

Outbreak of Chlamydia psittaci Infection in a Commercial Psittacine Breeding Aviary in Argentina
 , , , , , , , and
, , , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Case Presentation
2.2. Necropsy and Ancillary Tests
2.3. Microbiology
2.4. Histopathology
2.5. Inmunohistochemistry
2.6. Chlamydia Isolation Procedure
2.7. Polymerase Chain Reaction
3. Results
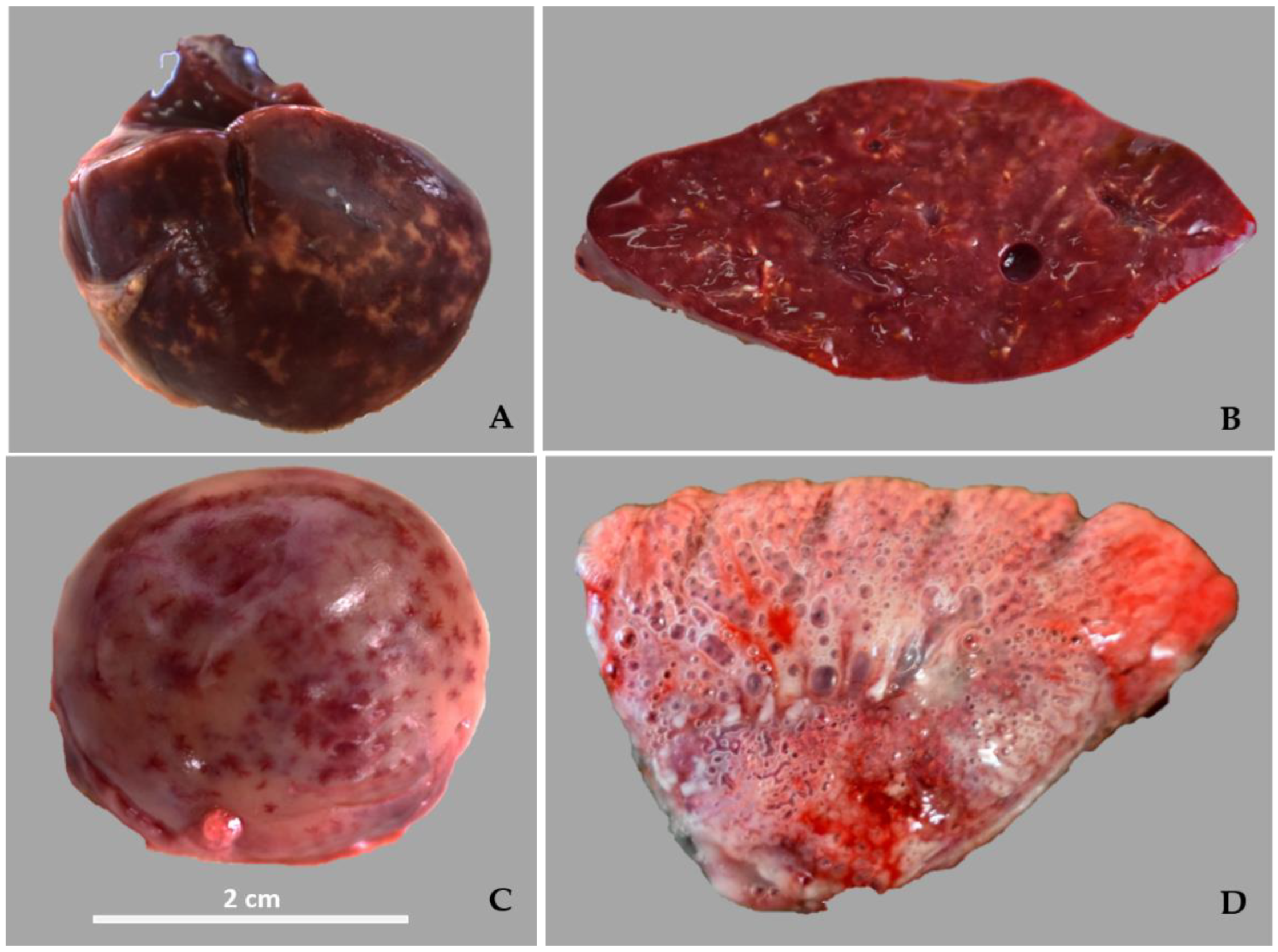
3.1. Necropsy
3.2. Microbiology
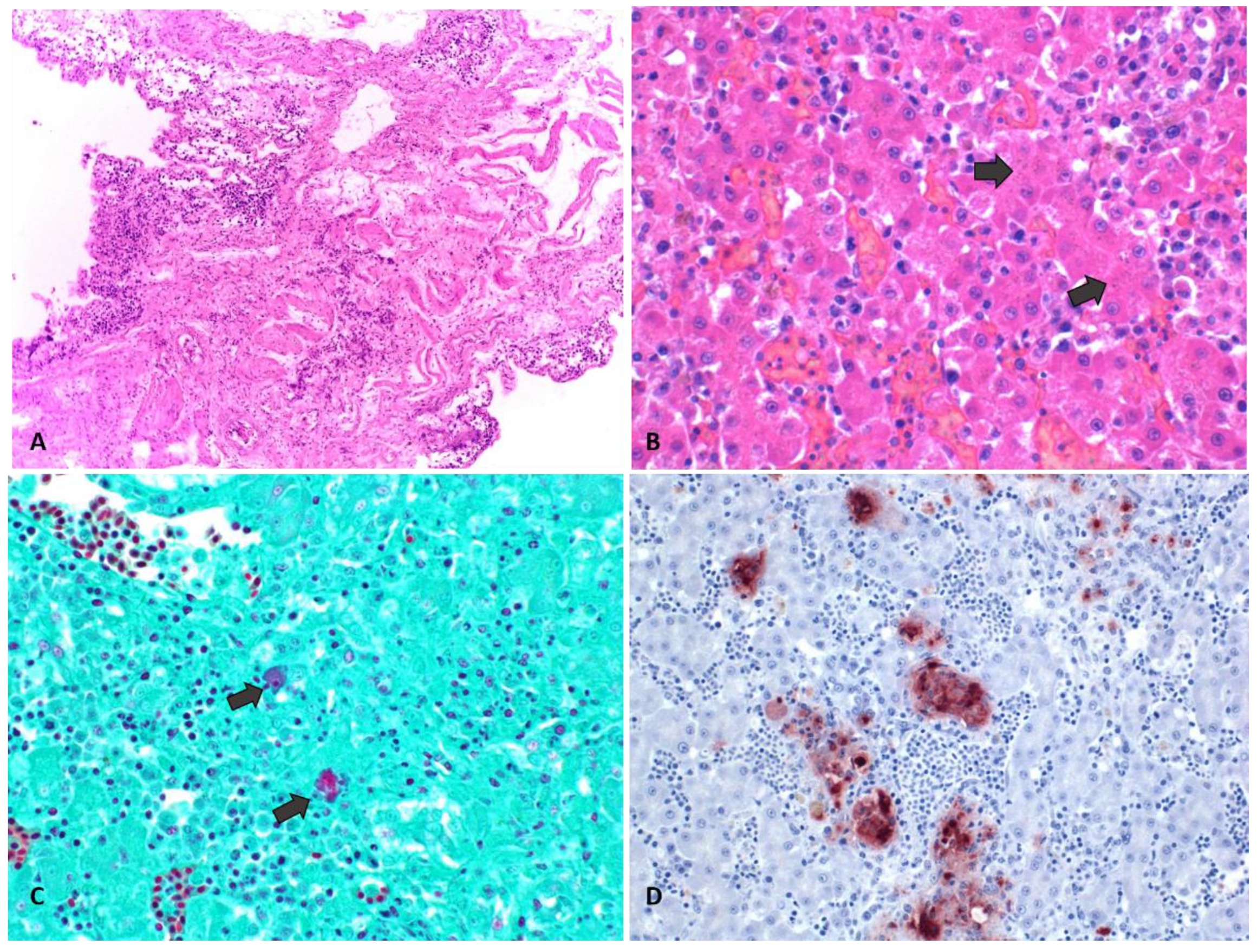
3.3. Histology
3.4. Immunohistochemistry
3.5. Chlamydia Isolation
3.6. Polymerase Chain Reaction
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stokes, H.S.; Berg, M.L.; Bennett, A.T.D. A Review of Chlamydial infections in wild birds. Pathogens 2021, 10, 948. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, K.; Anbazhagan, S.; Karthik, K.; Angappan, M.; Dhayananth, B. A comprehensive review on avian Chlamydiosis: A neglected zoonotic disease. Trop. Anim. Health Prod. 2021, 53, 414. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.; Jelocnik, M.; Onizawa, E.; McNally, J.; Coilparampil, R.; Pinczowski, P.; Bogema, D.; Westermann, T. Chlamydia pecorum ovine abortion: Associations between maternal infection and perinatal mortality. Pathogens 2021, 10, 1367. [Google Scholar] [CrossRef] [PubMed]
- Marti, H.; Jelocnik, M. Animal Chlamydiae: A concern for human and veterinary medicine. Pathogens 2022, 11, 364. [Google Scholar] [CrossRef] [PubMed]
- Vanrompay, D.; Harkinezhad, T.; van de Walle, M.; Beeckman, D.; van Droogenbroeck, C.; Verminnen, K.; Leten, R.; Martel, A.; Cauwerts, K. Chlamydophila psittaci transmission from pet birds to humans. Emerg. Infect. Dis. 2007, 13, 1108–1110. [Google Scholar] [CrossRef] [PubMed]
- Branley, J.M.; Weston, K.M.; England, J.; Dwyer, D.E.; Sorrell, T.C. Clinical features of endemic community-acquired Psittacosis. New Microbes New Infect. 2014, 2, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Telfer, B.L.; Moberley, S.A.; Hort, K.P.; Branley, J.M.; Dwyer, D.E.; Muscatello, D.J.; Correll, P.K.; England, J.; McAnulty, J.M. Probable Psittacosis outbreak linked to wild birds. Emerg. Infect. Dis. 2005, 11, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Balsamo, G.; Maxted, A.M.; Midla, J.W.; Murphy, J.M.; Wohrle, R.; Edling, T.M.; Fish, P.H.; Flammer, K.; Hyde, D.; Kutty, P.K.; et al. Compendium of measures to control Chlamydia psittaci infection among humans (Psittacosis) and pet birds (Avian Chlamydiosis), 2017. J. Avian Med. Surg. 2017, 31, 262–282. [Google Scholar] [CrossRef] [PubMed]
