
Rose or Red, but Still under Threat: Comparing Microplastics Ingestion between Two Sympatric Marine Crustacean Species (Aristaeomorpha foliacea and Parapenaeus longirostris)
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Sample Collection and Analytical Methods
2.2.1. Samples Processing
2.2.2. Stable Isotope Analysis
2.2.3. Microplastics Analysis
2.2.4. Quality Control for Microplastics Analysis
2.3. Statistical Analysis
3. Results
3.1. Stable Isotope Analysis
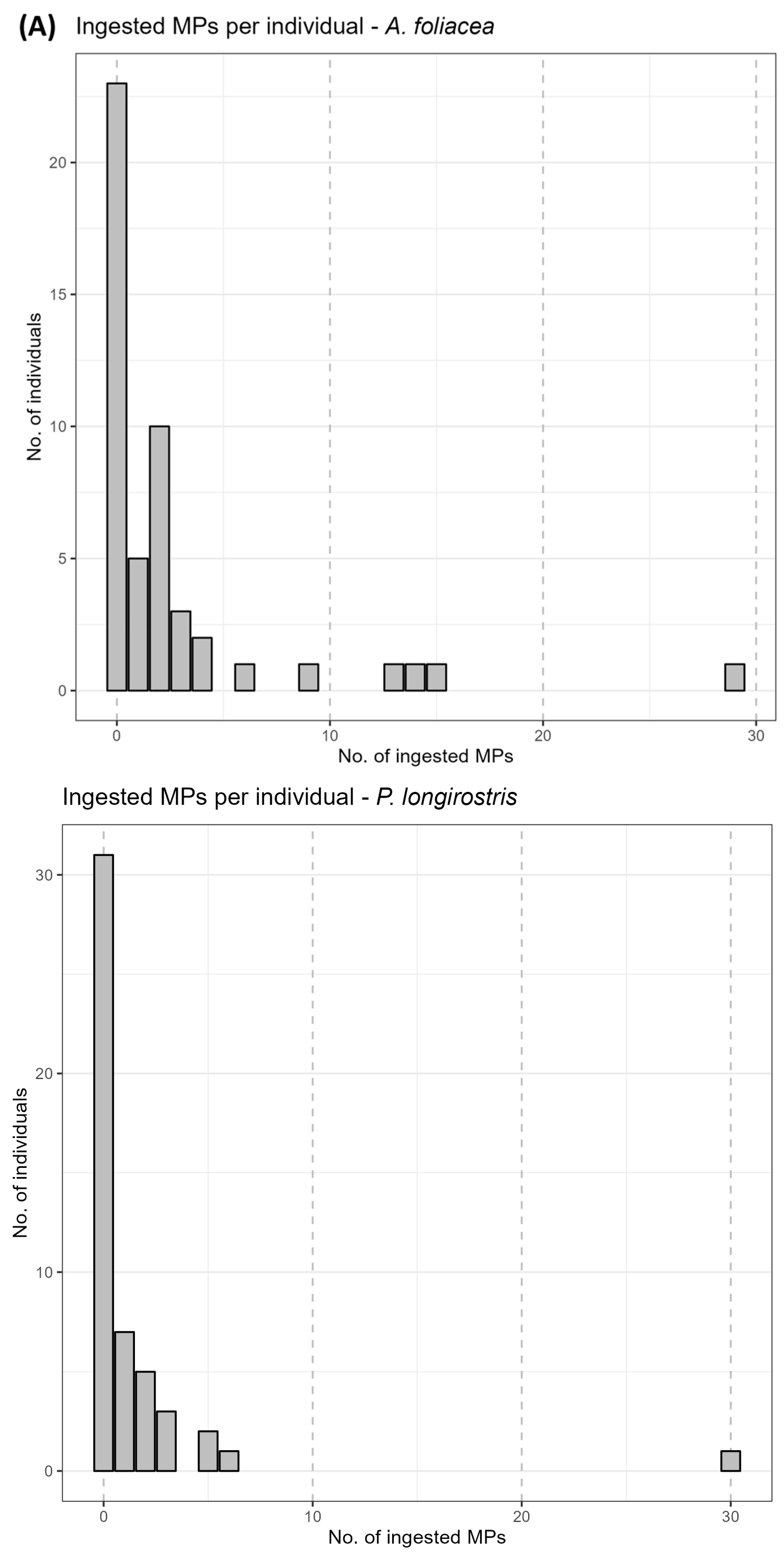
3.2. Microplastics Ingestion
Microplastics Characterisation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chamas, A.; Moon, H.; Zheng, J.; Qiu, Y.; Tabassum, T.; Jang, J.H.; Abu-Omar, M.; Scott, S.L.; Suh, S. Degradation Rates of Plastics in the Environment. ACS Sustain. Chem. Eng. 2020, 8, 3494–3511. [Google Scholar] [CrossRef]
- Bergmann, M.; Gutow, L.; Klages, M. Marine Anthropogenic Litter, 1st ed.; Springer Nature: Berlin, Germany, 2015; pp. 29–75. [Google Scholar]
- Gao, A.L.; Wan, Y. Life Cycle Assessment of Environmental Impact of Disposable Drinking Straws: A Trade-off Analysis with Marine Litter in the United States. Sci. Total Environ. 2022, 817, 153016. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Awasthi, A.K.; Wei, F.; Tan, Q.; Li, J. Single-Use Plastics: Production, Usage, Disposal, and Adverse Impacts. Sci. Total Environ. 2021, 752, 141772. [Google Scholar] [CrossRef] [PubMed]
- IUCN: International Union for Conservation of Nature. Marine Plastic Pollution; IUCN: Gland, Switzerland, 2021. [Google Scholar]
- Ciaralli, L.; Rotini, A.; Scalici, M.; Battisti, C.; Chiesa, S.; Christoforou, E.; Libralato, G.; Manfra, L. The Under-Investigated Plastic Threat on Seagrasses Worldwide: A Comprehensive Review. Environ. Sci. Pollut. Res. 2024, 31, 8341–8353. [Google Scholar] [CrossRef] [PubMed]
- Gouin, T. Toward an Improved Understanding of the Ingestion and Trophic Transfer of Microplastic Particles: Critical Review and Implications for Future Research. Environ. Toxicol. Chem. 2020, 39, 1119–1137. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.; Olsen, Y.; Mitchell, R.P.; Davis, A.; Rowland, S.J.; John, A.W.G.; McGonigle, D.; Russell, A.E. Lost at Sea: Where Is All the Plastic? Science 2004, 304, 838. [Google Scholar] [CrossRef]
- Aigars, J.; Barone, M.; Suhareva, N.; Putna-Nimane, I.; Dimante-Deimantovica, I. Occurrence and Spatial Distribution of Microplastics in the Surface Waters of the Baltic Sea and the Gulf of Riga. Mar. Pollut. Bull. 2021, 172, 112860. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, C.; Roscher, L.; Meyer, M.S.; Hildebrandt, L.; Prume, J.; Löder, M.G.J.; Primpke, S.; Gerdts, G. Spatial Distribution of Microplastics in Sediments and Surface Waters of the Southern North Sea. Environ. Pollut. 2019, 252, 1719–1729. [Google Scholar] [CrossRef]
