate and efficiency of mixed ruminal bacteria. J. Dairy Sci. 2002, 85, 2015–2022. [Google Scholar] [CrossRef] [PubMed]
- Kajikawa, H.; Mitsumori, M.; Tajima, K.; Kurihara, M. Short communication: Amino acid antagonistic to the amino acids inhibitory for growth rate of mixed ruminal bacteria. J. Dairy Sci. 2005, 88, 2601–2603. [Google Scholar] [CrossRef] [PubMed]
- Danna, H.M.; Ballard, C.S.; Grant, R.J.; Cotanch, K.W.; Carter, M.P.; Suekawa, M. Effects of glutamate on microbial efficiency and metabolism in continuous culture of ruminal contents and on performance of mid-lactation dairy cows. Anim. Feed Sci. Technol. 2006, 130, 204–224. [Google Scholar] [CrossRef]
- Nombekela, S.W.; Murphy, M.R.; Gonyou, H.W.; Marden, J.I. Dietary preferences in early lactation cows as affected by primary tastes and some common feed flavors. J. Dairy Sci. 1994, 77, 2393–2399. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, R.; San Gabriel, A.; Wu, G. Dietary supplementation with monosodium glutamate enhances milk production by lactating sows and the growth of suckling piglets. Amino Acids 2022, 54, 1055–1068. [Google Scholar] [CrossRef]
- Calderón-Leyva, D.R.M.G.; Meza-Herrera, C.A.; Arellano-Rodriguez, G.; Gaytan-Alemán, L.R.; Alvarado-Espino, A.S.; Gonzalez-Graciano, E.A.; Delgado-Bermejo, J.V.; Véliz-Deras, F.G. Effect of glutamate supplementation upon semen quality of young seasonally sexual-inactive Dorper rams. J. Anim. Res. 2017, 7, 419–424. [Google Scholar] [CrossRef]
- Roberts, A.; Lynch, B.; Rietjens, I. Risk assessment paradigm for glutamate. Ann. Nutr. Metab. 2018, 73 (Suppl. S5), 53–64. [Google Scholar] [CrossRef]
- Meza-Herrera, C.A.; Torres-Moreno, M.; Lopez-Medrano, J.I.; Gonzalez-Bulnes, A.; Veliz, F.G.; Mellado, M.; Wurzinger, M.; Soto-Sanchez, M.J.; Calderon-Leyva, M.G. Glutamate supply positively affects serum release of triiodothronine and insulin across time without increases of glucose during the onset of puberty in female goats. Anim. Reprod. Sci. 2011, 125, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Sawant, O.B.; Ramadoss, J.; Hankins, G.D.; Wu, G.; Washburn, S.E. Effects of L-glutamine supplementation on maternal and fetal hemodynamics in gestating ewes exposed to alcohol. Amino Acids 2014, 46, 1981–1996. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Meier, S.A.; Knabe, D.A. Dietary glutamine supplementation prevents jejunal atrophy in weaned pigs. J. Nutr. 1996, 126, 2578–2584. [Google Scholar] [CrossRef]
- Dong, G.Z.; Liu, S.M.; Wu, Y.X.; Lei, C.L.; Zhou, J.; Zhang, S. Diet-induced bacterial immunogens in the gastrointestinal tract of dairy cows: Impacts on immunity and metabolism. Acta Vet. Scand. 2011, 53, 48. [Google Scholar] [CrossRef] [PubMed]
- Ametaj, B.N.; Koenig, K.M.; Dunn, S.M.; Yang, W.Z.; Zebeli, Q.; Beauchemin, K.A. Backgrounding and finishing diets are associated with inflammatory responses in feedlot steers. J. Anim. Sci. 2009, 87, 1314–1320. [Google Scholar] [CrossRef]
- Mateo, R.D.; Wu, G.; Moon, H.K.; Carroll, J.A.; Kim, S.W. Effects of dietary arginine supplementation during gestation and lactation on the performance of lactating primiparous sows and nursing piglets. J. Anim. Sci. 2008, 86, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Lobley, G.E.; Hoskin, S.O.; McNeil, C.J. Glutamine in animal science and production. J. Nutr. 2001, 131, 2525S–2531S. [Google Scholar] [CrossRef]