- Dai, N.; Li, Q.; Geng, J.; Guo, W.; Yan, W. Severe pneumonia caused by Chlamydia psittaci: Report of two cases and literature review. J. Infect. Dev. Ctries. 2022, 16, 1101–1112. [Google Scholar] [CrossRef]
- Kalmar, I.D.; Dicxk, V.; Dossche, L.; Vanrompay, D. Zoonotic infection with Chlamydia psittaci at an avian refuge centre. Vet. J. 2014, 199, 300–302. [Google Scholar] [CrossRef]
- Szymańska-Czerwińska, M.; Niemczuk, K. Avian Chlamydiosis zoonotic disease. Vector-Borne Zoonotic Dis. 2016, 16, 1–3. [Google Scholar] [CrossRef]
- Kaleta, E.F.; Taday, E.M.A. Avian host range of Chlamydophila spp. based on isolation, antigen detection and serology. Avian Pathol. J. WVPA 2003, 32, 435–461. [Google Scholar] [CrossRef] [PubMed]
- Markey, B.K. Clinical Veterinary Microbiology, 2nd ed.; Elsevier: Edinburgh, Scotland, 2013; ISBN 978-0-7234-3237-1. [Google Scholar]
- Suwa, T.; Touchi, A.; Hirai, K.; Itakura, C. Pathological studies on Chlamydiosis in parakeets (Psittacula krameri manillensis). Avian Pathol. J. WVPA 1990, 19, 355–369. [Google Scholar] [CrossRef] [PubMed]
- Vanrompay, D.; Ducatelle, R.; Haesbrouck, F. Pathology of experimental chlamydiosis in turkeys. Vlaams Diergeneeskd. Tijdschr. 1995, 64, 19–24. [Google Scholar]
- Van Buuren, C.E.; Dorrestein, G.M.; Van Dijk, J.E. Chlamydia psittaci infections in birds: A review on the pathogenesis and histopathological features. Vet. Q. 1994, 16, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Burt, S.A.; Röring, R.E.; Heijne, M. Chlamydia psittaci and C. avium in feral pigeon (Columba livia domestica) droppings in two cities in the Netherlands. Vet. Q. 2018, 38, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, K.; Wójcik-Fatla, A. Chlamydia psittaci in faecal samples of feral pigeons (Columba livia forma urbana) in urban areas of Lublin City, Poland. Curr. Microbiol. 2022, 79, 367. [Google Scholar] [CrossRef] [PubMed]
- Perez-Sancho, M.; García-Seco, T.; Porrero, C.; García, N.; Gomez-Barrero, S.; Cámara, J.M.; Domínguez, L.; Álvarez, J. A Ten-year-surveillance program of zoonotic pathogens in feral pigeons in the city of Madrid (2005–2014): The importance of a systematic pest control. Res. Vet. Sci. 2020, 128, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Sukon, P.; Nam, N.H.; Kittipreeya, P.; Sara-In, A.; Wawilai, P.; Inchuai, R.; Weerakhun, S. Global Prevalence of Chlamydial Infections in Birds: A systematic review and meta-analysis. Prev. Vet. Med. 2021, 192, 105370. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Cao, K.; Wei, Y.; Qian, Y.; Liang, J.; Dong, D.; Tang, J.; Zhu, Z.; Gu, Q.; Yu, W. Metagenomic next-generation sequencing in the diagnosis of severe pneumonias caused by Chlamydia psittaci. Infection 2020, 48, 535–542. [Google Scholar] [CrossRef]
