1. Introduction
Gamma aminobutyric acid (GABA) is a non-protein amino acid necessary for inhibitory neurotransmission, one of the major physiological functions in animals. GABA has been evaluated as a dietary supplement for different fish species including fresh water, brackish water and marine water species representing herbivorous, omnivorous and carnivorous feeding habits. GABA was reported to improve growth performance, innate immunity, gut function, digestive enzyme activities, feed utilization, hypoxia tolerance and disease resistance in fish species as a feed additive [
1,
2,
3,
4]. The supplement level of GABA was also reported to range between 0.01% and 1% in diets for different fish species including 0.5–1.0% for Indian major carp, mrigal (
Cirrhinus mrigala) [
1], 0.02% (237 mg/kg) for olive flounder (
Paralichthys olivaceus) [
2,
4] and 0.2–0.5% for Nile tilapia (
Oreochromis niloticus) [
3].
Sodium butyrate (SB), the sodium salt of butyric acid, is also a functional ingredient used in aquafeed. It is used as a single feed additive or as a compound in mixtures of other short- or medium-chain carboxylic acid salts in aquafeeds [
5,
6]. Butyrate is also supplied as a form of butyric acid glycerides (butyrin) in aquafeeds [
7,
8]. SB is well documented to improve the growth performance and feed utilization efficiency of many aquaculture species. Interestingly, it improves the gut health and digestive enzyme activities in fish [
9,
10,
11,
12] and alleviates the negative impacts of high-fat diets on liver and gut microbiota [
13,
14] while improving the resistance of fish to heat stress [
15].
Fish meal (FM) replacement in aquafeed is a critical management practice for sustainable aquaculture which minimizes the use of marine-origin resources while improving production [
16]. In this regard, GABA and SB or other butyrate forms have been evaluated in low-FM diets for several fish species and both additives were reported to enhance the efficiency of low-FM diets. GABA prevented growth impairment and intestinal malfunctions in turbot (
Scophthalmus maximus) caused by high soybean meal diets [
17]. SB is especially reported to protect the fish gut from impairments caused by high levels of soybean meal in diets. For instance, intestinal enteropathy induced by soybean meal was alleviated in turbot by dietary SB by improving the intestinal tight junction, suppressing inflammation and altering the microbial community [
18]. Gut health was improved through enhanced anti-inflammatory activities and gut microbiota in largemouth bass (
Micropterus salmoides) fed a diet containing a high level of soybean meal [
19]. The feed conversion and distal intestine morphology of the yellow drum (
Nibea albiflora) was also enhanced by SB after feeding a low-FM diet containing soybean meal [
20].
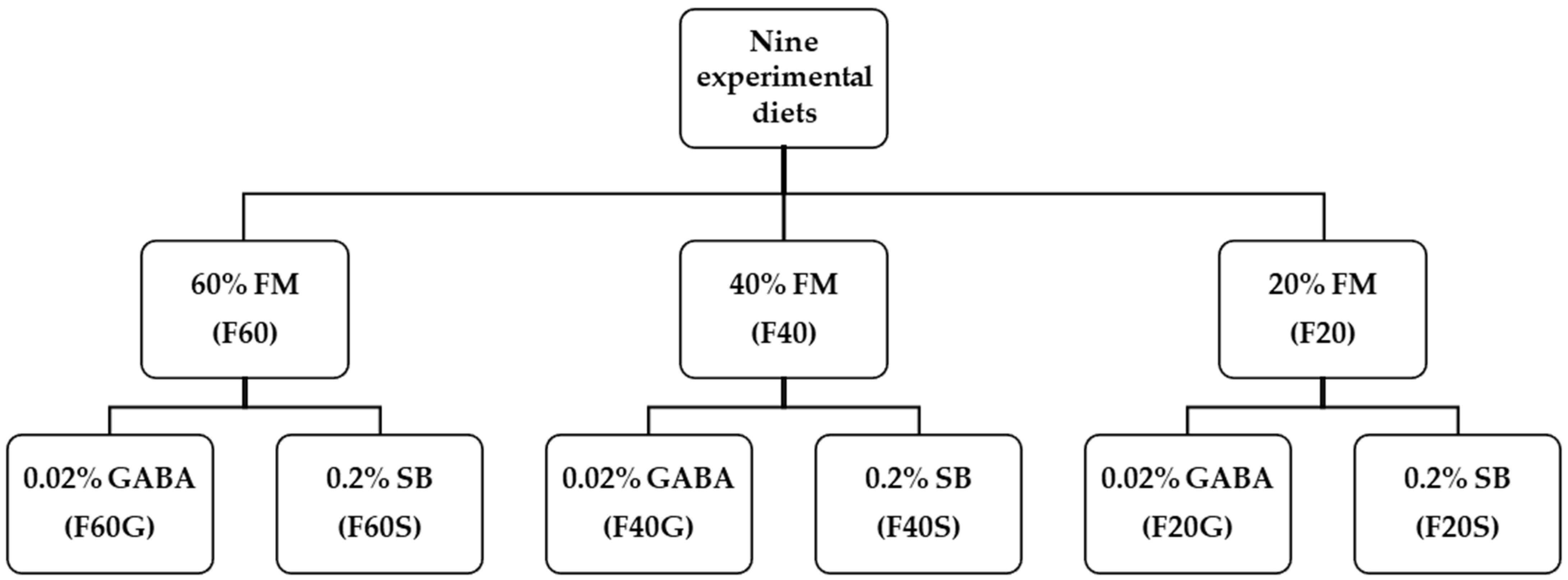
Red seabream is an important fish species cultured in South Korea. The effects of GABA, SB or other butyrate forms have not been evaluated in red seabream diets to the best of our knowledge. However, SB or other functional ingredients containing butyrate were evaluated in fish species belonging to the seabream family (Sparidae). Growth performance and nutrient digestion were increased in gilthead seabream (