- Halliwell, B. Biochemistry of oxidative stress. Biochem. Soc. Trans. 2007, 35, 1147–1150. [Google Scholar] [CrossRef] [PubMed]
- Prpar, S.; Martignani, E.; Dovc, P.; Baratta, M. Identification of goat mammary stem/progenitor cells. Biol. Reprod. 2012, 86, 117. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.K.; Aikman, P.C.; Lupoli, B.; Humphries, D.J.; Beever, D.E. Splanchnic metabolism of dairy cows during the transition from late gestation through early lactation. J. Dairy Sci. 2003, 86, 1201–1217. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.K.; Kristensen, N.B. Nitrogen recycling through the gut and the nitrogen economy of ruminants: An asynchronous symbiosis. J. Anim. Sci. 2008, 86 (Suppl. S14), E293–E305. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Edwards, C.C.; Burrin, D.G.; Kristensen, N.B.; Holst, J.J.; McLeod, K.R.; Harmon, D.L. Glucagon-like peptide-2 (GLP-2) increases net amino acid utilization by the portal-drained viscera of ruminating calves. Animal 2012, 6, 1985–1997. [Google Scholar] [CrossRef] [PubMed]
- Drackley, J.K.; Blome, R.M.; Bartlett, K.S.; Bailey, K.L. Supplementation of 1% L-glutamine to milk replacer does not overcome the growth depression in calves caused by soy protein concentrate. J. Dairy Sci. 2006, 89, 1688–1693. [Google Scholar] [CrossRef] [PubMed]
- da Silva, J.T.; Manzoni, T.; Rocha, N.B.; Santos, G.; Miqueo, E.; Slanzon, G.S.; Bittar, C.M.M. Evaluation of milk replacer supplemented with lysine and methionine in combination with glutamate and glutamine in dairy calves. J. Appl. Anim. Res. 2018, 46, 960–966. [Google Scholar] [CrossRef]
- Kidd, M.T.; Maynard, C.W.; Mullenix, G.J. Progress of amino acid nutrition for diet protein reduction in poultry. J. Anim. Sci. Biotechnol. 2021, 12, 45. [Google Scholar] [CrossRef] [PubMed]
| Variable | Adult Sheep (60 kg BW) | |
|---|---|---|
| Glutamate | Glutamine | |
| Feed intake (on an as-fed basis), a kg/day | 1.20 | 1.20 |
| Crude protein intake, g/day | 140 | 140 |
| Microbial and feed proteins entering the duodenum, b g/day | 95.2 | 95.2 |
| Nitrogen leaving the rumen as ammonia and reentering the rumen via N recycling for microbial protein synthesis, c g/day | 18.2 | 18.2 |
| Total microbial and feed proteins in the small intestine, g/day | 113.4 | 113.4 |
| Digestible microbial and feed proteins in the small intestine, d g/day | 96.4 | 96.4 |
| Glu or Gln released from digestible microbial and feed protein, e g/day | 7.71 | 5.30 |
| Glu or Gln entering the portal vein (100% extraction by the small intestine), g/day | 0 | 0 |
| Glu or Gln catabolism in the whole body, g/day | 51.5 f | 46.7 g |
| Glu or Gln required for protein deposition (maintenance), g/day | 0 | 0 |
| Glu or Gln oxidation in the portal-drained viscera, g/day | 7.71 | 5.30 |
| Endogenous synthesis of Glu and Gln, h g/day | 43.8 | 41.4 |
| % of dietary crude protein used for Glu and Gln synthesis i | 31.3 | 29.6 |
| Amino Acid | Net uptake (+) or Net Release (–) by the Liver | Amino Acid | Net Uptake (+) by the Liver |
|---|---|---|---|
| Ala | +11 | Leu | +2.1 |
| Arg | +4.9 | Lys | +2.2 |
| Asn | +17 | Met | +9.9 |
| Asp | +2.4 | Phe | +12 |
| Citrulline | +0.1 | Pro | +6.5 |
| Gln | +5.7 | Ser | +19 |
| Glu | –24 | Thr | +5.3 |
| Gly | +15 | Trp | +4.2 |
