
Cory’s Shearwater (Calonectris borealis): Exploring Normal Head Anatomy through Cross-Sectional Anatomy, Computed Tomography and Magnetic Resonance Imaging
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. CT Technique
2.3. MRI Technique
2.4. Macroscopic Anatomical Sections
2.5. Anatomic Evaluation
3. Result
3.1. Anatomical Sections
3.2. Computed Tomography (CT)
3.3. Magnetic Resonance Imaging (MRI)



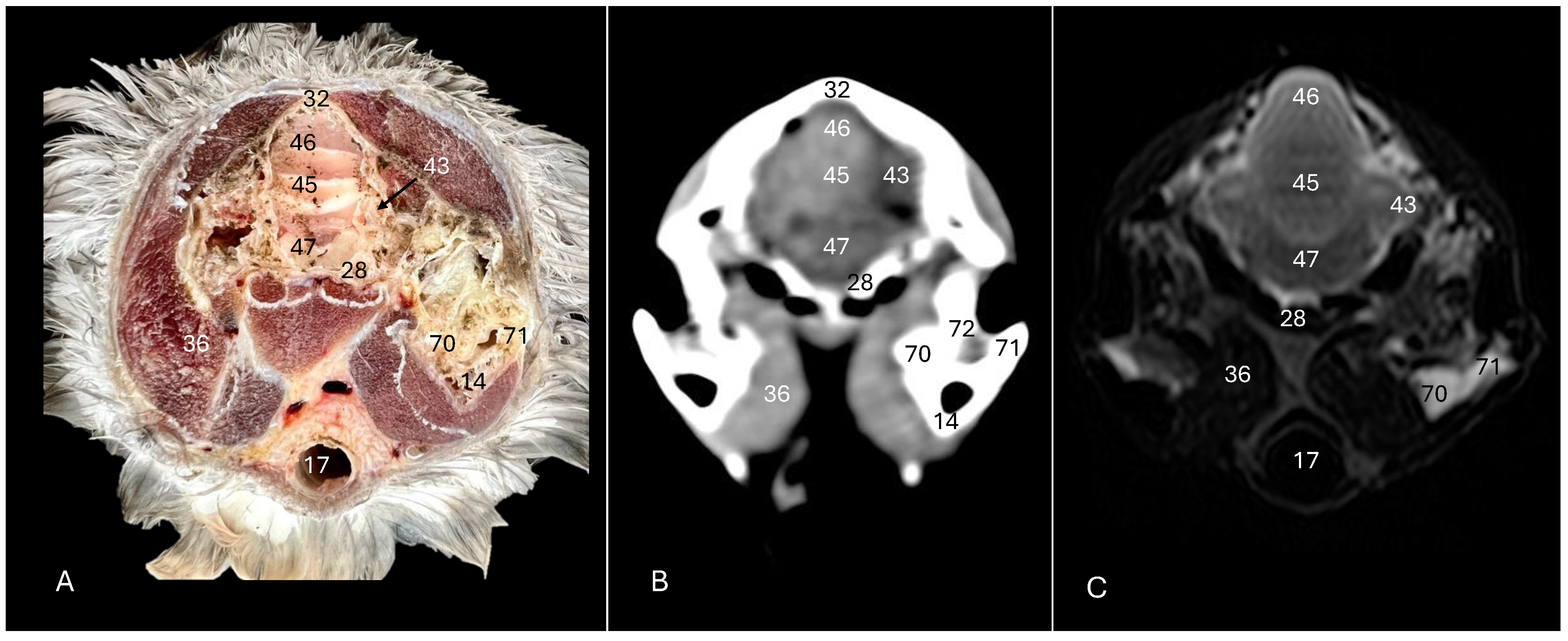





4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cory’s Shearwater—The Sound Approach. Available online: https://soundapproach.co.uk/species/corys-shearwater/ (accessed on 1 January 2024).
- Seabird of the Month—Cory’s Shearwater (Calonectris Borealis). Available online: https://www.birdlife.org/news/2021/05/31/seabird-month-corys-shearwater-calonectris-borealis/ (accessed on 1 January 2024).
- Madroño, A.; González, C.; Atienza, J.C. (Eds.) Libro Rojo de las Aves de España. In Dirección General para la Biodiversidad; SEO-BirdLife: Madrid, Spain, 2004. [Google Scholar]
- Migración y Ecología Espacial de las Poblaciones Españolas de Pardela Cenicienta. SEO BirdLife: Madrid, Spain, 2017. [CrossRef]
- La Pardela Cenicienta Elegida “Ave del año 2013”|Natura|elmundo.es. Available online: https://www.elmundo.es/elmundo/2013/03/21/natura/1363873461.html (accessed on 1 January 2024).
