
First Feeding of Cuttlefish Hatchlings: Pioneering Attempts in Captive Breeding
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cuttlefish Collection and Rearing
2.2. Experimental Diets and Sampling
2.3. Morphological Studies
2.4. Cuttlefish Gut Histology
2.5. Digestive Enzyme Activity
2.6. RNA Extraction, cDNA Synthesis and Real Time PCR Analysis
2.7. Statistical Analysis
3. Results
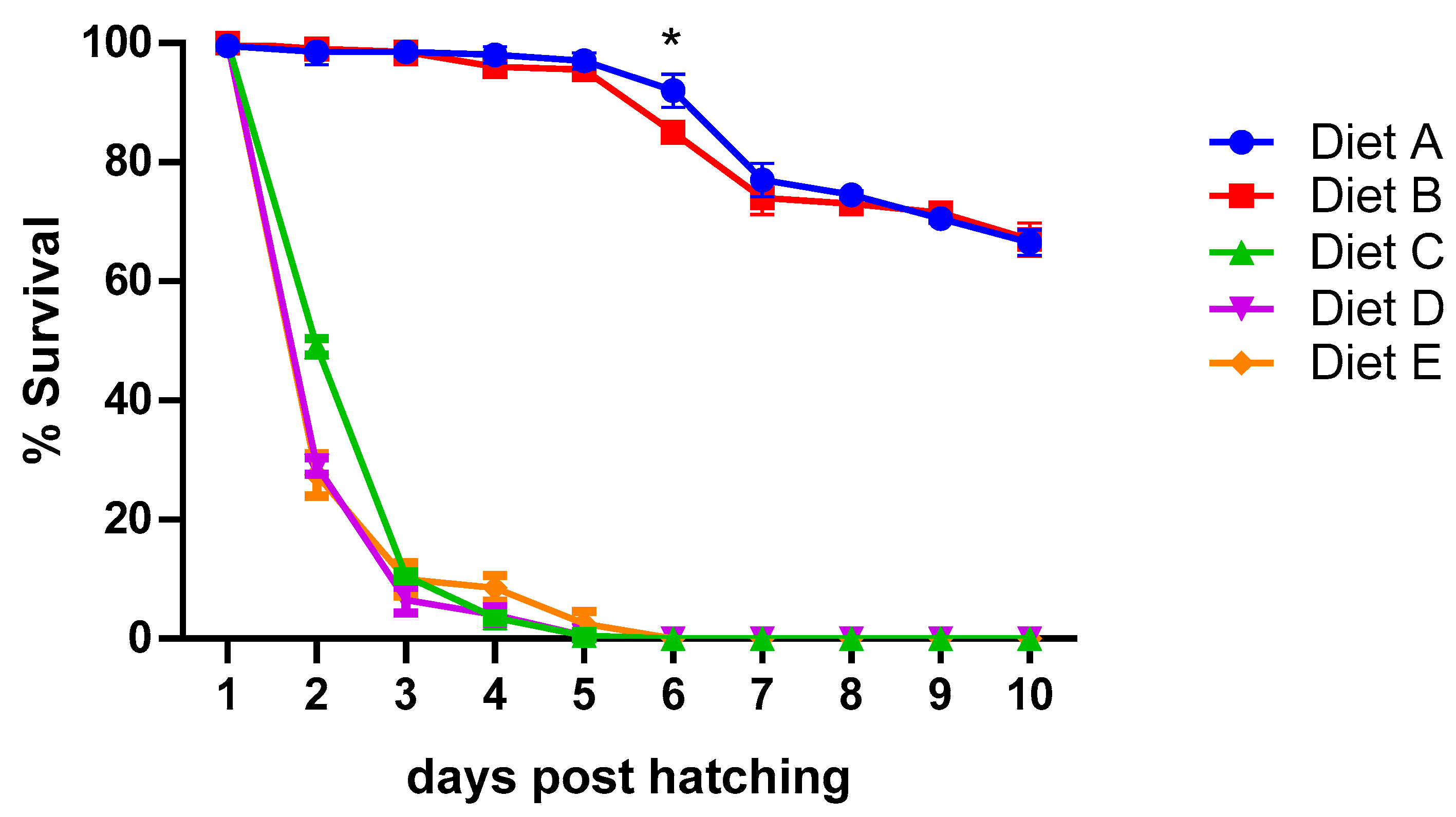
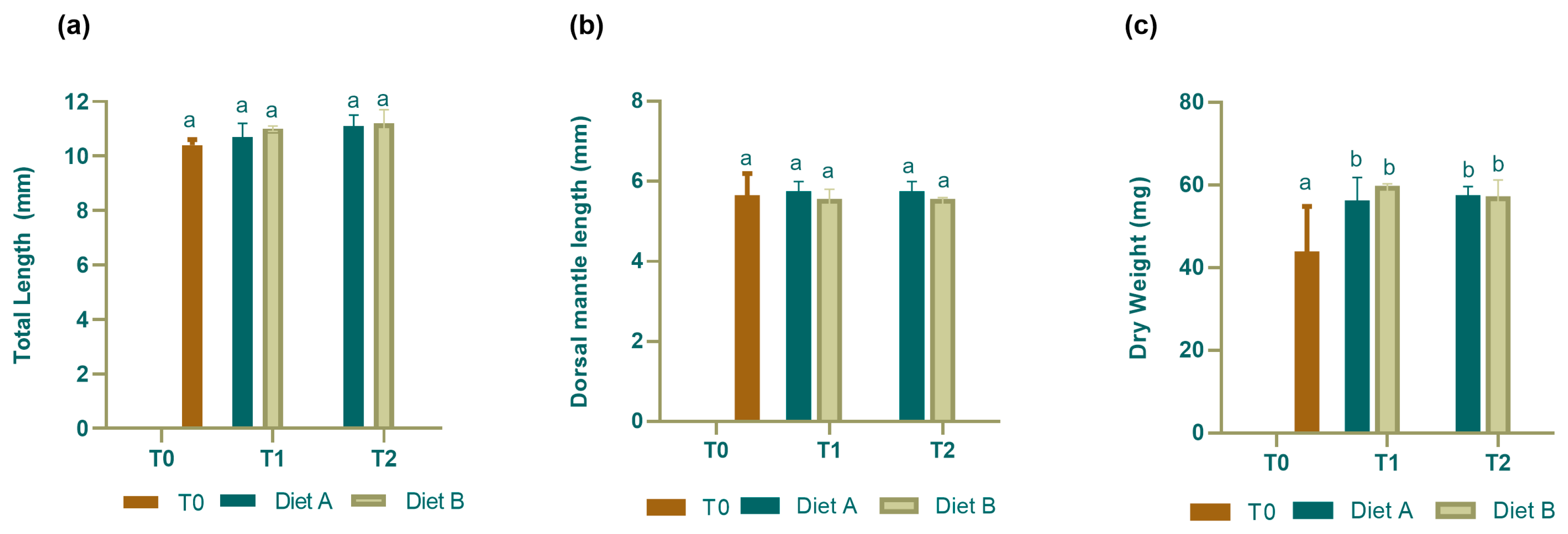
3.1. Feeding Trials
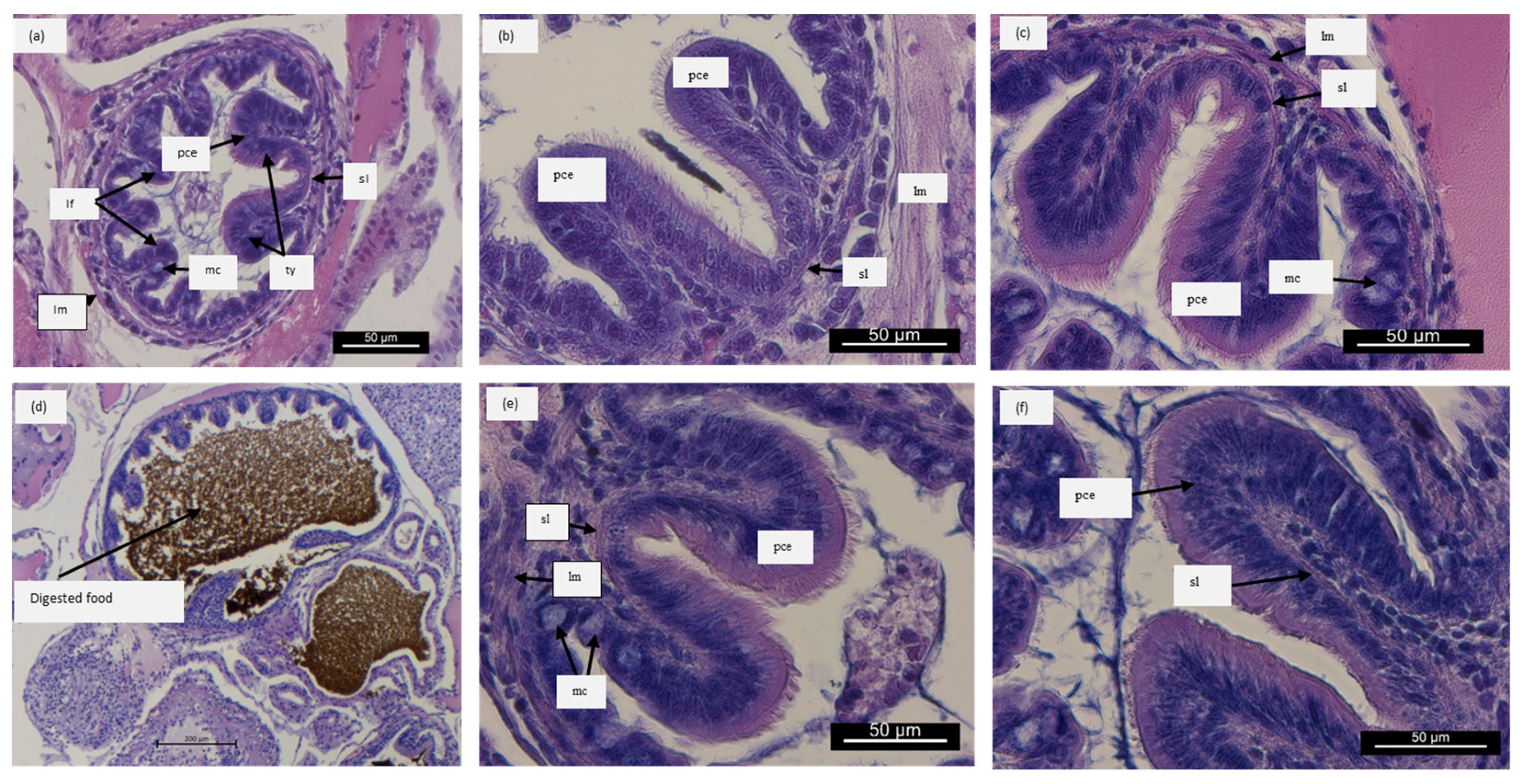
3.2. Histological Results
3.3. Digestive Enzyme Activity
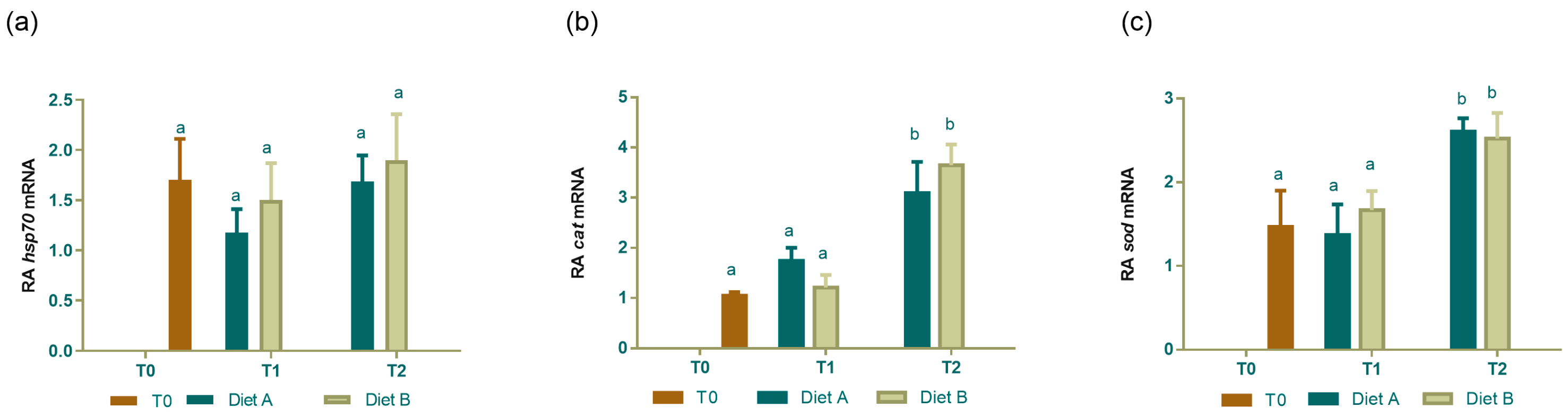
3.4. Real-Time PCR of Genes Involved in Stress Response and Antioxidant System
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barrett, C.J.; Bensbai, J.; Broadhurst, M.K.; Bustamante, P.; Clark, R.; Cooke, G.M.; Di Cosmo, A.; Drerup, C.; Escolar, O.; Fernández-Álvarez, F.A.; et al. Cuttlefish Conservation: A Global Review of Methods to Ameliorate Unwanted Fishing Mortality and Other Anthropogenic Threats to Sustainability. ICES J. Mar. Sci. 2023, 79, 2579–2596. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Veneroni, B.; Fernandes, P.G. Fishers’ Knowledge Detects Ecological Decay in the Mediterranean Sea. Ambio 2021, 50, 1159–1171. [Google Scholar] [CrossRef]
- Lega Coop Veneto-Agriteco. Fondo Europeo Pesca La Pesca Delle Seppie in Veneto: Una Proposta Di Gestione Pergarantirne La Riproduzione 2013; Lega Coop Veneto-Agriteco: Venice, Italy, 2011. [Google Scholar]
- Jereb, P.; Allcock, A.L.; Lefkaditou, E.; Piatkowski, U.; Hastie, L.C.; Pierce, G.J. Cephalopod Biology and Fisheries in Europe: II. Species Accounts; ICES: Copenhagen, Denmark, 2015; Volume 325. [Google Scholar]