- Rybarczyk, J.; Versteele, C.; Lernout, T.; Vanrompay, D. Human psittacosis: A review with emphasis on surveillance in Belgium. Acta Clin. Belg. 2020, 75, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Villagra, S.M. Brote de psitacosis durante la pandemia de COVID-19 en la provincia de santa Fe. Serie de casoS. Rev. Argent. Med. 2022, 10, 139–142. [Google Scholar]
- Szymańska-Czerwińska, M.; Mitura, A.; Niemczuk, K.; Zaręba, K.; Jodełko, A.; Pluta, A.; Scharf, S.; Vitek, B.; Aaziz, R.; Vorimore, F.; et al. Dissemination and genetic diversity of Chlamydial agents in polish wildfowl: Isolation and molecular characterisation of avian Chlamydia abortus strains. PLoS ONE 2017, 12, e0174599. [Google Scholar] [CrossRef] [PubMed]
- Mattmann, P.; Marti, H.; Borel, N.; Jelocnik, M.; Albini, S.; Vogler, B.R. Chlamydiaceae in wild, feral and domestic pigeons in Switzerland and insight into population dynamics by Chlamydia psittaci multilocus sequence typing. PLoS ONE 2019, 14, e0226088. [Google Scholar] [CrossRef] [PubMed]
- Origlia, J.A.; Cadario, M.E.; Frutos, M.C.; Lopez, N.F.; Corva, S.; Unzaga, M.F.; Piscopo, M.V.; Cuffini, C.; Petruccelli, M.A. Detection and molecular characterization of Chlamydia psittaci and Chlamydia abortus in psittacine pet birds in Buenos Aires Province, Argentina. Rev. Argent. Microbiol. 2019, 51, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Harkinezhad, T.; Verminnen, K.; Van Droogenbroeck, C.; Vanrompay, D. Chlamydophila psittaci Genotype E/B transmission from african grey parrots to humans. J. Med. Microbiol. 2007, 56, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Barrow, G.I.; Feltham, R.K.A. Cowan and Steel’s Manual for the Identification of Medical Bacteria, 3rd ed.; Cambridge University Press: Cambridge, UK, 1993; ISBN 978-0-521-54328-6. [Google Scholar]
- Laroucau, K.; Vorimore, F.; Aaziz, R.; Solmonson, L.; Hsia, R.C.; Bavoil, P.M.; Fach, P.; Hölzer, M.; Wuenschmann, A.; Sachse, K. Chlamydia buteonis, a new Chlamydia species isolated from a red-shouldered hawk. Syst. Appl. Microbiol. 2019, 42, 125997. [Google Scholar] [CrossRef]