- Wagner, S.; Klöckner, P.; Stier, B.; Römer, M.; Seiwert, B.; Reemtsma, T.; Schmidt, C. Relationship between Discharge and River Plastic Concentrations in a Rural and an Urban Catchment. Environ. Sci. Technol. 2019, 53, 10082–10091. [Google Scholar] [CrossRef] [PubMed]
- Cózar, A.; Sanz-Martín, M.; Martí, E.; González-Gordillo, J.I.; Ubeda, B.; Gálvez, J.Á.; Irigoien, X.; Duarte, C.M. Plastic Accumulation in the Mediterranean Sea. PLoS ONE 2015, 10, e0121762. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed]
- Cuttelod, A.; García, N.; Malak Abdul, D.; Temple, H.; Katariya, V. The Mediterranean: A Biodiversity Hotspot under Threat. In Wildlife in a Changing World–An Analysis of the 2008 IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2008. [Google Scholar]
- Sharma, S.; Sharma, V.; Chatterjee, S. Microplastics in the Mediterranean Sea: Sources, Pollution Intensity, Sea Health, and Regulatory Policies. Front. Mar. Sci. 2021, 8, 634934. [Google Scholar] [CrossRef]
- Suaria, G.; Avio, C.G.; Mineo, A.; Lattin, G.L.; Magaldi, M.G.; Belmonte, G.; Moore, C.J.; Regoli, F.; Aliani, S. The Mediterranean Plastic Soup: Synthetic Polymers in Mediterranean Surface Waters. Sci. Rep. 2016, 6, 37551. [Google Scholar] [CrossRef] [PubMed]
- Boudouresque, C.F.; Bernard, G.; Pergent, G.; Shili, A.; Verlaque, M. Regression of Mediterranean Seagrasses Caused by Natural Processes and Anthropogenic Disturbances and Stress: A Critical Review. Bot. Mar. 2009, 52, 395–418. [Google Scholar] [CrossRef]
- Danovaro, R. Pollution Threats in the Mediterranean Sea: An Overview. Chem. Ecol. 2003, 19, 15–32. [Google Scholar] [CrossRef]
- Inghilesi, R.; Ottolenghi, L.; Orasi, A.; Pizzi, C.; Bignami, F.; Santoleri, R. Fate of River Tiber Discharge Investigated through Numerical Simulation and Satellite Monitoring. Ocean Sci. 2012, 8, 773–786. [Google Scholar] [CrossRef]
- Cesarini, G.; Crosti, R.; Secco, S.; Gallitelli, L.; Scalici, M. From City to Sea: Spatiotemporal Dynamics of Floating Macrolitter in the Tiber River. Sci. Total Environ. 2023, 857, 159713. [Google Scholar] [CrossRef] [PubMed]
- De Lucia, G.; Vianello, A.; Camedda, A.; Vani, D.; Tomassetti, P.; Coppa, S.; Palazzo, L.; Amici, M.; Romanelli, G.; Zampetti, G.; et al. Sea Water Contamination in the Vicinity of the Italian Minor Islands Caused by Microplastic Pollution. Water 2018, 10, 1108. [Google Scholar] [CrossRef]
