

Dietary Supplementation of Crossbred Pigs with Glycerol, Vitamin C, and Niacinamide Alters the Composition of Gut Flora and Gut Flora-Derived Metabolites
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Sample Collection
2.3. Iron and Glycerol Contents in Fecal Samples
2.4. Metagenomic Sequencing
2.5. Metabolomic Examination of Feces Using Untargeted Liquid Chromatography/Mass Spectrometry (LC/MS)
2.6. Analysis of Fecal Bile Acids Based on Targeted LC/MS
2.7. Statistical Analysis
3. Results
3.1. Concentrations of Glycerol and Ferric and Ferrous Irons in Fecal Samples
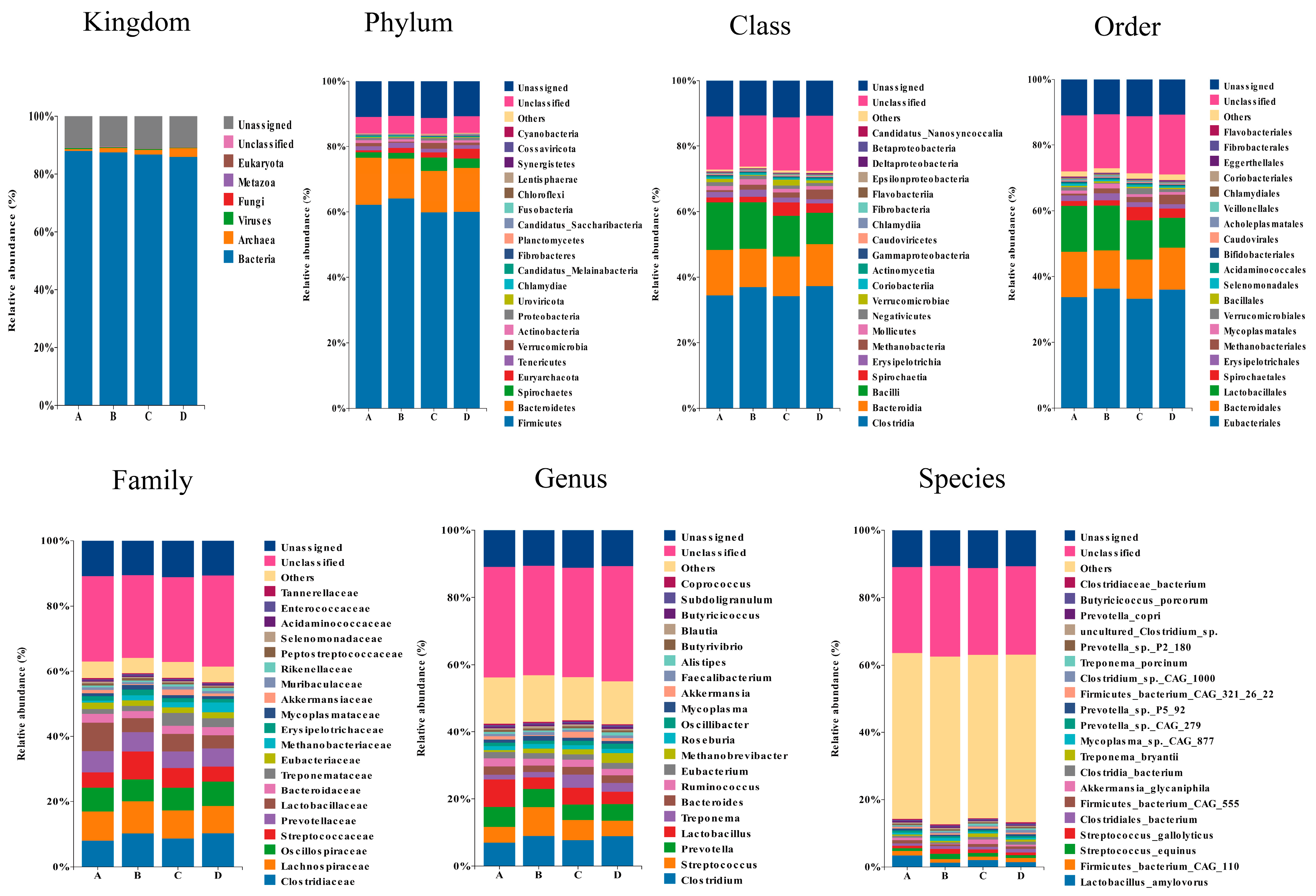
3.2. Fecal Metagenomic Profiles
3.3. Fecal Metabolomic Profiles
3.4. Concentrations of Fecal Bile Acids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Y.; Zhong, W.; Li, X.; Shen, F.; Ma, X.; Yang, Q.; Hong, S.; Sun, Y. Diets, gut microbiota and metabolites. Phenomics 2023, 3, 268–284. [Google Scholar] [CrossRef]
- Holman, D.B.; Gzyl, K.E.; Scott, H.; Service, C.; Prieto, N.; López-Campos, Ó. Associations between the rumen microbiota and carcass merit and meat quality in beef cattle. Appl. Microbiol. Biotechnol. 2024, 108, 287. [Google Scholar] [CrossRef]
- Lammers, P.J.; Kerr, B.J.; Honeyman, M.S. Biofuel co-products as swine feed ingredients: Combining corn distillers dried grains with solubles (DDGS) and crude glycerin. Anim. Feed Sci. Technol. 2015, 201, 110–114. [Google Scholar] [CrossRef]
- Eiras, C.E.; Barbosa, L.P.; Marques, J.A.; Araújo, F.L.; Lima, B.S.; Zawadzki, F.; Perotto, D.; Prado, I.N. Glycerine levels in the diets of crossbred bulls finished in feedlot: Apparent digestibility, feed intake and animal performance. Anim. Feed Sci. Technol. 2014, 197, 222–226. [Google Scholar] [CrossRef]
- Schieck, S.J.; Shurson, G.C.; Kerr, B.J.; Johnston, L.J. Evaluation of glycerol, a biodiesel coproduct, in grow-finish pig diets to support growth and pork quality. J. Anim. Sci. 2010, 88, 3927–3935. [Google Scholar] [CrossRef]
- De Weirdt, R.; Crabbe, A.; Roos, S.; Vollenweider, S.; Lacroix, C.; van Pijkeren, J.P.; Britton, R.A.; Sarker, S.; De Wiele, T.V.; Nickerson, C.A. Glycerol supplementation enhances L. reuteri’s protective effect against S. Typhimurium colonization in a 3-D model of colonic epithelium. PLoS ONE 2012, 7, e37116. [Google Scholar] [CrossRef]
- Wu, L.T.; Tan, L.M.; You, C.Y.; Lan, T.Y.; Li, W.X.; Xu, Y.T.; Ren, Z.X.; Ding, Q.; Zhou, C.Y.; Tang, Z.R.; et al. Effects of dietary niacinamide and CP concentrations on the nitrogen excretion, growth performance, and meat quality of pigs. Animal 2023, 17, 100869. [Google Scholar] [CrossRef] [PubMed]
- Akinmoladun, O.F.; Fon, F.N.; Mpendulo, C.T. Stress indicators, carcass characteristics and meat quality of Xhosa goats subjected to different watering regimen and vitamin C supplementation. Livest. Sci. 2020, 238, 104083. [Google Scholar] [CrossRef]
- Goldfarb, A.H.; Bloomer, R.J.; Mckenzie, M.J. Combined antioxidant treatment effects on blood oxidative stress after eccentric exercise. Med. Sci. Sports Exerc. 2005, 37, 234–239. [Google Scholar] [CrossRef]