- Pantchev, A.; Sting, R.; Bauerfeind, R.; Tyczka, J.; Sachse, K. New Real-Time PCR Tests for species-specific detection of Chlamydophila psittaci and Chlamydophila abortus from tissue samples. Vet. J. 2009, 181, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Katoh, H.; Ohya, K.; Fukushi, H. Development of Novel Real-Time PCR Assays for detecting DNA virus infections in Psittaciform birds. J. Virol. Methods 2008, 154, 92–98. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Pannekoek, Y.; Dickx, V.; Beeckman, D.S.A.; Jolley, K.A.; Keijzers, W.C.; Vretou, E.; Maiden, M.C.J.; Vanrompay, D.; van der Ende, A. Multi locus sequence typing of Chlamydia reveals an association between Chlamydia psittaci genotypes and host species. PLoS ONE 2010, 5, e14179. [Google Scholar] [CrossRef] [PubMed]
- Jelocnik, M.; Polkinghorne, A.; Pannekoek, Y. Multilocus Sequence typing (MLST) of Chlamydiales. Methods Mol. Biol. 2019, 2042, 69–86. [Google Scholar] [CrossRef]
- Hopkins, J.; Alsop, J.; Varga, C.; Pasma, T.; Jekel, P.; Rishi, L.; Hirji, M.; Filejski, C. Investigation and management of Psittacosis in a public aviary: A One Health approach. Can. Commun. Dis. Rep. 2016, 42, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Hogerwerf, L.; Roof, I.; de Jong, M.J.K.; Dijkstra, F.; van der Hoek, W. Animal sources for zoonotic transmission of Psittacosis: A systematic review. BMC Infect. Dis. 2020, 20, 192. [Google Scholar] [CrossRef]
- Maza, Y.; Chaparro, M.; Argañaráz, C.; Genero, S. Brote de Psitacosis en la localidad de Fontana (Chaco, Argentina) Durante Enero de 2014. Rev. Vet. 2016, 27, 45–47. [Google Scholar] [CrossRef]
- Mariño, B.; Sciabarrasi, A.; Imhoberdoff, P.; Talamé, M.P.; Frutos, M.C.; Mosmann, J.; Cuffini, C. Detección de Chlamydia psittaci en muestras de pichones de loro hablador (Amazona aestiva) incautadas en Santa Fe, Argentina. Rev. Fac. Cienc. Médicas Córdoba 2022, 79. Available online: https://revistas.unc.edu.ar/index.php/med/article/view/39023 (accessed on 30 May 2024).
- de Chazal, L.E. Brote de psitacosis en San Miguel de Tucumán y Gran San Miguel. Rev. Med. Tucumán 2007, 12, 30–34. [Google Scholar]
- Frutos, M.C.; Monetti, M.; Kiguen, X.; Venezuela, F.; Ré, V.; Cuffini, C. Genotyping of C. psittaci in central area of Argentina. Diagn. Microbiol. Infect. Dis. 2012, 74, 320–322. [Google Scholar] [CrossRef]
- Beeckman, D.S.A.; Vanrompay, D.C.G. Zoonotic Chlamydophila psittaci infections from a clinical perspective. Clin. Microbiol. Infect. 2009, 15, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Andersen, A.A.; Vanrompay, D. Avian Chlamydiosis. In Diseases of Poultry; Blackwell: Hoboken, NJ, USA, 2008; pp. 971–986. ISBN 978-0-8138-0718-8. [Google Scholar]