- Crosti, R.; Arcangeli, A.; Campana, I.; Paraboschi, M.; González-Fernández, D. ‘Down to the River’: Amount, Composition, and Economic Sector of Litter Entering the Marine Compartment, through the Tiber River in the Western Mediterranean Sea. Rend. Lincei Sci. Fis. Nat. 2018, 29, 859–866. [Google Scholar] [CrossRef]
- Arthur, C.; Baker, J.; Bamford, H. Occurrence, Effects, and Fate of Microplastic Marine Debris. In Proceedings of the International Research Workshop, Tacoma, WA, USA, 9–11 September 2009. [Google Scholar]
- Fossi, M.C.; Pedà, C.; Compa, M.; Tsangaris, C.; Alomar, C.; Claro, F.; Ioakeimidis, C.; Galgani, F.; Hema, T.; Deudero, S.; et al. Bioindicators for Monitoring Marine Litter Ingestion and Its Impacts on Mediterranean Biodiversity. Environ. Pollut. 2018, 237, 1023–1040. [Google Scholar] [CrossRef]
- Sbrana, A.; Valente, T.; Scacco, U.; Bianchi, J.; Silvestri, C.; Palazzo, L.; De Lucia, G.A.; Valerani, C.; Ardizzone, G.; Matiddi, M. Spatial Variability and Influence of Biological Parameters on Microplastic Ingestion by Boops boops (L.) along the Italian Coasts (Western Mediterranean Sea). Environ. Pollut. 2020, 263, 114429. [Google Scholar] [CrossRef] [PubMed]
- Abidli, S.; Pinheiro, M.; Lahbib, Y.; Neuparth, T.; Santos, M.M.; Trigui El Menif, N. Effects of Environmentally Relevant Levels of Polyethylene Microplastic on Mytilus galloprovincialis (Mollusca: Bivalvia): Filtration Rate and Oxidative Stress. Environ. Sci Pollut. Res. 2021, 28, 26643–26652. [Google Scholar] [CrossRef]
- Valente, T.; Costantini, M.L.; Careddu, G.; Berto, D.; Piermarini, R.; Rampazzo, F.; Sbrana, A.; Silvestri, C.; Ventura, D.; Matiddi, M. Tracing the Route: Using Stable Isotope Analysis to Understand Microplastic Pathways through the Pelagic-Neritic Food Web of the Tyrrhenian Sea (Western Mediterranean). Sci. Total Environ. 2023, 885, 163875. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Nag, R.; Cummins, E. Human Health Concerns Regarding Microplastics in the Aquatic Environment—From Marine to Food Systems. Sci. Total Environ. 2022, 823, 153730. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Nakaoka, M. Trophic Transfer of Microplastics from Mysids to Fish Greatly Exceeds Direct Ingestion from the Water Column. Environ. Pollut. 2021, 273, 116468. [Google Scholar] [CrossRef]
- Nelms, S.E.; Galloway, T.S.; Godley, B.J.; Jarvis, D.S.; Lindeque, P.K. Investigating Microplastic Trophic Transfer in Marine Top Predators. Environ. Pollut. 2018, 238, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Paul-Pont, I.; Tallec, K.; Gonzalez-Fernandez, C.; Lambert, C.; Vincent, D.; Mazurais, D.; Zambonino-Infante, J.-L.; Brotons, G.; Lagarde, F.; Fabioux, C.; et al. Constraints and Priorities for Conducting Experimental Exposures of Marine Organisms to Microplastics. Front. Mar. Sci. 2018, 5, 252. [Google Scholar] [CrossRef]