- Pardela Cenicienta, Especie Clave para la Conservación de los Mares-SEO/BirdLife. Available online: https://seo.org/pardela-cenicienta-especie-clave-para-la-conservacion-de-los-mares/ (accessed on 1 January 2024).
- Lauridsen, H.; Hansen, K.; Wang, T.; Agger, P.; Andersen, J.L.; Knudsen, P.S.; Rasmussen, A.S.; Uhrenholt, L.; Pedersen, M. Inside Out: Modern Imaging Techniques to Reveal Animal Anatomy. PLoS ONE 2011, 6, e17879. [Google Scholar] [CrossRef] [PubMed]
- Beuf, O.; Jaillon, F.; Saint-Jalmes, H. Small-Animal MRI: Signal-to-Noise Ratio Comparison at 7 and 1.5 T with Multiple-Animal Acquisition Strategies. Magn. Reson. Mater. Phys. 2006, 19, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Díaz, E.; González-Solís, J. Geographic assignment of seabirds to their origin: Combining morphologic, genetic, and biogeochemical analyses. Ecol. Appl. 2007, 17, 1484–1498. [Google Scholar] [CrossRef] [PubMed]
- Cory’s Shearwater|Audubon Field Guide. Available online: https://www.audubon.org/field-guide/bird/corys-shearwater (accessed on 1 January 2024).
- Navarro, J.; Kaliontzopoulou, A.; González-Solís, J. Sexual Dimorphism in Bill Morphology and Feeding Ecology in Cory’s Shearwater (Calonectris diomedea). Zoology 2009, 112, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Paiva, V.H.; Guilford, T.; Meade, J.; Geraldes, P.; Ramos, J.A.; Garthe, S. Flight Dynamics of Cory’s Shearwater Foraging in a Coastal Environment. Zoology 2010, 113, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Atchoi, E.; Mitkus, M.; Kus Vitta, P.; Machado, B.; Rocha, M.; Juliano, M.; Bried, J.L.; Rodríguez, A. Ontogenetic Exposure to Light Influences Seabird Vulnerability to Light Pollution. J. Exp. Biol. 2023, 226, jeb245126. [Google Scholar] [CrossRef] [PubMed]
- González-Varo, J.P.; Rumeu, B.; Albrecht, J.; Arroyo, J.M.; Bueno, R.S.; Burgos, T.; da Silva, L.P.; Escribano-Ávila, G.; Farwig, N.; García, D.; et al. Limited Potential for Bird Migration to Disperse Plants to Cooler Latitudes. Nature 2021, 595, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Gearheart, G.; Kooyman, G.L.; Goetz, K.T.; McDonald, B.I. Migration Front of Post-Moult Emperor Penguins. Polar Biol. 2013, 37, 435–439. [Google Scholar] [CrossRef]
- Bradarić, M.; Bouten, W.; Fijn, R.C.; Krijgsveld, K.L.; Shamoun-Baranes, J. Winds at Departure Shape Seasonal Patterns of Nocturnal Bird Migration over the North Sea. J. Avian Biol. 2020, 51, e02562. [Google Scholar] [CrossRef]
- Manola, I.; Bradarić, M.; Groenland, R.; Fijn, R.; Bouten, W.; Shamoun-Baranes, J. Associations of Synoptic Weather Conditions with Nocturnal Bird Migration Over the North Sea. Front. Ecol. Evol. 2020, 8, 542438. [Google Scholar] [CrossRef]
- Clairbaux, M.; Fort, J.; Mathewson, P.; Porter, W.; Strøm, H.; Grémillet, D. Climate Change Could Overturn Bird Migration: Transarctic Flights and High-Latitude Residency in a Sea Ice Free Arctic. Sci. Rep. 2019, 9, 17767. [Google Scholar] [CrossRef] [PubMed]
- Piersma, T.; Gutiérrez, J.S. Bird Migration: Flying High to Avoid Overheating? Curr. Biol. 2021, 31, R955–R957. [Google Scholar] [CrossRef] [PubMed]
- Galluzzo, F.G.; Cumbo, V.; Cammilleri, G.; Calabrese, V.; Pulvirenti, A.; Cicero, N.; Pantano, L.; Mascetti, A.; Lo Cascio, G.; Bacchi, E.; et al. Fatty Acids Composition of Stomach Oil of Scopoli’s Shearwater (Calonectris diomedea) from Linosa’s Colony. Animals 2022, 12, 1069. [Google Scholar] [CrossRef] [PubMed]