- Castro, B.G.; Guerra, A. The Diet of Sepia Officinalis (Linnaeus, 1758) and Sepia Elegans (D’Orbigny, 1835) (Cephalopoda, Sepioidea) from the Ria de Vigo (NW Spain). Sci. Mar. 1990, 54, 375–388. [Google Scholar]
- Koueta, N.; Boucaud-Camou, E. Food Intake and Growth in Reared Early Juvenile Cuttlefish. J. Exp. Biol. Ecol. 1999, 240, 93–109. [Google Scholar] [CrossRef]
- Sykes, A.V.; Domingues, P.M.; Andrade, J.P. Effects of Using Live Grass Shrimp (Palaemonetes varians) as the Only Source of Food for the Culture of Cuttlefish, Sepia officinalis (Linnaeus, 1758). Aquac. Int. 2006, 14, 551–568. [Google Scholar] [CrossRef]
- Domingues, P.; Poirier, R.; Dickel, L.; Almansa, E.; Sykes, A.; Andrade, J.P. Effects of Culture Density and Live Prey on Growth and Survival of Juvenile Cuttlefish, Sepia officinalis. Aquac. Int. 2003, 11, 225–242. [Google Scholar] [CrossRef]
- Domingues, P.; Márquez, L. Effects of Culture Density and Bottom Area on Growth and Survival of the Cuttlefish Sepia officinalis (Linnaeus, 1758). Aquac. Int. 2010, 18, 361–369. [Google Scholar] [CrossRef]
- Aravind, N.P.; Sheeba, P.; Nair, K.K.C.; Achuthankutty, C.T. Life History and Population Dynamics of an Estuarine Amphipod, Eriopisa chilkensis Chilton (Gammaridae). Estuar. Coast. Shelf Sci. 2007, 74, 87–95. [Google Scholar] [CrossRef]
- Nakajima, K.; Takeuchi, I. Rearing Method for Caprella mutica (Malacostraca: Amphipoda) in an Exhibition Tank in the Port of Nagoya Public Aquarium, with Notes on Reproductive Biology. J. Crustac. Biol. 2008, 28, 171–174. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Maradonna, F.; Faheem, M.; Harikrishnan, R.; Devi, G.; Ringø, E.; Van Doan, H.; Ashouri, G.; Gioacchini, G.; Carnevali, O. Sustainable Ornamental Fish Aquaculture: The Implication of Microbial Feed Additives. Animals 2023, 13, 1583. [Google Scholar] [CrossRef]