| His | +4.8 | Tyr | +11 |
| Ile | +1.9 | Val | +1.3 |
| Animals | Production of Metabolites from Glutamine (nmol/mg DNA/30 min) | |||
|---|---|---|---|---|
| CO2 | Ornithine | Citrulline | Arginine | |
| Brahman cattle (n = 5 per age group) | ||||
| 2-day-old calves | 3903 ± 228 a | 15.8 ± 1.4 a | 149 ± 13 a | 185 ± 20 a |
| 7-day-old calves | 3725 ± 301 a | 15.0 ± 1.2 a | 132 ± 16 a | 169 ± 22 a |
| 6-month-old calves | 906 ± 54 b | 5.13 ± 0.09 b | 44.7 ± 2.5 b | 9.22 ± 0.61 b |
| 24-month-old beef cattle | 410 ± 291 c | 3.76 ± 0.07 c | 30.2 ± 1.1 c | 2.30 ± 0.14 c |
| Suffolk sheep (n = 6 per age group) | ||||
| 0-day-old lambs | 4582 ± 170 a | 21.4 ± 0.79 a | 205 ± 9.0 a | 390 ± 13 a |
| 3-month-old lambs | 1359 ± 51 b | 10.6 ± 0.35 b | 86.8 ± 3.8 b | 40.2 ± 1.2 b |
| 24-month-old (non-pregnant and non-lactating) ewes | 406 ± 16 d | 6.12 ± 0.19 d | 39.0 ± 1.3 d | 2.84 ± 0.15 d |
| 24-month-old pregnant ewes | 493 ± 19 c | 7.56 ± 0.22 c | 48.7 ± 1.6 c | 3.72 ± 0.20 c |
| Animal | Basal Diet | Administration of L-Glutamate | Responses to L-Glutamate | Reference |
|---|---|---|---|---|
| Steer calves | A semi-purified diet with 97% of nitrogen from urea | Dietary supplementation with 0 or 3% Glu for 21 days | ↑ ruminal production of fatty acids with ≥4 carbons | Oltjen et al. [122] |
| Dairy valves | Milk replacer with 24.8% CP and 19.1% fat | Dietary supplementation with 0 or 0.3% Glu (on a DM basis) for 8 weeks between 3 and 59 days of age | No changes in feed intake, weight gain, or plasma concentrations of urea and proteinogenic AAs | Ahangarani et al. [148] |
| Dairy calves | Fresh whole milk (2.3 kg twice daily) plus a starter diet (0 to 21 days of age); alfalfa-based diet after weaning (21 days of age); 18% CP in solid diets | Either 0 or 0.2% MSG between 0 and 21 days of age; and between day 21 (weaning) and day 84 of age | ↑ 321% for solid feed intake in week 3 of age; ↑ 248%, 107%, and 40% feed intake in weeks 1, 2, and 3 postweaning, respectively | Waldern and van Dyk [123] |
| Dairy calves | Same as above, except that the weaning age was 28 days, but not 21 days | Either 0 or 0.2% MSG between 0 and 28 days of age; and between day 28 (weaning) and day 42 of age | ↓ weight loss (91 vs. 45 g per day in the first week after weaning | Waldern and van Dyk [123] |
| Growing steers | Soybean hull-based diets; BW of cattle = 259 kg | Duodenal infusions of either 0 or 133 g Glu and 1391 g raw cornstarch per day for 6 days | ↑ starch digestion in the small intestine | Brake et al. [76] |
| Growing steers | Soybean hull-based diets; BW of cattle = 179 kg | Duodenal infusions of either 0 or 121 g Glu and 1460 g raw cornstarch per day for 42 days | No effect on starch digestion in the small intestine | Acharya et al. [156] |
| Neonatal lambs | A diet consisting of lucerne and concentrates | Either 0 or 0.5% MSG beginning at 18 days of age | ↑ feed intake and weight gain between 90 and 135 days of age | Galgan and Russell [124] |
| Adult sheep | Ammoniated barley straw; or pelleted lucerne | Either 0 or 1% MSG for 64 days; or 0.5% and 4% MSG for 64 days | ↑ feed intake by 10%, 16%, or 40%, respectively | Colucci and Grovum [125] |
| Rams | Alfalfa hay and corn-based diet | im administration of Glu (0 or 7 mg/kg BW every 3 days) for 28 days | ↑ sperm production by 51%; ↑ GnRH secretion | Meza-Herrera [126] |