- Oliveira, M.; Ribeiro, A.; Hylland, K.; Guilhermino, L. Single and Combined Effects of Microplastics and Pyrene on Juveniles (0+ Group) of the Common Goby Pomatoschistus microps (Teleostei, Gobiidae). Ecol. Indic. 2013, 34, 641–647. [Google Scholar] [CrossRef]
- Li, B.; Su, L.; Zhang, H.; Deng, H.; Chen, Q.; Shi, H. Microplastics in Fishes and Their Living Environments Surrounding a Plastic Production Area. Sci. Total Environ. 2020, 727, 138662. [Google Scholar] [CrossRef]
- Timilsina, A.; Adhikari, K.; Yadav, A.K.; Joshi, P.; Ramena, G.; Bohara, K. Effects of Microplastics and Nanoplastics in Shrimp: Mechanisms of Plastic Particle and Contaminant Distribution and Subsequent Effects after Uptake. Sci. Total Environ. 2023, 894, 164999. [Google Scholar] [CrossRef]
- Gallo, F.; Fossi, C.; Weber, R.; Santillo, D.; Sousa, J.; Ingram, I.; Nadal, A.; Romano, D. Marine Litter Plastics and Microplastics and Their Toxic Chemicals Components: The Need for Urgent Preventive Measures. Environ. Sci. Eur. 2018, 30, 13. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.O.; Abrantes, N.; Gonçalves, F.J.M.; Nogueira, H.; Marques, J.C.; Gonçalves, A.M.M. Impacts of Plastic Products Used in Daily Life on the Environment and Human Health: What Is Known? Environ. Toxicol. Pharmacol. 2019, 72, 103239. [Google Scholar] [CrossRef] [PubMed]
- Alberghini, L.; Truant, A.; Santonicola, S.; Colavita, G.; Giaccone, V. Microplastics in Fish and Fishery Products and Risks for Human Health: A Review. Int. J. Environ. Res. Public Health 2022, 20, 789. [Google Scholar] [CrossRef] [PubMed]
- Rochman, C.M.; Hoh, E.; Kurobe, T.; Teh, S.J. Ingested Plastic Transfers Hazardous Chemicals to Fish and Induces Hepatic Stress. Sci. Rep. 2013, 3, 3263. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Santos, T.; Costa, M.; Mouneyrac, C. (Eds.) Handbook of Microplastics in the Environment; Springer International Publishing: Cham, Switzerland, 2020; ISBN 978-3-030-10618-8. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; FAO: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Cau, A.; Carbonell, A.; Follesa, M.C.; Mannini, A.; Orsi Relini, L.; Politou, C.Y.; Ragonese, S.; Rinelli, P. MEDITS-Based Information on the Deep Water Red Shrimps Aristaeomorpha foliacea and Aristeus antennatus (Crustacea: Decapoda: Aristeidae). Sci. Mar. 2002, 66, 103. [Google Scholar] [CrossRef]
- Fischer, W.; Bauchot, M.L.; Schneider, M. Fiches FAO d’identification Des Espèces Pour Les Besoins de La Pêche. (Révision 1); Méditerranée et Mer Noire. Zone de Pêche 37. Végétaux et Invertébrés. Publication Préparée Par La FAO, Résultat d’un Accord Entre La FAO et La Commission Des Communautés Européennes; Département Des pêches de la FAO (Food and Agriculture Organization of the United Nations): Rome, Italy, 1987. [Google Scholar]