Sparus aurata) fed butyrate [
21]. Two other studies revealed that SB improves the metabolic steady status of gilthead seabream fed diets containing low levels of FM and fish oil [
22,
23]. Microencapsulated SB resulted in enhanced body growth, intestinal development and antioxidative activity of juvenile black seabream (
Acanthopagrus schlegelii) when incorporated at 0.2% in diets [
24]. A mixture of dietary mono-, di- and tributyrin also improved the digestive enzyme activities of black seabream fed diets containing high levels of soybean meal [
7]. Zarei et al. [
25] evaluated increasing the levels of dietary butyric acid glycerides in yellowfin seabream diets (
Acanthopagrus latus) and reported positive influences on immunity, growth and liver antioxidant activities. The replacement of FM in the red seabream diet has been a well-studied research area over the last few decades. Different types of alternative protein sources were evaluated as FM replacers and most of these studies revealed potential ingredients to replace FM in the red seabream diet. Soy protein concentrate (SPC) is one of the successful protein ingredients used in red seabream diets [
26], although high SPC levels in diets resulted in growth impairments. Protein sources that were developed, i.e., protein hydrolysates and functional ingredients, were successfully applied to alleviate the impairments caused by high SPC levels in diets [
27,
28,
29]. Therefore, considering FM replacement and the aforementioned positive influences of both GABA and SB, we designed the present study to evaluate the effects of GABA and SB on growth performance, feed utilization, serum lysozyme activity, plasma biochemical parameters and muscle proximate composition when graded levels of FM were replaced with SPC in diets for juvenile red seabream.
4. Discussion
The efficiency of aquafeed is mainly represented by the growth performance and feed utilization of fish. The growth performance of red seabream was significantly reduced due to dietary FM replacement in the present study. GABA and SB improved fish growth performance in each FM replacement level compared to the respective control groups. Both GABA and SB are well documented as growth promoters of fish species. GABA enhanced the intestinal morphology and the expression of growth-related genes, GH, GHR2, IGF-I and IGF-II, in Nile tilapia [
3]. It upregulated the expression of ghrelin and IGF-I genes in rohu (
Labeo rohita) [
33]. Wu et al. [
34] revealed that GABA improves the expression of feeding-related genes in grass carp (
Ctenopharyngodon idellus). Dietary SB was also reported to enhance the expression of ghrelin, GH and IGF-1 in Nile tilapia [
11]. It improves the intestine absorption capacity of grass carp by accelerating the expression of peptide transporter 1 gene [
12]. SB accelerated the activities of digestive enzymes in several fish species fed diets containing a similar FM level [
35,
36]. Therefore, these positive influences of GABA and SB as fish growth promoters could be responsible for improved growth performance at each FM replacement level.