- Muroni, G.; Pinna, L.; Serra, E.; Chisu, V.; Mandas, D.; Coccollone, A.; Liciardi, M.; Masala, G. A Chlamydia psittaci outbreak in Psittacine birds in Sardinia, Italy. Int. J. Environ. Res. Public. Health 2022, 19, 14204. [Google Scholar] [CrossRef]
- Vanrompay, D.; Butaye, P. Characterization of avian Chlamydia psittaci strains using Ompl restriction mapping and serovar-specific monoclonal antibodies. Res. Microbiol. 1997, 148, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Ehricht, R.; Slickers, P.; Goellner, S.; Hotzel, H.; Sachse, K. Optimized DNA Microarray assay allows detection and genotyping of single PCR-amplifiable target copies. Mol. Cell. Probes 2006, 20, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.A.; Bradley, K.K.; Stobierski, M.G.; Tengelsen, L.A. Compendium of measures to control Chlamydophila psittaci (Formerly Chlamydia psittaci) infection among humans (Psittacosis) and pet birds, 2005. J. Am. Vet. Med. Assoc. 2005, 226, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Heddema, E.R.; Van Hannen, E.J.; Bongaerts, M.; Dijkstra, F.; Ten Hove, R.J.; De Wever, B.; Vanrompay, D. Typing of Chlamydia psittaci to monitor epidemiology of Psittacosis and aid disease control in the Netherlands, 2008 to 2013. Eurosurveillance 2015, 20, 21026. [Google Scholar] [CrossRef] [PubMed]
- Cadario, M.E.; Frutos, M.C.; Arias, M.B.; Origlia, J.A.; Zelaya, V.; Madariaga, M.J.; Lara, C.S.; Ré, V.; Cuffini, C.G. Epidemiological and molecular characteristics of Chlamydia psittaci from 8 human cases of Psittacosis and 4 related birds in Argentina. Rev. Argent. Microbiol. 2017, 49, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Kasimov, V.; White, R.T.; Foxwell, J.; Jenkins, C.; Gedye, K.; Pannekoek, Y.; Jelocnik, M. Whole-Genome sequencing of Chlamydia psittaci from Australasian avian hosts: A Genomics Approach to a Pathogen That Still Ruffles Feathers. Microb. Genom. 2023, 9, 001072. [Google Scholar] [CrossRef] [PubMed]
- Harkinezhad, T.; Geens, T.; Vanrompay, D. Chlamydophila psittaci infections in birds: A Review with emphasis on zoonotic consequences. Vet. Microbiol. 2009, 135, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.; Li, Y.; Pan, H.; Hui, T.; Yu, Z.; Wu, H.; Zhang, D.; Zheng, W.; Wang, S.; Zhou, Z.; et al. Atypical pneumonia caused by Chlamydia psittaci during the COVID-19 pandemic. Int. J. Infect. Dis. 2022, 122, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Liang, J.; Lao, X.; Xia, X.; Liang, J. Pneumonia caused by Chlamydia psittaci and SARS-cov-2 Coinfection Diagnosed using metagenomic next-generation sequencing: A case report. Int. Med. Case Rep. J. 2024, 17, 187–194. [Google Scholar] [CrossRef]