- Tom, M.; Goren, M.; Ovadia, M. The Benthic Phase of the Life Cycle of Parapenaeus longirostris (Crustacea, Decapoda, Penaeidae) along the Mediterranean Coast of Israel. Hydrobiologia 1988, 169, 339–352. [Google Scholar] [CrossRef]
- Bianchini, M.; Ragonese, S. Life Cycles and Fisheries of the Deep Water Shrimps Aristaeomorpha foliacea and Aristeus antennatus. In Proceedings of the International Workshop Held in the Istituto Tecnologia Pesca e Pescato, Mazara, Italy, 28–30 April 1994; Volume 3, pp. 1–88. [Google Scholar]
- Cartes, J. Diets of, and Trophic Resources Exploited by, Bathyal Penaeoidean Shrimps from the Western Mediterranean. Mar. Freshw. Res. 1995, 46, 889. [Google Scholar] [CrossRef]
- Cartes, J.E.; Fanelli, E.; Kapiris, K.; Bayhan, Y.K.; Ligas, A.; López-Pérez, C.; Murenu, M.; Papiol, V.; Rumolo, P.; Scarcella, G. Spatial Variability in the Trophic Ecology and Biology of the Deep-Sea Shrimp Aristaeomorpha foliacea in the Mediterranean Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 2014, 87, 1–13. [Google Scholar] [CrossRef]
- Kapiris, K. Feeding Ecology of Parapenaeus longirostris (Lucas, 1846) (Decapoda: Penaeidae) from the Ionian Sea (Central and Eastern Mediterranean Sea). Sci. Mar. 2004, 68, 247–256. [Google Scholar] [CrossRef]
- Chartosia, N.; Tzomos, T.H.; Kitsos, M.S.; Karani, I.; Tselepides, A.; Koukouras, A. Karani Diet Comparison of the Bathyal Shrimps, Aristeus antennatus (Risso, 1816) and Aristaeomorpha foliacea (Risso, 1827) (Decapoda, Aristeidae) in the Eastern Mediterranean. Crustaceana 2005, 78, 273–284. [Google Scholar] [CrossRef]
- O’Reilly, C.M.; Hecky, R.E.; Cohen, A.S.; Plisnier, P.-D. Interpreting Stable Isotopes in Food Webs: Recognizing the Role of Time Averaging at Different Trophic Levels. Limnol. Oceanogr. 2002, 47, 306–309. [Google Scholar] [CrossRef]
- Peterson, B.J.; Fry, B. STABLE ISOTOPES IN ECOSYSTEM STUDIES. Annu. Rev. Ecol. Syst. 1987, 18, 293–320. [Google Scholar] [CrossRef]
- Schoeninger, M.J.; DeNiro, M.J. Nitrogen and Carbon Isotopic Composition of Bone Collagen from Marine and Terrestrial Animals. Geochim. Cosmochim. Acta 1984, 48, 625–639. [Google Scholar] [CrossRef]
- Vizzini, S.; Mazzola, A. Stable Isotopes and Trophic Positions of Littoral Fishes from a Mediterranean Marine Protected Area. Environ. Biol. Fish. 2009, 84, 13–25. [Google Scholar] [CrossRef]
- Fanelli, E.; Azzurro, E.; Bariche, M.; Cartes, J.E.; Maynou, F. Depicting the Novel Eastern Mediterranean Food Web: A Stable Isotopes Study Following Lessepsian Fish Invasion. Biol. Invasions 2015, 17, 2163–2178. [Google Scholar] [CrossRef]
