

Investigation through Animal–Computer Interaction: A Proof-of-Concept Study for the Behavioural Experimentation of Colour Vision in Zoo-Housed Primates

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
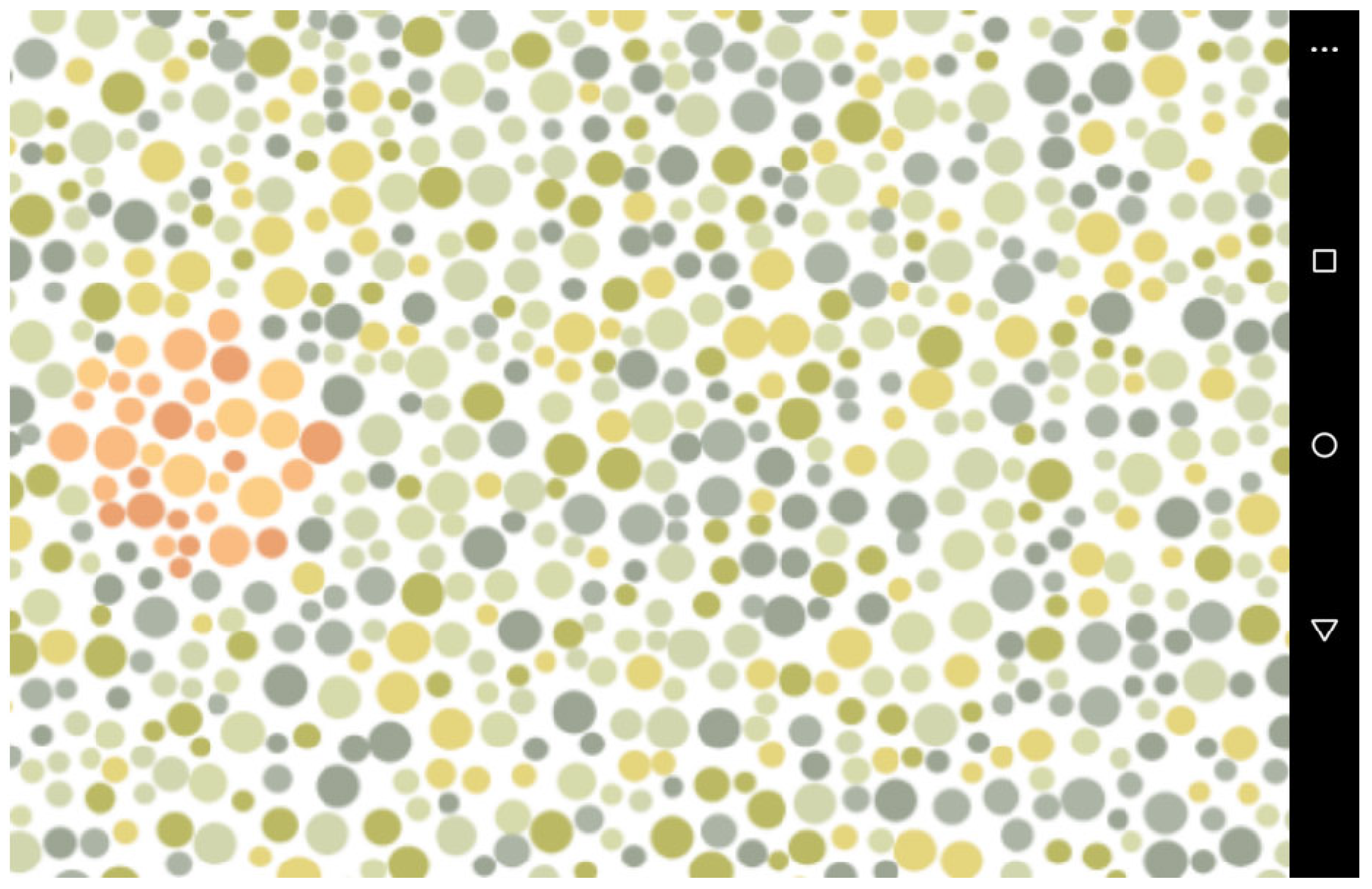
2.2. Stimuli
2.3. Apparatus
2.4. Procedure
3. Results
3.1. Stimuli
3.2. Marmosets and Titis
3.3. Spider Monkeys
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barongi, R.; Fisken, F.A.; Parker, M.; Gusset, M. Committing to Conservation: The World Zoo and Aquarium Conservation Strategy; WAZA: Gland, Switzerland, 2015. [Google Scholar]
- Patrick, P.G.; Matthews, C.E.; Ayers, D.F.; Tunnicliffe, S.D. Conservation and Education: Prominent Themes in Zoo Mission Statements. J. Environ. Educ. 2007, 38, 53–60. [Google Scholar] [CrossRef]
- Tribe, A.; Booth, R. Assessing the Role of Zoos in Wildlife Conservation. Hum. Dimens. Wildl. 2003, 8, 65–74. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Martin, A.L. Animal Training, Environmental Enrichment, and Animal Welfare: A History of Behavior Analysis in Zoos. J. Zool. Bot. Gard. 2021, 2, 531–543. [Google Scholar] [CrossRef]
- Kleiman, D.G. Behavior Research in Zoos: Past, Present, and Future. Zoo Biol. 1992, 11, 301–312. [Google Scholar] [CrossRef]
- Hosey, G.R. Behavioural Research in Zoos: Academic Perspectives. Appl. Anim. Behav. Sci. 1997, 51, 199–207. [Google Scholar] [CrossRef]
- Jacobson, S.L.; Hopper, L.M.; Shender, M.A.; Ross, S.R.; Leahy, M.; McNernie, J. Zoo Visitors’ Perceptions of Chimpanzee Welfare Are Not Affected by the Provision of Artificial Environmental Enrichment Devices in a Naturalistic Exhibit. J. Zoo Aquar. Res. 2017, 5, 56–61. [Google Scholar] [CrossRef]
- McGrath, R.E. Species-Appropriate Computer Mediated Interaction. In Proceedings of the CHI ’09 Extended Abstracts on Human Factors in Computing Systems, Boston, MA, USA, 4–9 April 2009; ACM: New York, NY, USA, 2019; pp. 2529–2534. [Google Scholar]
- Ritvo, S.E.; Allison, R.S. Designing for the Exceptional User: Nonhuman Animal-Computer Interaction (ACI). Comput. Hum. Behav. 2017, 70, 222–233. [Google Scholar] [CrossRef]
- Wirman, H.; Zamansky, A. Toward Characterization of Playful ACI. Interactions 2016, 23, 47–51. [Google Scholar] [CrossRef]
- Mancini, C. Animal-Computer Interaction: A Manifesto. Interactions 2011, 18, 69–73. [Google Scholar] [CrossRef]
- Graf, P.M.; Wilson, R.P.; Qasem, L.; Hackländer, K.; Rosell, F. The Use of Acceleration to Code for Animal Behaviours; A Case Study in Free-Ranging Eurasian Beavers Castor Fiber. PLoS ONE 2015, 10, e0136751. [Google Scholar] [CrossRef]
- Nathan, R.; Getz, W.M.; Revilla, E.; Holyoak, M.; Kadmon, R.; Saltz, D.; Smouse, P.E. A Movement Ecology Paradigm for Unifying Organismal Movement Research. Proc. Natl. Acad. Sci. USA 2008, 105, 19052–19059. [Google Scholar] [CrossRef]