- Amerson, A.B. Tick Distribution in the Central Pacific as Influenced by Sea Bird Movement. J. Med. Entomol. 1968, 5, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Díaz, E.; González-Solís, J.; Peinado, M.A.; Page, R.D.M. Phylogeography of the Calonectris Shearwaters Using Molecular and Morphometric Data. Mol. Phylogenet. Evol. 2006, 41, 322–332. [Google Scholar] [CrossRef]
- Ramírez, A.S.; Poveda, J.B.; Dijkman, R.; Poveda, C.; Suárez-Pérez, A.; Rosales, R.S.; Feberwee, A.; Szostak, M.P.; Ressel, L.; Viver, T.; et al. Mycoplasma Bradburyae Sp. Nov. Isolated from the Trachea of Sea Birds. Syst. Appl. Microbiol. 2023, 46, 126472. [Google Scholar] [CrossRef] [PubMed]
- Chrastansky, A.; Callies, U.; Fleet, D.M. Estimation of the Impact of Prevailing Weather Conditions on the Occurrence of Oil-Contaminated Dead Birds on the German North Sea Coast. Environ. Pollut. 2009, 157, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.R.; Thompson, A. PCBs and Organochlorine Pesticides in Shag (Phalacrocorax dristotelis) Eggs from the Central Irish Sea: A Preliminary Study. Mar. Pollut. Bull. 1996, 32, 890–892. [Google Scholar] [CrossRef]
- Mironov, O.G. Hydrocarbon Pollution of the Sea and Its Influence on Marine Organisms. Helgol. Mar. Res. 1968, 17, 335–339. [Google Scholar] [CrossRef]
- Camphuysen, C.J.; Heubeck, M. Marine Oil Pollution and Beached Bird Surveys: The Development of a Sensitive Monitoring Instrument. Environ. Pollut. 2001, 112, 443–461. [Google Scholar] [CrossRef] [PubMed]
- Dunnet, G.M. Seabirds and North Sea Oil. Philos. Trans. R Soc. Lond. B 1987, 316, 513–524. [Google Scholar]
- Stienen, E.W.M.; Courtens, W.; Van de walle, M.; Vanermen, N.; Verstraete, H. Long-Term Monitoring Study of Beached Seabirds Shows That Chronic Oil Pollution in the Southern North Sea Has Almost Halted. Mar. Pollut. Bull. 2017, 115, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Stańczyk, E.K.; Velasco Gallego, M.L.; Nowak, M.; Hatt, J.M.; Kircher, P.R.; Carrera, I. 3.0 Tesla Magnetic Resonance Imaging Anatomy of the Central Nervous System, Eye, and Inner Ear in Birds of Prey. Vet. Radiol. Ultrasound 2018, 59, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Vilaplana Grosso, F. Orthopedic Diagnostic Imaging in Exotic Pets. Vet. Clin. Exot. Anim. Pract. 2019, 22, 149–173. [Google Scholar] [CrossRef] [PubMed]
- Fleming, G.J.; Lester, N.V.; Stevenson, R.; Silver, X.S. High field strength (4.7T) magnetic resonance imaging of hydrocephalus in an African Grey parrot (Psittacus erithacus). Vet. Radiol. Ultrasound 2003, 44, 542–545. [Google Scholar] [CrossRef] [PubMed]
- de Francisco, O.N.; Feeney, D.; Armién, A.G.; Wuenschmann, A.; Redig, P.T. Correlation of Brain Magnetic Resonance Imaging of Spontaneously Lead Poisoned Bald Eagles (Haliaeetus leucocephalus) with Histological Lesions: A Pilot Study. Res. Vet. Sci. 2016, 105, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Delk, K.W.; Mejia-Fava, J.; Jiménez, D.A.; Kent, M.; Myrna, K.; Mayer, J.; Divers, S. Diagnostic Imaging of Peripheral Vestibular Disease in a Chinese Goose (Anser cygnoides). J. Avian Med. Surg. 2014, 28, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Wernick, M.B.; Dennler, M.; Beckmann, K.; Schybli, M.; Albini, S.; Hoop, R.K.; Steffen, F.; Kircher, P.; Hatt, J.M. Peripheral Nerve Sheath Tumor in a Subadult Golden Eagle (Aquila chrysaetos). J. Avian