- Latham, C.M.; Guy, C.P.; Wesolowski, L.T.; White-Springer, S.H. Fueling equine performance: Importance of mitochondrial phenotype in equine athletes. Anim. Front. 2022, 12, 6–14. [Google Scholar] [CrossRef]
- Asare, P.T.; Greppi, A.; Geirnaert, A.; Pennacchia, A.; Babst, A.; Lacroix, C. Glycerol and reuterin-producing Limosilactobacillus reuteri enhance butyrate production and inhibit Enterobacteriaceae in broiler chicken cecal microbiota PolyFermS model. BMC Microbiol. 2023, 23, 384. [Google Scholar] [CrossRef]
- Louvado, A.; Coelho, F.J.R.C.; Palma, M.; Tavares, L.C.; Ozorio, R.O.A.; Magnoni, L.; Viegas, I.; Gomes, N.C.M. Effect of glycerol feed-supplementation on seabass metabolism and gut microbiota. Appl. Microbiol. Biot. 2020, 104, 8439–8453. [Google Scholar] [CrossRef] [PubMed]
- Anwar, F.; Alhayyani, S.; Al-Abbasi, F.A.; Nadeem, M.S.; Kumar, V. Pharmacological role of vitamin C in stress-induced cardiac dysfunction via alteration in gut microbiota. J. Biochem. Mol. Toxicol. 2022, 36, e22986. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Sun, Y.; Pan, D.; Chang, B.; Sang, L.X. Nicotinamide ameliorates dextran sulfate sodium-induced chronic colitis in mice through its anti-inflammatory properties and modulates the gut microbiota. J. Immunol. Res. 2021, 2021, 5084713. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.L.; Wang, Q.P.; Gu, S.; Jin, J.M.; Yang, N. Emerging perspectives in the gut-muscle axis: The gut microbiota and its metabolites as important modulators of meat quality. Microb. Biotechnol. 2024, 17, e14361. [Google Scholar] [CrossRef]
- Lu, S.Y.; Xu, Y.X.; Song, X.H.; Li, J.Y.; Jiang, J.Q.; Qin, C.B.; Wu, K.N.; Cui, K.Q.; Liu, Y.; Liu, Q.Y.; et al. Multi-omics reveal the effects and regulatory mechanism of dietary neutral detergent fiber supplementation on carcass characteristics, amino acid profiles, and meat quality of finishing pigs. Food Chem. 2024, 445, 138765. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.L.; Hao, S.B.; Zou, W.J.; Wei, P.T.; Sun, W.C.; Wu, H.D.; Lu, W.; He, Y.Y. Effects of supplementing growing-finishing crossbred pigs with glycerin, vitamin C and niacinamide on carcass characteristics and meat quality. Animals 2023, 13, 3635. [Google Scholar] [CrossRef]
- Riemer, J.; Hoepken, H.H.; Czerwinska, H.; Robinson, S.R.; Dringen, R. Colorimetric ferrozine-based assay for the quantitation of iron in cultured cells. Anal. Biochem. 2004, 331, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef]
- Li, D.; Liu, C.M.; Luo, R.; Sadakane, K.; Lam, T.W. Megahit: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. Quast: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Zhu, W.; Lomsadze, A.; Borodovsky, M. Ab initio gene identification in metagenomic sequences. Nucleic Acids Res. 2010, 38, 132. [Google Scholar] [CrossRef]
- Steinegger, M.; Söding, J. MMseqs2 enables sensitive protein sequence searching for the analysis of massive data sets. Nat. Biotechnol. 2017, 35, 1026–1028. [Google Scholar] [CrossRef]