- Takemoto, A.; Miwa, M.; Koba, R.; Yamaguchi, C.; Suzuki, H.; Nakamura, K. Individual Variability in Visual Discrimination and Reversal Learning Performance in Common Marmosets. Neurosci. Res. 2015, 93, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, A.; Izumi, A.; Miwa, M.; Nakamura, K. Development of a Compact and General-Purpose Experimental Apparatus with a Touch-Sensitive Screen for Use in Evaluating Cognitive Functions in Common Marmosets. J. Neurosci. Methods 2011, 199, 82–86. [Google Scholar] [CrossRef]
- Melin, A.D.; Kline, D.W.; Hickey, C.M.; Fedigan, L.M. Food Search through the Eyes of a Monkey: A Functional Substitution Approach for Assessing the Ecology of Primate Color Vision. Vis. Res. 2013, 86, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Pessoa, D.M.A.; Cunha, J.F.; Tomaz, C.; Pessoa, V.F. Colour Discrimination in the Black-Tufted-Ear Marmoset (Callithrix penicillata): Ecological Implications. Folia Primatol. 2005, 76, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, S. Molecular Evolution of Vertebrate Visual Pigments. Prog. Retin. Eye Res. 2000, 19, 385–419. [Google Scholar] [CrossRef]
- Bowmaker, J.K. Evolution of Vertebrate Visual Pigments. Vis. Res. 2008, 48, 2022–2041. [Google Scholar] [CrossRef]
- Jacobs, G.H. Evolution of Colour Vision in Mammals. Phil. Trans. R. Soc. B 2009, 364, 2957–2967. [Google Scholar] [CrossRef] [PubMed]
- Thoen, H.H.; How, M.J.; Chiou, T.-H.; Marshall, J. A Different Form of Color Vision in Mantis Shrimp. Science 2014, 343, 411–413. [Google Scholar] [CrossRef]
- Hunt, D.M.; Dulai, K.S.; Cowing, J.A.; Julliot, C.; Mollon, J.D.; Bowmaker, J.K.; Li, W.-H.; Hewett-Emmett, D. Molecular Evolution of Trichromacy in Primates. Vis. Res. 1998, 38, 3299–3306. [Google Scholar] [CrossRef]
- Jacobs, G.H. New World Monkeys and Color. Int. J. Primatol. 2007, 28, 729–759. [Google Scholar] [CrossRef]
- De Valois, R.L.; Jacobs, G.H. Primate Color Vision: The Macaque and Squirrel Monkey Differ in Their Color Vision and in the Physiology of Their Visual Systems. Science 1968, 162, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Dulai, K.S.; Von Dornum, M.; Mollon, J.D.; Hunt, D.M. The Evolution of Trichromatic Color Vision by Opsin Gene Duplication in New World and Old World Primates. Genome Res. 1999, 9, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, G.H.; Neitz, M.; Deegan, J.F.; Neitz, J. Trichromatic Colour Vision in New World Monkeys. Nature 1996, 382, 156–158. [Google Scholar] [CrossRef] [PubMed]
- Dominy, N.J.; Lucas, P.W. Ecological Importance of Trichromatic Vision to Primates. Nature 2001, 410, 363–366. [Google Scholar] [CrossRef]