- Vidal, E.A.G.; Shea, E.K. Cephalopod Ontogeny and Life Cycle Patterns. Front. Mar. Sci. 2023, 10, 1162735. [Google Scholar] [CrossRef]
- Darmaillacq, A.S.; Chichery, R.; Dickel, L. Food Imprinting, New Evidence from the Cuttlefish Sepia Officinalis. Biol. Lett. 2006, 2, 345–347. [Google Scholar] [CrossRef]
- Sykes, A.V.; Gonçalves, R.A.; Andrade, J.P. Early Weaning of Cuttlefish (Sepia officinalis, L.) with Frozen Grass Shrimp (Palaemonetes varians) from the First Day after Hatching. Aquac. Res. 2013, 44, 1815–1823. [Google Scholar] [CrossRef]
- Jiang, M.; Chen, Q.; Zhou, S.; Han, Q.; Peng, R.; Jiang, X. Optimum Weaning Method for Pharaoh Cuttlefish, Sepia pharaonis Ehrenberg, 1831, in Small- and Large-Scale Aquaculture. Aquac. Res. 2021, 52, 1078–1087. [Google Scholar] [CrossRef]
- Melli, V.; Riginella, E.; Nalon, M.; Mazzoldi, C. From Trap to Nursery. Mitigating the Impact of an Artisanal Fishery on Cuttlefish Offspring. PLoS ONE 2014, 9, e90542. [Google Scholar] [CrossRef] [PubMed]
- Sykes, A.V.; Pereira, D.; Rodríguez, C.; Lorenzo, A.; Andrade, J.P. Effects of Increased Tank Bottom Areas on Cuttlefish (Sepia officinalis, L.) Reproduction Performance. Aquac. Res. 2013, 44, 1017–1028. [Google Scholar] [CrossRef]
- Iglesias, J.; Villanueva, R.; Fuentes, L. (Eds.) Cephalopod Culture; Springer: Dordrecht, The Netherlands, 2014; ISBN 9789401786485. [Google Scholar]
- Capaz, J.C.; Hernández-Brooke, D.; Balvet, S.; Couto, A.T.; Alves, A.C.; Gonçalves, R.A.; Frias, P.A.; Andrade, J.P.; Sykes, A.V. Control of Zootechnology Leads to Improved Cuttlefish (Sepia officinalis, L.) Reproduction Performance up to Pre-Industrial Levels. Front. Mar. Sci. 2020, 7, 112. [Google Scholar] [CrossRef]