| Female goats | A mixed diet b | im or iv administration of Glu (0 or 7 mg/kg BW twice weekly) From mid-June to late October | ↑ the onset of puberty without affecting BW | Torres-Moreno et al. [127] |
| Lactating cows | Soybean meal (25%)-based diet | Dietary supplementation with 0, 0.24, 0.48, or 0.72 % Glu as 5, 10, and 15% MSG by-product, respectively | ↑ milk production by 15, 22, and 33%, respectively | Nakanishi et al. [128] |
| Sheep with heat stress | Total mixed ration with 14.7% CP | Dietary supplementation with 0 or 3 g Glu/head/day for 90 days | ↑ rumen fermentation; ↑ antioxidative capacity; ↑ growth performance | Li et al. [129] |
| Lactating cows | Total mixed ration with 16.5% CP (prepartum) and 18.3% CP (postpartum) | Dietary supplementation with 0 or 1.54% Glu (prepartum) plus 1.14% Glu (Postpartum) as MSG | ↑ feed intake; ↑ digestive capacity; ↓ losses of whole-body fat and protein after calving | Hisadomi et al. [130] |
| Female goats | Total mixed ration | Dietary supplementation with 0 or 1 g MSG/kg BW for 23 days | ↑ feed intake; ↑ ruminal movement; ↑ rumination; ↑ ovarian follicle number; ↑ intraovarian blood flow | Soares et al. [131] |
| Animal | Basal Diet | Administration of L-Glutamine | Responses to L-Glutamine | Reference |
|---|---|---|---|---|
| Healthy calves | Grain-based diet with 21.25% CP | iv infusion of Gln (0 or 16 g/day) for 14 days between 35 and 49 days of age | ↑ villus height and crypt depth in the duodenum, Jejunum, and ileum; ↑ plasma glucose Conc. | Hu et al. [144] |
| Healthy calves | Liquid milk at 28–34 days of age; grain- based diet between day 35 (weaning) and day 42 of age | Dietary supplementation with 0 or 2% Gln (on a DM basis) for one week both before and after weaning | ↑ daily weight gain; ↑ adaptation to weaning ration; ↑ hip width and body length | Wickramasinghe and Appuhamy [145] |
| Healthy calves | Reconstituted whole milk with 26% CP and 26% fat (on a DM basis); daily feed intake at 10 or 20% of initial BW | Dietary supplementation with 0 or 1% Gln (on a DM basis) for 35 days between 4 and 39 days of age | ↑ villus height, width, and surface area in duodenum and jejunum of calves with high-level feed intake; no effect in calves with low-level feed intake | van Keulen et al. [143] |
| Calves with diarrhea | Cow’s milk | Oral electrolyte solution with 0 or 40 mM Gln | ↑ morphology of the SI; ↑ hydration status | Naylor et al. [132] |
| Calves with diarrhea | Cow’s milk | Oral electrolyte solution with no or high Gln concentration | ↑ hydration status; ↑ recovery from diarrhea; ↓ weight loss by 2.3–2.6 kg | Brooks et al. [133] |
| Calves with diarrhea | Cow’s milk | Oral electrolyte solution with no or high Gln concentration | ↑ morphology of the SI; ↑ recovery from diarrhea | Brooks et al. [134] |
| Calves with diarrhea | Cow’s milk | Oral electrolyte solution with no or high Gln concentration | ↑ hydration status; ↑ recovery from diarrhea | Pal and Pachauri [135] |
| Beef cattle (feed-lot) | Grain-based diet | Dietary supplementation with 0 or 1% Gln for 25 days | ↑ clearance of endotoxin from the blood | Jin et al. [136] |
| Dairy cows | Concentrates-based diet with 18% CP | Intra-abomasal infusion of Gln (0 or 300 g/day) for 21 days | ↑ milk yield by 3% | Doepel et al. [87] |