- Morgan, M.J.; Adam, A.; Mollon, J.D. Dichromats Detect Colour-Camouflaged Objects That Are Not Detected by Trichromats. Proc. R. Soc. Lond. B 1992, 248, 291–295. [Google Scholar] [CrossRef]
- Osorio, D.; Vorobyev, M. Colour Vision as an Adaptation to Frugivory in Primates. Proc. R. Soc. B. 1996, 263, 593–599. [Google Scholar] [CrossRef]
- Pessoa, D.M.A.; Maia, R.; De Albuquerque Ajuz, R.C.; De Moraes, P.Z.P.M.R.; Spyrides, M.H.C.; Pessoa, V.F. The Adaptive Value of Primate Color Vision for Predator Detection. Am. J. Primatol. 2014, 76, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Verhulst, S.; Maes, F.W. Scotopic Vision in Colour-Blinds. Vis. Res. 1998, 38, 3387–3390. [Google Scholar] [CrossRef]
- Melin, A.D.; Fedigan, L.M.; Hiramatsu, C.; Sendall, C.L.; Kawamura, S. Effects of Colour Vision Phenotype on Insect Capture by a Free-Ranging Population of White-Faced Capuchins, Cebus Capucinus. Anim. Behav. 2007, 73, 205–214. [Google Scholar] [CrossRef]
- Melin, A.D.; Chiou, K.L.; Walco, E.R.; Bergstrom, M.L.; Kawamura, S.; Fedigan, L.M. Trichromacy Increases Fruit Intake Rates of Wild Capuchins (Cebus Capucinus Imitator). Proc. Natl. Acad. Sci. USA 2017, 114, 10402–10407. [Google Scholar] [CrossRef] [PubMed]
- Maia, R.; Eliason, C.M.; Bitton, P.; Doucet, S.M.; Shawkey, M.D. Pavo: An R Package for the Analysis, Visualization and Organization of Spectral Data. Methods Ecol. Evol. 2013, 4, 906–913. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Shettleworth, S.J. Animal Cognition and Animal Behaviour. Anim. Behav. 2001, 61, 277–286. [Google Scholar] [CrossRef]
- Endler, J.A. The Color of Light in Forests and Its Implications. Ecol. Monogr. 1993, 63, 1–27. [Google Scholar] [CrossRef]
- Isbell, L.A. Snakes as Agents of Evolutionary Change in Primate Brains. J. Hum. Evol. 2006, 51, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M. Sensory Ecology, Behaviour, and Evolution; Oxford University Press: Oxford, UK, 2013; ISBN 978-0-19-960177-6. [Google Scholar]
- Melin, A.D.; Fedigan, L.M.; Hiramatsu, C.; Kawamura, S. Polymorphic Color Vision in White-Faced Capuchins (Cebus Capucinus): Is There Foraging Niche Divergence among Phenotypes? Behav. Ecol. Sociobiol. 2008, 62, 659–670. [Google Scholar] [CrossRef]
- Hiwatashi, T.; Okabe, Y.; Tsutsui, T.; Hiramatsu, C.; Melin, A.D.; Oota, H.; Schaffner, C.M.; Aureli, F.; Fedigan, L.M.; Innan, H.; et al. An Explicit Signature of Balancing Selection for Color-Vision Variation in New World Monkeys. Mol. Biol. Evol. 2010, 27, 453–464. [Google Scholar] [CrossRef]
- Duarte, M.H.L.; Goulart, V.D.L.R.; Young, R.J. Designing Laboratory Marmoset Housing: What Can We Learn from Urban Marmosets? Appl. Anim. Behav. Sci. 2012, 137, 127–136. [Google Scholar] [CrossRef]