- D’Iglio, C.; Famulari, S.; Albano, M.; Giordano, D.; Rinelli, P.; Capillo, G.; Spanò, N.; Savoca, S. Time-Scale Analysis of Prey Preferences and Ontogenetic Shift in the Diet of European Hake Merluccius merluccius (Linnaeus, 1758) in Southern and Central Tyrrhenian Sea. Fishes 2022, 7, 167. [Google Scholar] [CrossRef]
- Ricci, P.; Carlucci, R.; Capezzuto, F.; Carluccio, A.; Cipriano, G.; D’Onghia, G.; Maiorano, P.; Sion, L.; Tursi, A.; Libralato, S. Contribution of Intermediate and High Trophic Level Species to Benthic-Pelagic Coupling: Insights From Modelling Analysis. Front. Mar. Sci. 2022, 9, 887464. [Google Scholar] [CrossRef]
- Ricci, P.; Libralato, S.; Capezzuto, F.; D’Onghia, G.; Maiorano, P.; Sion, L.; Tursi, A.; Solidoro, C.; Carlucci, R. Ecosystem Functioning of Two Marine Food Webs in the North-Western Ionian Sea (Central Mediterranean Sea). Ecol. Evol. 2019, 9, 10198–10212. [Google Scholar] [CrossRef]
- Carbery, M.; O’Connor, W.; Palanisami, T. Trophic Transfer of Microplastics and Mixed Contaminants in the Marine Food Web and Implications for Human Health. Environ. Int. 2018, 115, 400–409. [Google Scholar] [CrossRef]
- Frias, J.P.G.L.; Otero, V.; Sobral, P. Evidence of Microplastics in Samples of Zooplankton from Portuguese Coastal Waters. Mar. Environ. Res. 2014, 95, 89–95. [Google Scholar] [CrossRef]
- Farrell, P.; Nelson, K. Trophic Level Transfer of Microplastic: Mytilus edulis (L.) to Carcinus maenas (L.). Environ. Pollut. 2013, 177, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Cau, A.; Avio, C.G.; Dessì, C.; Follesa, M.C.; Moccia, D.; Regoli, F.; Pusceddu, A. Microplastics in the Crustaceans Nephrops norvegicus and Aristeus antennatus: Flagship Species for Deep-Sea Environments? Environ. Pollut. 2019, 255, 113107. [Google Scholar] [CrossRef] [PubMed]
- Garcia, F.; De Carvalho, A.R.; Riem-Galliano, L.; Tudesque, L.; Albignac, M.; Ter Halle, A.; Cucherousset, J. Stable Isotope Insights into Microplastic Contamination within Freshwater Food Webs. Environ. Sci. Technol. 2021, 55, 1024–1035. [Google Scholar] [CrossRef] [PubMed]
- Falciai, L.; Minervini, R. Guida Dei Crostacei Decapodi d’Europa; Franco Muzzio: Padova, Italy, 1992; ISBN 978-88-7021-557-1. [Google Scholar]
- IAEA−600. Available online: https://analytical-reference-materials.iaea.org/iaea-600 (accessed on 23 November 2023).
- USGS40. Available online: https://analytical-reference-materials.iaea.org/usgs40 (accessed on 23 November 2023).
- IAEA-C-8. Available online: https://analytical-reference-materials.iaea.org/iaea-c-8 (accessed on 23 November 2023).