| Dairy cows | Concentrates-based diet with 18% CP | iv infusion of Gln (0, 106, or 212 g/8 h per day) for 7 days | ↑ production of acute-phase protein by the liver | Jafari et al. [70] |
| Dairy cows | Concentrates-based diet with 18% CP | Rumen-protected Gln (0, 160, or 320 g/day) for 60 days | ↑ immune response; ↑ anti-oxidative capacity | Caroprese et al. [137] |
| Sheep with trickle infection | Roughage-based diet | Daily intra-abomasal infusion of Gln (0 or 5 g) and cysteine (0 or 1 g) for 12 weeks | ↑ N retention in the body; ↓ systemic infection | Hoskin et al. [69] |
| Gestating sheep | Roughage-based diet with 12% CP | iv infusion of Gln (0 or 100 mg/kg BW 3 times daily) for 23 days to ewes with alcohol-induced acidosis | Prevents alcohol-induced acidosis in fetus; prevents alcohol-induced increase in fetal brain blood flow | Washburn et al. [138] |
| Gestating sheep | Roughage-based diet with 12% CP | iv infusion of Gln (0 or 100 mg/kg BW 3 times daily) to ewes with alcohol-induced acidosis between 109 and 132 days of gestation | Prevent alcohol-induced acid-base imbalance and alcohol-induced IUGR | Sawant et al. [72,167] |
| Goats with heat stress | Corn offals and hays | Oral Gln (0 or 0.2 g/kg BW, once daily) for 21 days | ↓ rectal temperature; ↓ erythrocyte fragility; ↓ oxidative stress | Ocheja et al. [68] |
| Lactating cows | Total mixed ration with 16.3% CP | Dietary supplementation with rumen-protected Gln (0, 250 or 350 g/cow/day) for 21 days | ↑ DM intake; ↑ milk yield; ↑ plasma levels of glucose, and proteins; ↓ plasma BHB and NEFA; ↓ SCC in milk | Nemati et al. [139] |
| Lactating cows | Total mixed ration with 16.3% CP | Dietary supplementation with rumen-protected Gln (0 or 150 g/cow/day) for 21 days | NC for DM intake or milk yield; ↑ plasma levels of glucose and proteins; ↓ plasma BHB and NEFA; ↓ SCC in milk | Nemati et al. [139] |
| Lambs with heat stress | Total mixed rations with 13.4–14.5% CP | Dietary supplementation with rumen-protected Gln (0 or 0.2 g/kg BW per day) for 45 days | ↑ antioxidative capacity; ↓ cortisol in serum; ↓ systemic NO production | Feyz et al. [140] |
| Lambs (healthy) | High-concentrate diet with 13.3% CP | Dietary supplementation with 0, 0.5 or 1% Gln for 60 days | ↑ expression of claudin-1 and interleukin-10 in the ruminal epithelium; ↑ Ac/Prop ratio and lipase activity in ruminal fluid | Wu et al. [141] |
| Lambs with heat stress | Total mixed rations with 13.4–14.5% CP | Dietary supplementation with rumen-protected Gln (0 or 0.2 g/kg BW per day) for 45 days | ↑ antioxidative capacity, as indicated by reduced | Mohamadzadeh et al. [142] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, G.; Bazer, F.W.; Johnson, G.A.; Satterfield, M.C.; Washburn, S.E. Metabolism and Nutrition of L-Glutamate and L-Glutamine in Ruminants. Animals 2024, 14, 1788. https://doi.org/10.3390/ani14121788
Wu G, Bazer FW, Johnson GA, Satterfield MC, Washburn SE. Metabolism and Nutrition of L-Glutamate and L-Glutamine in Ruminants. Animals. 2024; 14(12):1788. https://doi.org/10.3390/ani14121788
Chicago/Turabian StyleWu, Guoyao, Fuller W. Bazer, Gregory A. Johnson, M. Carey Satterfield, and Shannon E. Washburn. 2024. "Metabolism and Nutrition of L-Glutamate and L-Glutamine in Ruminants" Animals 14, no. 12: 1788. https://doi.org/10.3390/ani14121788