- Goulart, V.D.L.R.; Teixeira, C.P.; Young, R.J. Analysis of Callouts Made in Relation to Wild Urban Marmosets (Callithrix penicillata) and Their Implications for Urban Species Management. Eur. J. Wildl. Res. 2010, 56, 641–649. [Google Scholar] [CrossRef]
- Teixeira, B.; Hirsch, A.; Goulart, V.D.L.R.; Passos, L.; Teixeira, C.P.; James, P.; Young, R. Good Neighbours: Distribution of Black-Tufted Marmoset (Callithrix penicillata) in an Urban Environment. Wildl. Res. 2015, 42, 579. [Google Scholar] [CrossRef]
- Carlstead, K. A Comparative Approach to the Study of Keeper–Animal Relationships in the Zoo. Zoo Biol. 2009, 28, 589–608. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Cost |
|---|---|
| Kindle Fire tablet computer, 7-inch 1024 × 600 screen, 313 g | GBP 35.00 |
| Arduino Uno microcontroller board ATmega328P | GBP 17.30 |
| DC motor-powered dispenser | GBP 5.00 |
| Bluetooth module Hm-10 | GBP 6.00 |
| L298N dual H-bridge DC motor module | GBP 5.00 |
| Rugged tablet case with screen protector | GBP 15.00 |
| Colour Category | Hex-Colour Code | CIE-Lab L | CIE-Lab A | CIE-Lab L |
|---|---|---|---|---|
| Green | d9CA594 | 66.6087 | −6.3958 | 7.7143 |
| Green | ACB4A5 | 72.3152 | −5.5814 | 6.6789 |
| Green | BBB946 | 73.3536 | −13.7646 | 56.6906 |
| Green | D1D6AF | 84.3514 | −8.2588 | 18.7266 |
| Green | D7DAAA | 85.7519 | −8.8320 | 23.3500 |
| Green | E5D57D | 84.8571 | −6.2664 | 45.3062 |
| Red | EBA170 | 72.4431 | 22.2198 | 36.4731 |
| Red | F9BB82 | 80.3436 | 15.6987 | 37.4752 |
| Red | FCCD84 | 85.0087 | 7.6967 | 42.4568 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goulart, V.D.L.R.; Young, R.J. Investigation through Animal–Computer Interaction: A Proof-of-Concept Study for the Behavioural Experimentation of Colour Vision in Zoo-Housed Primates. Animals 2024, 14, 1979. https://doi.org/10.3390/ani14131979
Goulart VDLR, Young RJ. Investigation through Animal–Computer Interaction: A Proof-of-Concept Study for the Behavioural Experimentation of Colour Vision in Zoo-Housed Primates. Animals. 2024; 14(13):1979. https://doi.org/10.3390/ani14131979
Chicago/Turabian StyleGoulart, Vinícius Donisete Lima Rodrigues, and Robert John Young. 2024. "Investigation through Animal–Computer Interaction: A Proof-of-Concept Study for the Behavioural Experimentation of Colour Vision in Zoo-Housed Primates" Animals 14, no. 13: 1979. https://doi.org/10.3390/ani14131979
APA StyleGoulart, V. D. L. R., & Young, R. J. (2024). Investigation through Animal–Computer Interaction: A Proof-of-Concept Study for the Behavioural Experimentation of Colour Vision in Zoo-Housed Primates. Animals, 14(13), 1979. https://doi.org/10.3390/ani14131979