- Chemello, G.; Faraoni, V.; Notarstefano, V.; Maradonna, F.; Carnevali, O.; Gioacchini, G. First Evidence of Microplastics in the Yolk and Embryos of Common Cuttlefish (Sepia officinalis) from the Central Adriatic Sea: Evaluation of Embryo and Hatchling Structural Integrity and Development. Animals 2023, 13, 95. [Google Scholar] [CrossRef] [PubMed]
- Randazzo, B.; Zarantoniello, M.; Secci, G.; Faccenda, F.; Fava, F.; Marzorati, G.; Belloni, A.; Maradonna, F.; Orazi, V.; Cerri, R.; et al. Towards the Identification of a Suitable Commercial Diet for Carpione (Salmo carpio, Linnaeus 1758): A Multidisciplinary Study on Fish Performances, Animal Welfare and Quality Traits. Animals 2022, 12, 1918. [Google Scholar] [CrossRef]
- Mardones, O.; Oyarzún-Salazar, R.; Labbé, B.S.; Miguez, J.M.; Vargas-Chacoff, L.; Muñoz, J.L.P. Intestinal Variation of Serotonin, Melatonin, and Digestive Enzymes Activities along Food Passage Time through GIT in Salmo Salar Fed with Supplemented Diets with Tryptophan and Melatonin. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2022, 266, 111159. [Google Scholar] [CrossRef]
- Le Bihan, E.; Perrin, A.; Koueta, N. Effect of Different Treatments on the Quality of Cuttlefish (Sepia officinalis L.) Viscera. Food Chem. 2007, 104, 345–352. [Google Scholar] [CrossRef]
- Versaw, W.K.; Cuppett, S.L.; Winters, D.D.; Williams, L.E. An Improved Colorimetric Assay for Bacterial Lipase in Nonfat Dry Milk. J. Food Sci. 1989, 54, 1557–1558. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Using the Principle of Protein Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Forner-Piquer, I.; Mylonas, C.C.; Calduch-Giner, J.; Maradonna, F.; Gioacchini, G.; Allarà, M.; Piscitelli, F.; Di Marzo, V.; Pérez-Sánchez, J.; Carnevali, O. Endocrine Disruptors in the Diet of Male Sparus aurata: Modulation of the Endocannabinoid System at the Hepatic and Central Level by Di-Isononyl Phthalate and Bisphenol A. Environ. Int. 2018, 119, 54–65. [Google Scholar] [CrossRef]
- Forner-Piquer, I.; Santangeli, S.; Maradonna, F.; Rabbito, A.; Piscitelli, F.; Habibi, H.R.; Di, V. Disruption of the Gonadal Endocannabinoid System in Zebra Fi Sh Exposed to Diisononyl Phthalate. Environ. Pollut. 2018, 241, 1–8. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Thøgersen, R.; Bertram, H.C.; Vangsoe, M.T.; Hansen, M. Krill Protein Hydrolysate Provides High Absorption Rate for All Essential Amino Acids—A Randomized Control Cross-over Trial. Nutrients 2021, 13, 3187. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Park, S.Y.; Han, E.J.; Kim, H.S.; Kang, D.S.; Je, J.Y.; Ahn, C.B.; Ahn, G. Isolation of an Antioxidant Peptide from Krill Protein Hydrolysates as a Novel Agent with Potential Hepatoprotective Effects. J. Funct. Foods 2020, 67, 103889. [Google Scholar] [CrossRef]
- Rodríguez-Tomàs, E.; Baiges-Gaya, G.; Castañé, H.; Arenas, M.; Camps, J.; Joven, J. Trace Elements under the Spotlight: A Powerful Nutritional Tool in Cancer. J. Trace Elem. Med. Biol. 2021, 68, 126858. [Google Scholar] [CrossRef]
- Davis, D.A.; Gatlin, D.M. Dietary Mineral Requirements of Fish and Marine Crustaceans Dietary Mineral Requirements of Fish and Marine Crustaceans. Rev. Fish. Sci. 1996, 4, 75–99. [Google Scholar] [CrossRef]