- Santos, S.; Neto, I.F.F.; Machado, M.D.; Soares, H.M.V.M.; Soares, E.V. Siderophore production by Bacillus megaterium: Effect of growth phase and cultural conditions. Appl. Biochem. Biotech. 2014, 172, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Virpiranta, H.; Banasik, M.; Taskila, S.; Leiviskä, T.; Halttu, M.; Sotaniemi, V.; Tanskanen, J. Isolation of efficient metal-binding bacteria from boreal peat soils and development of microbial biosorbents for improved nickel scavenging. Water 2020, 12, 2000. [Google Scholar] [CrossRef]
- Colin, T.; Bories, A.; Lavigne, C.; Moulin, G. Effects of Acetate and Butyrate During Glycerol Fermentation by Clostridium butyricum. Curr. Microbiol. 2001, 43, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.J.; Yin, F.G.; Yang, Y.H.; Lepp, D.; Yu, H.; Ruan, Z.; Yang, C.B.; Yin, Y.L.; Hou, Y.Q.; Leeson, S.; et al. Dietary butyrate glycerides modulate intestinal microbiota composition and serum metabolites in broilers. Sci. Rep. 2018, 8, 4940. [Google Scholar] [CrossRef]
- Dishisha, T.; Pereyra, L.P.; Pyo, S.H.; Britton, R.A.; Hatti-Kaul, R. Flux analysis of the Lactobacillus reuteri propanediol-utilization pathway for production of 3-hydroxypropionaldehyde, 3-hydroxypropionic acid and 1,3-propanediol from glycerol. Microb. Cell Fact. 2014, 13, 76. [Google Scholar] [CrossRef] [PubMed]
- Otten, A.T.; Bourgonje, A.R.; Peters, V.; Alizadeh, B.Z.; Dijkstra, G.; Harmsen, H.J.M. Vitamin C supplementation in healthy individuals leads to shifts of bacterial populations in the gut-A pilot study. Antioxidants 2021, 10, 1278. [Google Scholar] [CrossRef]
- He, Z.Y.; Li, X.M.; Yang, H.X.; Wu, P.; Wang, S.S.; Cao, D.; Guo, X.X.; Xu, Z.R.; Gao, J.Y.; Zhang, W.Y.; et al. Effects of oral vitamin C supplementation on liver health and associated parameters in patients with non-alcoholic fatty liver disease: A randomized clinical trial. Front. Nutr. 2021, 8, 745609. [Google Scholar] [CrossRef]
- Jaeggi, T.; Kortman, G.A.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H.; et al. Iron fortification adversely affects the gut microbiome, increases pathogen abundance and induces intestinal inflammation in Kenyan infants. Gut 2015, 64, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Kalipatnapu, S.; Kuppuswamy, S.; Venugopal, G.; Kaliaperumal, V.; Ramadass, B. Fecal total iron concentration is inversely associated with fecal Lactobacillus in preschool children. J. Gastroenterol. Hepatol. 2017, 32, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Fang, S.M.; Wei, H.; He, M.Z.; Fu, H.; Xiong, X.W.; Zhou, Y.Y.; Wu, J.Y.; Gao, J.; Yang, H.; et al. Prevotella copri increases fat accumulation in pigs fed with formula diets. Microbiome 2021, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Mailloux, R.J. Proline and dihydroorotate dehydrogenase promote a hyper-proliferative state and dampen ferroptosis in cancer cells by rewiring mitochondrial redox metabolism. BBA-Mol. Cell Res. 2024, 1871, 119639. [Google Scholar] [CrossRef]