The WG and SGR of fish fed F40G and F20G diets were statistically comparable with fish fed the respective control diets (F40 and F20). The fish fed the F40G diet fully restored the performances of the diets containing 60% FM. Li et al. [
17] observed significantly higher growth performance in turbot fed a high soybean meal with GABA. They also observed that GABA ameliorates the adverse effects of a high soybean meal diet on the turbot intestine as the main reason for restored growth performance, in addition to increased FI. However, the effects of GABA were not well studied in carnivorous fish fed low-FM diets. We used SPC as the FM replacement in the red seabream diet in the present study. It is well documented that high levels of dietary SPC retarded the growth performance of red seabream [
37,
38]. FI was not significantly increased in fish groups fed diets containing GABA compared to the respective control groups, indicating that growth was not improved due to FI in the present study. SPC contains a lesser amount of antinutritional factors due to its processing technology compared to soybean meal. Therefore, the growth inhibitory effect of SPC might be lower, resulting in sufficient growth in fish fed on diets containing 40% and 20% FM without GABA or SB. On the other hand, several studies reported an optimal level of GABA in diets for better performance in fish [
33,
39]. In the current study, the inclusion level of GABA was decided according to previously reported optimum levels for other carnivorous fish species [
2]. The optimum level can be different from one fish species to another even in fish species classified in the same family. Therefore, we suggest that an optimum level of GABA should be found for red seabream diets containing low-FM levels. Further studies should also be conducted to reveal the effects of GABA on the growth performance of fish fed low-FM diets.
The growth performance of fish fed low-FM diets containing SB exhibited significantly higher results than the respective control groups. The F40S diet fully replicated the performance of the fish fed diets containing 60% FM. Several studies reported that SB improves the growth performance and feed utilization of fish fed low-FM diets. Liu et al. [
40] reported that the digestive enzyme activities, intestinal morphology, disease resistance and gut microbiota of rainbow trout was improved after feeding a low-FM diet containing SB. Liu et al. [
18] observed that SB improves the intestinal tight junction, microbial composition and anti-inflammatory activity while alleviating soybean-induced enteropathy in turbot fed high-soybean meal diets. In largemouth bass, dietary SB improved the gut health through reduced inflammation and improved gut morphology when supplemented in a high soybean meal diet [
41]. SB also improved the expression of tight junction protein and the intestinal structure of yellow drum (
Nibea albiflora) fed high soybean meal diets [
20]. Therefore, these types of positive influences might be considered as reasons for the significantly improved growth performance and feed utilization in fish fed diets containing SB compared to the respective control diets. In particular, restored feed utilization might be attributed to its effects on gut morphology, digestive enzyme activities and intestine microbiota. Future studies should be conducted to evaluate the effects of SB on the gut functions of red seabream.
Serum lysozyme activity was evaluated in all treatment groups as an indicator of innate immune functions. Serum lysozyme activity was significantly higher in the F60S group compared to the F40 and F20 groups. All the groups exhibited significantly higher results than the F20 group except for F40. Fish fed GABA- and SB-containing diets resulted in higher activity compared to the respective control diets containing similar FM levels, indicating the positive influence of both additives on innate immunity. The F20G and F20S groups exhibited significantly higher results compared to the F20 group, highlighting the restorative role of GABA and SB on retarded immunity due to FM replacement. SOD activity was slightly improved by dietary GABA at 40% and 20% FM-replacement levels although the results were not significantly different. Temu et al. [
39] observed significant improvement in SOD level in Nile tilapia when they were fed diets containing GABA, although the SOD activity was not significantly enhanced due to GABA in the present study. Li et al. [
17] observed improved antioxidant capacity in turbot fed a low-FM diet containing GABA. They suggested that GABA alleviated oxidative stress caused by high soy protein through direct involvement during lipid peroxidation, as observed by Deng et al. [
42]. SOD activity in serum and lysozyme activity in head kidney and spleen of Jian carp (
Cyprinus carpio var. Jian) were significantly improved by dietary GABA [
19]. They assumed that GABA stimulates the immune system of fish as a reason for the results. In line with present results, Farris et al. [
2] reported that dietary GABA significantly improved serum lysozyme activity of olive flounder while SOD activity was not significantly affected. They assumed the GABA-associated macrophage activation in the immune system, according to Kim et al. [
43], as the reason for improved lysozyme activity. In phagocytes, GABA participates in the GABAergic signaling process that leads to accelerated phagocytic activity [
44]. Lysozyme is one of the main enzymes utilized in phagocytic activity. Therefore, in our study, lysozyme activity in red seabream might be improved by dietary GABA, since these mechanisms have been observed to enhance activities compared to control groups. In the case of SB, the lysozyme activity of grey mullet (
Liza ramada) was significantly improved by dietary SB supplementation [
36]. They suggested that role of SB in immune stimulation and protection against lipid peroxidation to be the reason. Dawood et al. [
15] observed that the lysozyme activity of Nile tilapia was improved by dietary SB even after being subjected to heat stress. They assumed the improved activity of the intestinal microbiota was the reason for this observation. SB also exhibits the ability to reduce inflammatory responses in several fish species [
10]. Consequently, the immune responses of red seabream might be improved by SB in the present study.