- IAEA-CH6. Available online: https://analytical-reference-materials.iaea.org/iaea-ch-6 (accessed on 23 November 2023).
- IAEA-N-1. Available online: https://analytical-reference-materials.iaea.org/iaea-n-1 (accessed on 23 November 2023).
- IAEA-N-2. Available online: https://analytical-reference-materials.iaea.org/iaea-n-2 (accessed on 23 November 2023).
- Post, D.M. Using Stable Isotopes To Estimate Trophic Position: Models, Methods, and Assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Skinner, M.M.; Martin, A.A.; Moore, B.C. Is Lipid Correction Necessary in the Stable Isotope Analysis of Fish Tissues? Rapid Commun. Mass Spectrom. 2016, 30, 881–889. [Google Scholar] [CrossRef] [PubMed]
- European Commission; Joint Research Centre; MSFD Technical Group on Marine Litter. Guidance on the Monitoring of Marine Litter in European Seas: An Update to Improve the Harmonised Monitoring of Marine Litter under the Marine Strategy Framework Directive; Publications Office: Luxembourg, 2023. [Google Scholar]
- Matiddi, M.; Pham, C.K.; Anastasopoulou, A.; Andresmaa, E.; Avio, C.G.; Bianchi, J.; Chiaeb, O.; Palazzo, L.; Darmon, G.; De Lucia, G.A.; et al. Monitoring Micro-Litter Ingestion in Marine Fish: A Harmonized Protocol for MSFD and RSCs Areas. INDICIT II Project 2021. Available online: https://accedacris.ulpgc.es/bitstream/10553/114417/1/Report_Monitoring-microlitter-ingestion-in-marine-fish.pdf (accessed on 23 November 2023).
- RStudio Team. RStudio: Integrated Development Environment for R. Version 4.3.2. 2023. Available online: https://www.rstudio.com/ (accessed on 23 November 2023).
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing Isotopic Niche Widths among and within Communities: SIBER—Stable Isotope Bayesian Ellipses in R: Bayesian Isotopic Niche Metrics. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Layman, C.A.; Araujo, M.S.; Boucek, R.; Hammerschlag-Peyer, C.M.; Harrison, E.; Jud, Z.R.; Matich, P.; Rosenblatt, A.E.; Vaudo, J.J.; Yeager, L.A.; et al. Applying Stable Isotopes to Examine Food-web Structure: An Overview of Analytical Tools. Biol. Rev. 2012, 87, 545–562. [Google Scholar] [CrossRef] [PubMed]
- Layman, C.A.; Arrington, D.A.; Montaña, C.G.; Post, D.M. CAN STABLE ISOTOPE RATIOS PROVIDE FOR COMMUNITY-WIDE MEASURES OF TROPHIC STRUCTURE? Ecology 2007, 88, 42–48. [Google Scholar] [CrossRef] [PubMed]
- McLeod, A.I. Kendall Rank Correlation and Mann-Kendall Trend Test. R Package Kendall 2005, 602, 1–10. [Google Scholar]
- Iitembu, J.A.; Dalu, T. Patterns of Trophic Resource Use among Deep-Sea Shrimps in the Northern Benguela Current Ecosystem, Namibia. Food Webs 2018, 16, e00089. [Google Scholar] [CrossRef]
- Leila, B.; Sedláček, P.; Anastasopoulou, A. Plastic Pollution in the Deep-Sea Giant Red Shrimp, Aristaeomorpha foliacea, in the Eastern Ionian Sea; an Alarm Point on Stock and Human Health Safety. Sci. Total Environ. 2023, 877, 162783. [Google Scholar] [CrossRef]
- D’Iglio, C.; Di Fresco, D.; Spanò, N.; Albano, M.; Panarello, G.; Laface, F.; Faggio, C.; Capillo, G.; Savoca, S. Occurrence of Anthropogenic Debris in Three Commercial Shrimp Species from South-Western Ionian Sea. Biology 2022, 11, 1616. [Google Scholar] [CrossRef] [PubMed]