- Rodríguez-Graciani, K.; Chapa-Dubocq, X.; Chorna, N.; Javadov, S. Metabolomic profile of ferroptosis in cardiac cells. FASEB J. 2021, 35, 1–3. [Google Scholar] [CrossRef]
- Chibani, C.M.; Mahnert, A.; Borrel, G.; Almeida, A.; Werner, A.; Brugere, J.F.; Gribaldo, S.; Finn, R.D.; Schmitz, R.A.; Moissl-Eichinger, C. A catalogue of 1,167 genomes from the human gut archaeome. Nat. Microbiol. 2022, 7, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Christgen, S.L.; Becker, D.F. Role of proline in pathogen and host interactions. Antioxid. Redox Sign. 2019, 30, 683–709. [Google Scholar] [CrossRef] [PubMed]
- Watt, A.E.; Browning, G.F.; Legione, A.R.; Bushell, R.N.; Stent, A.; Cutler, R.S.; Young, N.D.; Marenda, M.S. A novel Glaesserella sp. isolated from pigs with severe respiratory infections has a mosaic genome with virulence factors putatively acquired by horizontal transfer. Appl. Environ. Microb. 2018, 84, e00092-18. [Google Scholar] [CrossRef]
- Cassat, J.E.; Skaar, E.P. Iron in infection and immunity. Cell Host Microbe 2013, 13, 509–519. [Google Scholar] [CrossRef]
- Fardeau, S.; Mullié, C.; Dassonville-Klimpt, A.; Audic, N.; Sasaki, A.; Sonnet, P. Bacterial iron uptake: A promising solution against multidrug resistant bacteria. In Science against Microbial Pathogens: Communicating Current Research and Technological Advances; Méndez-Vilas, A., Ed.; Microbiology book series number 3; Formatex Research Center: Badajoz, Spain, 2011; Volume 2, pp. 695–705. [Google Scholar]
- Su, H.C.; Hu, X.J.; Xu, W.J.; Xu, Y.; Wen, G.L.; Cao, Y.C. Diversity, abundances and distribution of antibiotic resistance genes and virulence factors in the south China Sea revealed by metagenomic sequencing. Sci. Total Environ. 2022, 814, 152803. [Google Scholar] [CrossRef]
- Lee, C.R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, K., II; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, antibiotic resistance mechanisms, and prospective treatment options. Front. Cell Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef]
- Ciancio, M.J.; Chang, E.B. Do heat shock proteins play any role in gut inflammation? Inflamm. Bowel Dis. 2010, 14, 102–103. [Google Scholar] [CrossRef]
- Lee, S. Bacteriocins of Listeria monocytogenes and their potential as a virulence factor. Toxins 2020, 12, 103. [Google Scholar] [CrossRef] [PubMed]
- Molloy, E.M.; Casjens, S.R.; Cox, C.L.; Maxson, T.; Ethridge, N.A.; Margos, G.; Fingerle, V.; Mitchell, D.A. Identification of the minimal cytolytic unit for Streptolysin S and an expansion of the toxin family. BMC Microbiol. 2015, 15, 141. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, D.J.; Lee, S.W.; Hensler, M.E.; Markley, A.L.; Dahesh, S.; Mitchell, D.A.; Bandeira, N.; Nizet, V.; Dixon, J.E.; Dorrestein, P.C. Clostridiolysin S, a post-translationally modified biotoxin from Clostridium botulinum. J. Biol. Chem. 2010, 285, 28220–28228. [Google Scholar] [CrossRef]