- Lemos, D.; Coelho, R.; Zwart, S.; Tacon, A.G.J. Performance and Digestibility of Inorganic Phosphates in Diets for Juvenile Shrimp (Litopenaeus vannamei): Dicalcium Phosphate, Monocalcium Phosphate, and Monoammonium Phosphate. Aquac. Int. 2021, 29, 681–695. [Google Scholar] [CrossRef]
- El Idrissi, O.; Gobert, S.; Delmas, A.; Demolliens, M.; Aiello, A.; Pasqualini, V.; Ternengo, S. Effects of Trace Elements Contaminations on the Larval Development of Paracentrotus lividus Using an Innovative Experimental Approach. Aquat. Toxicol. 2022, 246, 106152. [Google Scholar] [CrossRef] [PubMed]
- Levallois, A.; Costil, K.; Caplat, C.; Basuyaux, O.; Lebel, J.M.; Guegan, C.; Serpentini, A. Comparative Effects of Trace Metal Elements Released from Dissolution of Aluminum-Based Galvanic Anodes, Aluminum Chloride, Zinc Chloride, and Their Mixture on the Development of the Pacific Oyster D-Larvae, Crassostrea gigas. Environ. Sci. Pollut. Res. 2023, 30, 101535–101545. [Google Scholar] [CrossRef] [PubMed]
- Holt, G.J. Larval Fish Nutrition; John Wiley & Sons: Hoboken, NJ, USA, 2011; ISBN 9780813817927. [Google Scholar]
- Lucas, J.S.; Southgate, P.C.; Tucker, C.S. (Eds.) Aquaculture: Farming Aquatic Animals and Plants, 3rd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013; ISBN 978-1-119-23086-1. [Google Scholar]
- Pan, X.; Zheng, Y.; Lei, K.; Tao, W.; Zhou, N. Systematic Analysis of Heat Shock Protein 70 (HSP70) Gene Family in Radish and Potential Roles in Stress Tolerance. BMC Plant Biol. 2024, 24, 2. [Google Scholar] [CrossRef] [PubMed]
- Maradonna, F.; Bavestrello, G.; Cardinali, M.; Olivotto, I.; Cerrano, C.; Giovine, M.; Carnevali, O. Role of Substrate on Larval Development of the Freshwater Teleost Pelvicachromis Pulcher. Mol. Reprod. Dev. 2003, 66, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Maradonna, F.; Carnevali, O. Vitellogenin, Zona Radiata Protein, Cathepsin D and Heat Shock Protein 70 as Biomarkers of Exposure to Xenobiotics. Biomarkers 2007, 12, 240–255. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, M.S.; Faleiro, F.; Diniz, M.; Machado, J.; Pousão-Ferreira, P.; Peck, M.A.; Pörtner, H.O.; Rosa, R. Oxidative Stress and Digestive Enzyme Activity of Flatfish Larvae in a Changing Ocean. PLoS ONE 2015, 10, e0134082. [Google Scholar] [CrossRef]
- Rønnestad, I.; Yúfera, M.; Ueberschär, B.; Ribeiro, L.; Sæle, Ø.; Boglione, C. Feeding Behaviour and Digestive Physiology in Larval Fish: Current Knowledge, and Gaps and Bottlenecks in Research. Rev. Aquac. 2013, 5, S59–S98. [Google Scholar] [CrossRef]
- Bakke, A.M.; Glover, C.; Krogdahl, Å. Feeding, Digestion and Absorption of Nutrients. In Feeding, Digestion and Absorption of Nutrients; Grosell, M., Farrell, A., Brauner, C., Eds.; Elsevier: London, UK, 2011; p. 444. [Google Scholar]
- Lallès, J.P. Intestinal Alkaline Phosphatase in the Gastrointestinal Tract of Fish: Biology, Ontogeny, and Environmental and Nutritional Modulation. Rev. Aquac. 2020, 12, 555–581. [Google Scholar] [CrossRef]
- Navarro, J.C.; Villanueva, R. Lipid and Fatty Acid Composition of Early Stages of Cephalopods: An Approach to Their Lipid Requirements. Aquaculture 2000, 183, 161–177. [Google Scholar] [CrossRef]
- Almansa, E.; Domingues, P.; Sykes, A.; Tejera, N.; Lorenzo, A.; Andrade, J.P. The Effects of Feeding with Shrimp or Fish Fry on Growth and Mantle Lipid Composition of Juvenile and Adult Cuttlefish (Sepia officinalis). Aquaculture 2006, 256, 403–413. [Google Scholar] [CrossRef]
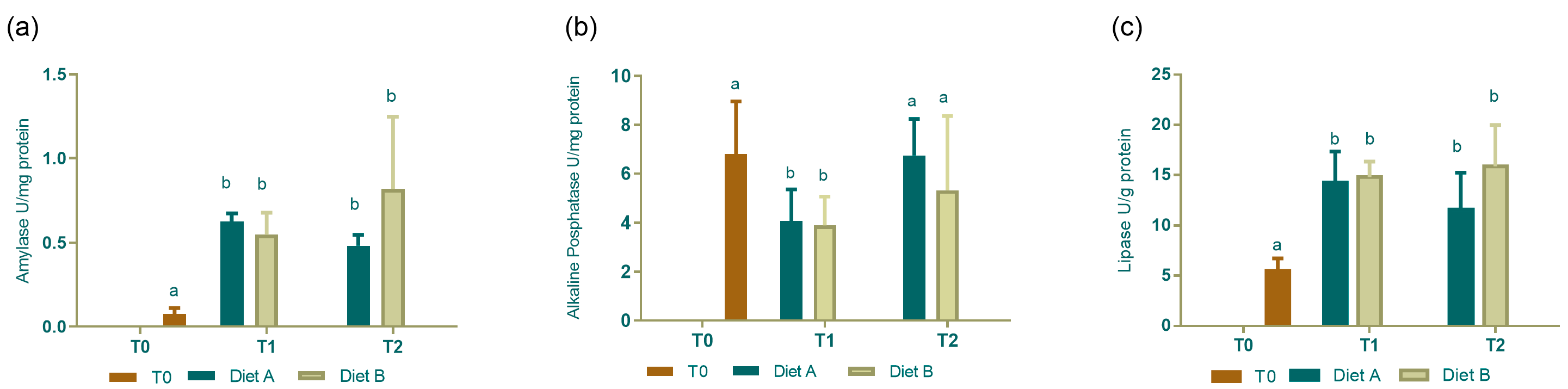

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mRNA | Primer Sequence | Annealing T |