Med. Surg. 2014, 28, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Stauber, E.; Holmes, S.; DeGhetto, D.L.; Finch, N. Magnetic Resonance Imaging Is Superior to Radiography in Evaluating Spinal Cord Trauma in Three Bald Eagles (Haliaeetus leucocephalus). J. Avian Med. Surg. 2007, 21, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Behroozi, M.; Helluy, X.; Ströckens, F.; Gao, M.; Pusch, R.; Tabrik, S.; Tegenthoff, M.; Otto, T.; Axmacher, N.; Kumsta, R.; et al. Event-Related Functional MRI of Awake Behaving Pigeons at 7T. Nat. Commun. 2020, 11, 4715. [Google Scholar] [CrossRef] [PubMed]
- Orije, J.E.M.J.; Van der Linden, A. A Brain for All Seasons: An in Vivo MRI Perspective on Songbirds. J. Exp. Zool. A Ecol. Integr. Physiol. 2022, 337, 967–984. [Google Scholar] [CrossRef] [PubMed]
- Romagnano, A.; Shiroma, J.T.; Heard, D.J.; Johnson, R.D.; Schiering, M.R.; Mladinich, C.R.J. Magnetic resonance imaging of the brain and coelomic cavity of the domestic pigeon (Columba livia domestica). Vet. Radiol. Ultrasound 1996, 37, 431–440. [Google Scholar] [CrossRef]
- Jirak, D.; Janacek, J.; Kear, B.P. A Combined MR and CT Study for Precise Quantitative Analysis of the Avian Brain OPEN. Sci. Rep. 2015, 5, 16002. [Google Scholar] [CrossRef] [PubMed]
- Faillace, A.C.L.; Vieira, K.R.A.; Santana, M.I.S. Computed tomographic and gross anatomy of the head of the blue-fronted Amazon Parrot (Amazona aestiva). Anat. Histol. Embryol. 2021, 50, 192–205. [Google Scholar] [CrossRef]
- Baumel, J.J.; Anthony, S.; King, J.E.; James, E. Handbook of Avian Anatomy: Nomina Anatomica Avium, 2nd ed.; Nuttall Ortinithological Club: Cambridge, MA, USA, 1993; pp. 318–467. [Google Scholar]
- Hadden, P.W.; Ober, W.C.; Gerneke, D.A.; Thomas, D.; Scadeng, M.; McGhee, C.N.J.; Zhang, J. Micro-CT guided illustration of the head anatomy of penguins (Aves: Sphenisciformes: Spheniscidae). J. Morphol. 2022, 283, 827–851. [Google Scholar] [CrossRef] [PubMed]
- Koenig, H.E.; Korbel, R.; Liebich, H.G.; Klupiec, C. Avian Anatomy: Textbook and Colour Atlas, 2nd ed.; 5m Books Ltd.: Sheffield, UK, 2016. [Google Scholar]
- Banzato, T.; Hellebuyck, T.; Van Caelenberg, A.; Saunders, J.H.; Zotti, A. A review of diagnostic imaging of snakes and lizards. Vet. Rec. 2013, 173, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Fumero-Hernández, M.; Encinoso, M.; Melian, A.; Nuez, H.A.; Salman, D.; Jaber, J.R. Cross Sectional Anatomy and Magnetic Resonance Imaging of the Juvenile Atlantic Puffin Head (Aves, Alcidae, Fratercula Arctica). Animals 2023, 13, 3434. [Google Scholar] [CrossRef] [PubMed]
- Medina, L.; Abellán, A.; Desfilis, E. Evolving Views on the Pallium. Brain Behav. Evol. 2021, 96, 181–199. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Sarel, A.; Derdikman, D.; Ulanovsky, N.; Gutfreund, Y. Spatial Coding in the Hippocampus and Hyperpallium of Flying Owls. Proc. Natl. Acad. Sci. USA 2023, 120, e2212418120. [Google Scholar] [CrossRef] [PubMed]
- Evans, H.E. Avian anatomy. In Handbook of Bird Biology, 3rd ed.; Lovette, I.J., Fitzpatrick, J.W., Eds.; Wiley & Sons: Chester, UK, 2016; p. 219. [Google Scholar]
- Portman, A.; Werner, S. The central nervous system. In Biology and Comparative Physiology of Birds; Marshall, A.J., Ed.; Academic Press: New York, NY, USA, 1961. [Google Scholar]