- Clayton, E.M.; Daly, K.M.; Guinane, C.M.; Hill, C.; Cotter, P.D.; Ross, P.R. Atypical Listeria innocua strains possess an intact LIPI-3. BMC Microbiol. 2014, 14, 58. [Google Scholar] [CrossRef]
- Quereda, J.J.; Dussurget, O.; Nahori, M.A.; Ghozlane, A.; Volant, S.; Dillies, M.A.; Regnault, B.; Kennedy, S.; Mondot, S.; Villoing, B.; et al. Bacteriocin from epidemic Listeria strains alters the host intestinal microbiota to favor infection. Proc. Natl. Acad. Sci. USA 2016, 113, 5706–5711. [Google Scholar] [CrossRef]
- Fuchs, C.D.; Trauner, M. Role of bile acids and their receptors in gastrointestinal and hepatic pathophysiology. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 432–450. [Google Scholar] [CrossRef]
- Collins, S.L.; Stine, J.G.; Bisanz, J.E.; Okafor, C.D.; Patterson, A.D. Bile acids and the gut microbiota: Metabolic interactions and impacts on disease. Nat. Rev. Microbiol. 2023, 21, 236–247. [Google Scholar] [CrossRef]
- Thanissery, R.; Winston, J.A.; Theriot, C.M. Inhibition of spore germination, growth, and toxin activity of clinically relevant C. difficile strains by gut microbiota derived secondary bile acids. Anaerobe 2017, 45, 86–100. [Google Scholar] [CrossRef]
- Buffie, C.G.; Bucci, V.; Stein, R.R.; McKenney, P.T.; Ling, L.; Gobourne, A.; No, D.; Liu, H.; Kinnebrew, M.; Viale, A.; et al. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature 2015, 517, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.D.; Myers, C.J.; Harris, S.C.; Kakiyama, G.; Lee, I.K.; Yun, B.S.; Matsuzaki, K.; Furukawa, M.; Min, H.K.; Bajaj, J.S.; et al. Bile acid 7α-dehydroxylating gut bacteria secrete antibiotics that inhibit Clostridium difficile: Role of secondary bile acids. Cell Chem. Biol. 2019, 26, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, J.U.; Huber, T. Structural evolution of the ABC transporter subfamily B. Evolut. Bioinform. 2007, 3, 309–316. [Google Scholar] [CrossRef]
- Abe, K.; Obana, N.; Nakamura, K. Effects of depletion of RNA-binding protein Tex on the expression of toxin genes in Clostridium perfringens. Biosci. Biotechnol. Biochem. 2010, 74, 1564–1571. [Google Scholar] [CrossRef] [PubMed]
- Star, C.R.; Engleberg, N.C. Role of hyaluronidase in subcutaneous spread and growth of group A streptococcus. Infect. Immun. 2006, 74, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Kaul, A.; Short, W.D.; Wang, X.Y.; Keswani, S.G. Hyaluronidases in human diseases. Int. J. Mol. Sci. 2021, 22, 3204. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.J.; Chang, W.P.; Gao, L.; Deng, L.; Ji, W.K. Tex2 is required for lysosomal functions at TMEM55-dependent ER membrane contact sites. J. Cell Biol. 2023, 222, e202205133. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Li, M.H.; Duan, H.M.; Li, L.Y.; Ouyang, P.; Chen, D.F.; Geng, Y.; Huang, X.L.; Yang, S.Y.; Yin, L.Z.; et al. Microbial analysis reveals the potential colonization of pathogens in the intestine of crayfish (Procambarus clarkii) in traditional aquaculture environments. Ecotoxicol. Environ. Saf. 2021, 224, 112705. [Google Scholar] [CrossRef] [PubMed]