|---|---|---|
| βtub for | 5′ GGTGCTTGTCAAGGTCCCTA 3′ | 56 |
| βtub rev | 5′ GTCAACAGCAACCAGTTTCCT 3′ | |
| 18S for | 5′ GTCGGTTTTCTCACGCACTT 3′ | 57 |
| 18S rev | 5′ CGGGAGGTGGTTAAGAGGTT 3′ | |
| cat for | 5′ TTCGTTTCTCTACCGTTGGTG 3′ | 58 |
| cat rev | 5′ AAGTCCCAGTTACCGTCTTCC 3′ | |
| Cu-Zn sod for | 5′ GAGACTTTCGTTAGGACGGATA 3′ | 56 |
| Cu-Zn sod rev | 5′ AGCCATTCCCCTTATTTCAC 3′ | |
| hsp70 for | 5′ GAGTTCAAGCGAAAGCACA 3′ | 59 |
| hsp70 rev | 5′ AGCGACCTACAAGGACAAT 3′ |
| Treatments | |||||
|---|---|---|---|---|---|
| Diet A | Diet B | ||||
| Analyzed Parameters | T0 | T1 | T2 | T1 | T2 |
| Fold height (μm) | 100.6 ± 2.0 a | 124.2 ± 7.8 bc | 122.2 ± 4.8 c | 122.6 ± 4.3 c | 140.0 ± 8.9 b |
| Fold width (μm) | 61.0 ± 1.1 a | 66.5 ± 2.9 ab | 64.0 ± 1.1 a | 62.5 ± 1.8 a | 70.9 ± 2.6 b |
| Cilia height (μm) | 6.0 ± 0.2 a | 5.9 ± 0.3 a | 6.2 ± 0.2 a | 5.7 ± 0.3 a | 5.9 ± 0.3 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maradonna, F.; Pessina, A.; Ashouri, G.; Notti, E.; Chemello, G.; Russo, G.; Gioacchini, G.; Carnevali, O. First Feeding of Cuttlefish Hatchlings: Pioneering Attempts in Captive Breeding. Animals 2024, 14, 1993. https://doi.org/10.3390/ani14131993
Maradonna F, Pessina A, Ashouri G, Notti E, Chemello G, Russo G, Gioacchini G, Carnevali O. First Feeding of Cuttlefish Hatchlings: Pioneering Attempts in Captive Breeding. Animals. 2024; 14(13):1993. https://doi.org/10.3390/ani14131993
Chicago/Turabian StyleMaradonna, Francesca, Andrea Pessina, Ghasem Ashouri, Emilio Notti, Giulia Chemello, Giulia Russo, Giorgia Gioacchini, and Oliana Carnevali. 2024. "First Feeding of Cuttlefish Hatchlings: Pioneering Attempts in Captive Breeding" Animals 14, no. 13: 1993. https://doi.org/10.3390/ani14131993