- Bommana, S.; Polkinghorne, A. Mini Review: Antimicrobial control of Chlamydial infections in animals: Current practices and issues. Front. Microbiol. 2019, 10, 113. [Google Scholar] [CrossRef]
- Prohl, A.; Lohr, M.; Ostermann, C.; Liebler-Tenorio, E.; Berndt, A.; Schroedl, W.; Rothe, M.; Schubert, E.; Sachse, K.; Reinhold, P. Enrofloxacin and macrolides alone or in combination with rifampicin as antimicrobial treatment in a bovine model of acute Chlamydia psittaci infection. PLoS ONE 2015, 10, e0119736. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Lesion Observed | P1 | P2 | P3 | P4 | C 1 | C 2 | Green-and-Red Macaw | Scarlet Macaw |
|---|---|---|---|---|---|---|---|---|
| Multifocal lymphoplasmacytic, heterophilic, and necrotizing splenitis with intracytoplasmic elementary bodies | ||||||||
| Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes | |
| Pulmonary congestion with edema | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Multifocal, lymphoplasmacytic, and necrotizing hepatitis with intracytoplasmic elementary bodies | ||||||||
| No | Yes | No | No | Yes | No | No | Yes | |
| Multifocal necro-heterophilic hepatitis | No | No | No | Yes | No | Yes | No | No |
| Multifocal lymphoplasmacytic, heterophilic airsacculitis | Yes | Yes | No | No | No | Yes | Yes | Yes |
| Multifocal lymphoplasmacytic nephritis | No | No | No | No | No | No | Yes | No |
| Heterophilic pneumonia | No | No | No | No | Yes | No | No | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riccio, M.B.; García, J.P.; Chiapparrone, M.L.; Cantón, J.; Cacciato, C.; Origlia, J.A.; Cadario, M.E.; Diab, S.S.; Uzal, F.A. Outbreak of Chlamydia psittaci Infection in a Commercial Psittacine Breeding Aviary in Argentina. Animals 2024, 14, 1959. https://doi.org/10.3390/ani14131959
Riccio MB, García JP, Chiapparrone ML, Cantón J, Cacciato C, Origlia JA, Cadario ME, Diab SS, Uzal FA. Outbreak of Chlamydia psittaci Infection in a Commercial Psittacine Breeding Aviary in Argentina. Animals. 2024; 14(13):1959. https://doi.org/10.3390/ani14131959
Chicago/Turabian StyleRiccio, María Belén, Jorge Pablo García, María Laura Chiapparrone, Juliana Cantón, Claudio Cacciato, Javier Anibal Origlia, María Estela Cadario, Santiago Sain Diab, and Francisco Alejandro Uzal. 2024. "Outbreak of Chlamydia psittaci Infection in a Commercial Psittacine Breeding Aviary in Argentina" Animals 14, no. 13: 1959. https://doi.org/10.3390/ani14131959