- González Rodríguez, E.; Encinoso Quintana, M.; Morales Bordon, D.; Garcés, J.G.; Artiles Nuez, H.; Jaber, J.R. Anatomical Description of Rhinoceros Iguana (Cyclura cornuta cornuta) Head by Computed Tomography, Magnetic Resonance Imaging and Gross-Sections. Animals 2023, 13, 955. [Google Scholar] [CrossRef] [PubMed]
- Arencibia, A.; Hidalgo, M.R.; Vázquez, J.M.; Contreras, S.; Ramírez, G.; Orós, J. Sectional Anatomic and Magnetic Resonance Imaging Features of the Head of Juvenile Loggerhead Sea Turtles (Caretta caretta). Am. J. Vet. Res. 2012, 73, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- Díaz Martínez, E.; Arencibia Espinosa, A.; Soler Laguía, M.; Kilroy, D.; Martínez Gomariz, F.; Casas García, D.L.; Sánchez Collado, C.; Gil Cano, F.; Jaber, J.R.; Ramírez Zarzosa, G. An Anatomical Study Using Computed Tomography, Magnetic Resonance Imaging, and Rhinoscopy of the Nasal Cavity of Domestic Cat (Felis silvestris catus L.) and Big Cats: Lion (Panthera leo leo L.), Leopard (Panthera pardus kotiya L.), and Cheetah (Acinonyx jubatus jubatus S.). Animals 2024, 14, 1172. [Google Scholar] [CrossRef] [PubMed]
- Morales-Bordon, D.; Encinoso, M.; Arencibia, A.; Jaber, J.R. Cranial Investigations of Crested Porcupine (Hystrix cristata) by Anatomical Cross-Sections and Magnetic Resonance Imaging. Animals 2023, 13, 2551. [Google Scholar] [CrossRef] [PubMed]
- Raduan Jaber, J.; Encinoso, M.; Morales, D.; Artiles, A.; Santana, M.; Blanco, D.; Arencibia, A. Anatomic Study of the Normal Bengal Tiger (Panthera tigris tigris) Brain and Associated Structures Using Low Field Magnetic Resonance Imaging. Eur. J. Anat. 2016, 20, 195–203. [Google Scholar]
- Arencibia, A.; Matos, J.; Encinoso, M.; Gil, F.; Artiles, A.; Martínez-Gomariz, F.; Vázquez, J.M. Computed Tomography and Magnetic Resonance Imaging Study of a Normal Tarsal Joint in a Bengal Tiger (Panthera tigris). BMC Vet. Res. 2019, 15, 126. [Google Scholar] [CrossRef] [PubMed]
- Capello, V. Diagnostic Imaging of Dental Disease in Pet Rabbits and Rodents. Vet. Clin. Exot. Anim. Pract. 2016, 19, 757–782. [Google Scholar] [CrossRef] [PubMed]
- Morales Espino, A.; Déniz, S.; Fumero-Hernández, M.; Encinoso, M.; Calabuig, P.; Conde-Felipe, M.; Jaber, J.R. A Cadaveric Study Using Anatomical Cross-Section and Computed Tomography for the Coelomic Cavity in Juvenile Cory’s Shearwater (Aves, Procellariidae, Calonectris borealis). Animals 2024, 14, 858. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales-Espino, A.; Déniz, S.; Paz-Oliva, P.; Roldán-Medina, N.; Encinoso, M.; Suárez-Cabrera, F.; Jaber, J.R. Cory’s Shearwater (Calonectris borealis): Exploring Normal Head Anatomy through Cross-Sectional Anatomy, Computed Tomography and Magnetic Resonance Imaging. Animals 2024, 14, 1962. https://doi.org/10.3390/ani14131962
Morales-Espino A, Déniz S, Paz-Oliva P, Roldán-Medina N, Encinoso M, Suárez-Cabrera F, Jaber JR. Cory’s Shearwater (Calonectris borealis): Exploring Normal Head Anatomy through Cross-Sectional Anatomy, Computed Tomography and Magnetic Resonance Imaging. Animals. 2024; 14(13):1962. https://doi.org/10.3390/ani14131962
Chicago/Turabian StyleMorales-Espino, Alejandro, Soraya Déniz, Pablo Paz-Oliva, Natalia Roldán-Medina, Mario Encinoso, Francisco Suárez-Cabrera, and Jose Raduan Jaber. 2024. "Cory’s Shearwater (Calonectris borealis): Exploring Normal Head Anatomy through Cross-Sectional Anatomy, Computed Tomography and Magnetic Resonance Imaging" Animals 14, no. 13: 1962. https://doi.org/10.3390/ani14131962