Plasma biochemical parameters are important for understanding the health status and physiological condition of fish. Cholesterol and TP concentrates of red seabream plasma were significantly influenced by dietary treatments. Both parameters exhibited significant interaction with FM level or SPC level in diets regardless of the GABA or SB. In a previous study, we observed that different protein sources including SPC, alone or mixed with other protein sources, had no significant influence on plasma cholesterol or TP levels when each diet contained similar FM levels [
45]. A similar trend was observed by Biswas et al. [
46]. In contrast, neither parameter was significantly affected when a graded level of FM was replaced with fermented rapeseed meal in the juvenile red seabream diet [
47]. Kader et al. [
48] also observed no significant influence on the blood cholesterol level of red seabream fed diets with different levels of FM replaced with dehulled soybean meal. However, the TP level was significantly increased with an increase in dehulled soybean meal in diets. Biswas et al. [
49] reported no significant changes in plasma cholesterol level of red seabream fed decreasing levels of SPC. The biochemical parameters of red seabream are dependent on different factors including environmental conditions and immune status, in addition to feed [
50,
51]. Therefore, these discrepancies with the results of the previous studies might be attributed to such factors. The nutrient content of diets, mainly the amino acid and fatty acid levels, was changed with the level of feed ingredients [
45]. High dietary SPC levels affected the somatic indexes, gut enzyme activities and lipid accumulation in red seabream [
37]. These changes can influence the nutrient absorption and metabolism of fish [
52]. Accordingly, different combinations of FM and SPC in diets might have significant effects on the plasma cholesterol and TP levels in the red seabream used in the present study.
The CF, HSI and VSI were not significantly different among the dietary groups. However, the interaction of FM and SPC levels with SB exhibited a significant impact on HSI. SB showed a significant influence on VSI according to two-way ANOVA. HSI and VSI indicate the weight of liver and viscera compared to body weight. In accordance with aforementioned evidence from previous studies, SB improves intestinal structure, nutrient absorption, microbial community and health status while suppressing intestinal inflammation [
18,
41]. Accordingly, we assumed that HSI and VSI were improved due to those influences which might enhance the growth of fish fed SB-containing diets compared to each control group with different FM replacement levels. Muscle proximate compositions were also not significantly changed by the different FM or SPC levels, GABA and SB supplementation in diets. A number of previous studies have observed similar results when fish were fed diets containing similar protein and lipid levels [
6,
22]. The dietary composition of fatty acid, amino acid and other compounds, i.e., vitamins, was documented to reflect in the muscle, whole-body or liver samples [
45,
53]. Therefore, further studies should be conducted to evaluate the effects of nutrient levels in diets containing SB or GABA on the muscle, liver and whole-body composition of red seabream.
A high level of FM replacement resulted in lower performance according to most analyzed or calculated parameters in the present study. We evaluated F20 diets containing 20% FM and approximately 40% SPC as the highest FM replacement level. However, the growth performance of the control diet was not replicated even after incorporating GABA or SB. Several studies reported that some measure of FM could be replaced in red seabream diets with SPC. Hossain et al. [
28] reduced the FM level in same-sized red seabream to 15% with approximately 40% SPC, lysine, methionine and inosine monophosphate while restoring growth performance, immune responses and nutrient digestibility of diet. Biswas et al. [
46] (2017) observed restored growth of juvenile red seabream (3.9 g) when FM level was reduced to 30% with 30% SPC, 10% corn gluten and taurine compared to a control diet containing 70% FM. Biswas et al. [
26] reduced the FM level to 20% with 40% SPC, 10% corn gluten and taurine replicating the performance of red seabream (23 g) fed a control diet containing 67% FM. These previous findings are slightly inconsistent with the results of the present study indicating influences from other factors by which the growth performance of red seabream was either accelerated or decreased. Experimental feed formulated by Hossain et al. [
28] contained lysine and methionine in addition to inosine monophosphate. The studies conducted by Biswas et al. [
26,
46] formulated diets to contain approximately 13% crude ash, lecithin and 1% taurine without lysine and methionine. Water temperatures were also different among these studies. Therefore, the discrepancy can be attributed to these differences, including feed formulations, environmental conditions and fish growth stages. We suggest that the level of functional feed additives such as lysine, methionine, taurine, mineral and vitamin levels should be increased in low-FM diets in our formulation to enhance the effectiveness of F20 diets. Moreover, the trial period might also be one of the main reasons for the discrepancies. For instance, the growth performance of red seabream was not restored after a 15-week feeding trial when FM level was reduced from 40% to 25% with SPC [
6]. Future studies should be conducted to elucidate these matters and an appropriate FM level in between 40% and 20% should be found in diets containing SPC with GABA or SB.


 ,
, 


{kind=link}
{kind=link}
{kind=link}