- Oehme, D.P.; Shafee, T.; Downton, M.T.; Bacic, A.; Doblin, M.S.; Gallicchio, E. Differences in protein structural regions that impact functional specificity in GT2 family β-glucan synthases. PLoS ONE 2019, 14, e0224442. [Google Scholar] [CrossRef]
- Bockwoldt, J.A.; Meng, C.; Ludwig, C.; Kupetz, M.; Ehrmann, M.A. Proteomic Analysis Reveals Enzymes for β-D-Glucan Formation and Degradation in Levilactobacillus brevis TMW 1.2112. Int. J. Mol. Sci. 2022, 23, 3393. [Google Scholar] [CrossRef]
- Isono, N.; Mizutani, E.; Hayashida, H.; Katsuzaki, H.; Saburi, W. Functional characterization of a novel GH94 glycoside phosphorylase, 3-O-β-d-glucopyranosyl β-d-glucuronide phosphorylase, and implication of the metabolic pathway of acidic carbohydrates in Paenibacillus borealis. Biochem. Biophys. Res. Commun. 2022, 625, 60–65. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group A | Group B | Group C | Group D | |||||
|---|---|---|---|---|---|---|---|---|
| 10–60 kg | 60–120 kg | 10–60 kg | 60–120 kg | 10–60 kg | 60–120 kg | 10–60 kg | 60–120 kg | |
| Ingredients | ||||||||
| Corn | 59.00 | 64.00 | 48.00 | 52.00 | 58.89 | 63.89 | 47.89 | 51.89 |
| Wheat bran | 16.00 | 16.00 | 15.00 | 15.00 | 16.00 | 16.00 | 15.00 | 15.00 |
| Soybean meal | 14.00 | 10.00 | 15.00 | 11.00 | 14.00 | 10.00 | 15.00 | 11.00 |
| Fishmeal | 2.00 | 0.00 | 2.00 | 0.00 | 2.00 | 0.00 | 2.00 | 0.00 |
| Rapeseed meal | 5.00 | 6.00 | 6.00 | 8.00 | 5.00 | 6.00 | 6.00 | 8.00 |
| 4% Premix (1) | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 |
| Glycerin | 0.00 | 0.00 | 10.00 | 10.00 | 0.00 | 0.00 | 10.00 | 10.00 |
| Vitamin C | 0.00 | 0.00 | 0.00 | 0.00 | 0.06 | 0.06 | 0.06 | 0.06 |
| Niacinamide | 0.00 | 0.00 | 0.00 | 0.00 | 0.05 | 0.05 | 0.05 | 0.05 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 |
| Nutrient levels (2) | ||||||||
| Metabolizable energy (MJ/kg) | 12.15 | 12.24 | 12.26 | 12.31 | 12.15 | 12.24 | 12.26 | 12.31 |
| Crude protein | 17.30 | 15.10 | 17.34 | 15.18 | 17.22 | 15.05 | 17.36 | 15.21 |
| Crude fiber | 3.84 | 3.68 | 3.69 | 3.78 | 3.74 | 3.71 | 3.47 | 3.63 |
| Ether extract | 3.81 | 3.68 | 3.43 | 3.50 | 3.94 | 3.88 | 3.57 | 3.65 |
| Calcium | 0.79 | 0.65 | 0.82 | 0.70 | 0.73 | 0.69 | 0.78 | 0.63 |
| Total phosphorus | 0.67 | 0.54 | 0.63 | 0.55 | 0.69 | 0.59 | 0.66 | 0.60 |
| Lysine | 1.09 | 0.84 | 1.09 | 0.86 | 1.09 | 0.84 | 1.09 | 0.86 |
| Methionine + Cystine | 0.57 | 0.44 | 0.56 | 0.43 | 0.57 | 0.44 | 0.56 | 0.43 |
| Group A | Group B | Group C | Group D | SEM | p-Value | |||
|---|---|---|---|---|---|---|---|---|
| GLY | VC + NAM | GLY × (VC + NAM) | ||||||
| Glycerol (µg/g) | 122.02 | 117.16 | 111.22 | 100.29 | 19.70 | 0.325 | 0.092 | 0.702 |
| Ferric ion (µg/g) | 43.35 b | 32.04 c | 37.84 bc | 54.30 a | 2.15 | 0.363 | 0.007 | <0.001 |
| Ferrous ion (µg/g) | 12.69 ab | 10.45 b | 11.41 ab | 15.27 a | 3.40 | 0.524 | 0.173 | 0.024 |
| Group A | Group B | Group C | Group D | SEM | p-Value | |
|---|---|---|---|---|---|---|
| Ace | 9612.02 a | 9072.30 ab | 9048.48 ab | 8129.06 b | 896.70 | 0.024 |
| Chao 1 | 9584.16 a | 9046.86 ab | 9025.50 ab | 8121.57 b | 891.96 | 0.027 |
| Shannon | 6.42 | 6.53 | 6.45 | 6.47 | 0.12 | 0.467 |
| Simpson | 0.99 | 0.99 | 0.99 | 0.99 | 0.00 | 0.152 |
| Differential Microbiota | Iron Uptake-Related Components of Differential Microbiota | |
|---|---|---|
| Group D | s_Methanobrevibacter_sp._YE315 | ABC transporter permease |
| g_Methanobrevibacter | Siderophore transport system ATP-binding protein YusV | |
| Ferredoxin oxidoreductase | ||
| Ferrous iron transport protein B | ||
| Desulfoferrodoxin | ||
| ABC transporter permease | ||
| ABC transporter ATP-binding protein | ||
| Iron ABC transporter substrate-binding protein | ||
| Iron ABC transporter permease | ||
| Iron ABC transporter permease | ||
| g_Clostridium | Siderophore/surfactin synthetase-related protein | |
| Siderophore | ||
| Siderophore transport system permease protein yfhA | ||
| Siderophore transport system ATP-binding protein YusV | ||
| Pyruvate:ferredoxin (flavodoxin) oxidoreductase | ||
| Ferredoxin | ||
| 4Fe-4S ferredoxin | ||
| ABC transporter ATP-binding protein | ||
| ABC transporter permease | ||
| Ferrous iron transporter | ||
| f_Clostridiaceae | Pyruvate ferredoxin oxidoreductase | |
| Ferredoxin | ||
| ABC transporter ATP-binding protein | ||
| ABC transporter permease | ||
| ABC transporter substrate-binding protein | ||
| s_Clostridium sp. CAG:1000 | ABC transporter ATP-binding protein | |
| ABC-type transport system involved in Fe-S cluster assembly permease component | ||
| ABC transporter-related protein | ||
| ABC transporter permease protein | ||
| s_Methanobrevibacter_sp._YE315 | ABC transporter permease | |
| s_Alistipes_sp._CAG_435 | Ferritin | |
| 2-oxoacid:ferredoxin oxidoreductase γ-subunit | ||
| ABC transporter ATP-binding protein | ||
| Efflux ABC transporter permease protein | ||
| s_Alistipes_sp._CAG_514 | Ferredoxin | |
| Ferredoxin-NADP+ reductase subunit-α | ||
| ABC transporter ATP-binding protein | ||
| s_Faecalibacterium_sp. CAG:74_58_120 | ABC transporter ATP-binding protein | |
| ABC transporter permease | ||
| ABC transporter substrate-binding protein | ||
| Group A | s_Prevotella_copri | Iron ABC transporter permease |
| ABC transporter substrate-binding protein | ||
| ABC transporter permease | ||
| g_Dorea | Metal ABC transporter permease | |
| ABC transporter ATP-binding protein | ||
| g_Coprococcus | Ferredoxin | |
| ABC transporter ATP-binding protein | ||
| ABC transporter permease |
| #ID | Name | Groups | Fold Change (FC) | log2FC | p-Value | VIP |
|---|---|---|---|---|---|---|
| ABC transporters (ko02010) | ||||||
| pos_2704 | l-Proline | B/A | 1.3386 | 0.4208 | <0.0000 | 2.3914 |
| neg_1618 | l-Proline | C/A | 2.4404 | 1.2871 | 0.0038 | 2.2233 |
| pos_2704 | l-Proline | D/A | 1.4592 | 0.5451 | 0.0037 | 2.0634 |
| neg_1618 | l-Proline | C/B | 6.2859 | 2.6521 | 0.0007 | 2.2128 |
| pos_2704 | l-Proline | D/C | 1.3624 | 0.4461 | 0.0065 | 2.0721 |
| Mineral absorption (ko04978) | ||||||
| pos_2704 | l-Proline | B/A | 1.3386 | 0.4208 | <0.0000 | 2.3914 |
| neg_1618 | l-Proline | C/A | 2.4404 | 1.2871 | 0.0038 | 2.2233 |
| neg_1957 | l-Tryptophan | C/A | 2.7506 | 1.4597 | 0.0004 | 2.5711 |
| neg_3312 | Calcitriol | D/A | 0.3881 | −1.3658 | 0.0053 | 1.8416 |
| neg_1957 | l-Tryptophan | C/B | 4.6449 | 2.2156 | <0.0000 | 2.3925 |
| neg_1618 | l-Proline | C/B | 6.2859 | 2.6521 | 0.0007 | 2.2128 |
| pos_2704 | l-Proline | D/C | 1.3624 | 0.4461 | 0.0065 | 2.0721 |
| Name | Category | Concentration in Feces (ng/g) | Fold Change | p-Value | |
|---|---|---|---|---|---|
| Group B | Group A | ||||
| DCA | Secondary bile acid | 445.87 ± 78.89 | 94.42 ± 30.18 | 4.72 | 0.0019 |
| GCA | Primary bile acid | 6.78 ± 0.90 | 2.59 ± 0.62 | 2.62 | 0.0032 |
| α-MCA | Primary bile acid | 84.50 ± 16.45 | 137.31 ± 15.81 | 0.62 | 0.0431 |
| GDCA | Secondary bile acid | 4.53 ± 0.40 | 2.03 ± 0.36 | 2.25 | 0.0008 |
| Group C | Group A | ||||
| DCA | Secondary bile acid | 442.21 ± 94.41 | 94.42 ± 30.18 | 4.68 | 0.0127 |
| TUDCA | Secondary bile acid | 49.10 ± 3.13 | 15.65 ± 1.53 | 3.14 | <0.0000 |
| HCA | Primary bile acid | 335.59 ± 31.15 | 646.12 ± 95.80 | 0.52 | 0.0214 |
| Group D | Group A | ||||
| alloLCA | Secondary bile acid | 6455.71 ± 509.98 | 4193.75 ± 335.40 | 1.54 | 0.0041 |
| DCA | Secondary bile acid | 811.36 ± 188.50 | 94.42 ± 30.18 | 8.59 | 0.0121 |
| TUDCA | Secondary bile acid | 37.25 ± 3.28 | 15.65 ± 1.53 | 2.38 | 0.0001 |
| GDCA | Secondary bile acid | 6.52 ± 1.41 | 2.03 ± 0.36 | 3.21 | 0.0235 |
| Group B | Group C | ||||
| alloLCA | Secondary bile acid | 3670.87 ± 316.31 | 5505.35 ± 504.44 | 0.67 | 0.0116 |
| TUDCA | Secondary bile acid | 15.29 ± 1.99 | 49.10 ± 3.13 | 0.31 | <0.0000 |
| Group B | Group D | ||||
| alloLCA | Secondary bile acid | 3670.87 ± 316.31 | 6455.71 ± 509.98 | 0.57 | 0.0009 |
| TUDCA | Secondary bile acid | 15.29 ± 1.99 | 37.25 ± 3.28 | 0.41 | 0.0002 |
| Group C | Group D | ||||
| TUDCA | Secondary bile acid | 49.10 ± 3.13 | 37.25 ± 3.28 | 1.32 | 0.0259 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, P.; Sun, W.; Hao, S.; Deng, L.; Zou, W.; Wu, H.; Lu, W.; He, Y. Dietary Supplementation of Crossbred Pigs with Glycerol, Vitamin C, and Niacinamide Alters the Composition of Gut Flora and Gut Flora-Derived Metabolites. Animals 2024, 14, 2198. https://doi.org/10.3390/ani14152198
Wei P, Sun W, Hao S, Deng L, Zou W, Wu H, Lu W, He Y. Dietary Supplementation of Crossbred Pigs with Glycerol, Vitamin C, and Niacinamide Alters the Composition of Gut Flora and Gut Flora-Derived Metabolites. Animals. 2024; 14(15):2198. https://doi.org/10.3390/ani14152198
Chicago/Turabian StyleWei, Panting, Wenchen Sun, Shaobin Hao, Linglan Deng, Wanjie Zou, Huadong Wu, Wei Lu, and Yuyong He. 2024. "Dietary Supplementation of Crossbred Pigs with Glycerol, Vitamin C, and Niacinamide Alters the Composition of Gut Flora and Gut Flora-Derived Metabolites" Animals 14, no. 15: 2198. https://doi.org/10.3390/ani14152198