- Bono, G.; Scannella, D.; Falsone, F.; Falco, F.; Maio, F.D.; Gabriele, M.; Gancitano, V.; Geraci, M.L.; Mancuso, M.; Okpala, C.; et al. Microplastics and Alien Black Particles as Contaminants of Deep-Water Rose Shrimp (Parapenaeus Longistroris Lucas, 1846) in the Central Mediterranean Sea. J. Adv. Biotechnol. Bioeng. 2020, 8, 23–28. [Google Scholar] [CrossRef]
- Yücel, N. Detection of Microplastic Fibers Tangle in Deep-Water Rose Shrimp (Parapenaeus longirostris, Lucas, 1846) in the Northeastern Mediterranean Sea. Environ. Sci. Pollut. Res. 2022, 30, 10914–10924. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Chen, B.; Xia, B.; Shi, X.; Qu, K. Polystyrene Microplastics Alter the Behavior, Energy Reserve and Nutritional Composition of Marine Jacopever (Sebastes schlegelii). J. Hazard. Mater. 2018, 360, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Grigorakis, S.; Mason, S.A.; Drouillard, K.G. Determination of the Gut Retention of Plastic Microbeads and Microfibers in Goldfish (Carassius auratus). Chemosphere 2017, 169, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Foekema, E.M.; De Gruijter, C.; Mergia, M.T.; Van Franeker, J.A.; Murk, A.J.; Koelmans, A.A. Plastic in North Sea Fish. Environ. Sci. Technol. 2013, 47, 8818–8824. [Google Scholar] [CrossRef] [PubMed]
- Neves, D.; Sobral, P.; Ferreira, J.L.; Pereira, T. Ingestion of Microplastics by Commercial Fish off the Portuguese Coast. Mar. Pollut. Bull. 2015, 101, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Valente, T.; Pelamatti, T.; Avio, C.G.; Camedda, A.; Costantini, M.L.; De Lucia, G.A.; Jacomini, C.; Piermarini, R.; Regoli, F.; Sbrana, A.; et al. One Is Not Enough: Monitoring Microplastic Ingestion by Fish Needs a Multispecies Approach. Mar. Pollut. Bull. 2022, 184, 114133. [Google Scholar] [CrossRef]
- Carreras-Colom, E.; Constenla, M.; Soler-Membrives, A.; Cartes, J.E.; Baeza, M.; Padrós, F.; Carrassón, M. Spatial Occurrence and Effects of Microplastic Ingestion on the Deep-Water Shrimp Aristeus antennatus. Mar. Pollut. Bull. 2018, 133, 44–52. [Google Scholar] [CrossRef]
- Chubarenko, I.; Bagaev, A.; Zobkov, M.; Esiukova, E. On Some Physical and Dynamical Properties of Microplastic Particles in Marine Environment. Mar. Pollut. Bull. 2016, 108, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Ruz, V.; Gutow, L.; Thompson, R.C.; Thiel, M. Microplastics in the Marine Environment: A Review of the Methods Used for Identification and Quantification. Environ. Sci. Technol. 2012, 46, 3060–3075. [Google Scholar] [CrossRef] [PubMed]
- Van Cauwenberghe, L.; Devriese, L.; Galgani, F.; Robbens, J.; Janssen, C.R. Microplastics in Sediments: A Review of Techniques, Occurrence and Effects. Mar. Environ. Res. 2015, 111, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Erni-Cassola, G.; Zadjelovic, V.; Gibson, M.I.; Christie-Oleza, J.A. Distribution of Plastic Polymer Types in the Marine Environment; A Meta-Analysis. J. Hazard. Mater. 2019, 369, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Lopes, C.; Raimundo, J.; Caetano, M.; Garrido, S. Microplastic Ingestion and Diet Composition of Planktivorous Fish. Limnol. Ocean. Lett. 2020, 5, 103–112. [Google Scholar] [CrossRef]
- Ory, N.C.; Sobral, P.; Ferreira, J.L.; Thiel, M. Amberstripe Scad Decapterus muroadsi (Carangidae) Fish Ingest Blue Microplastics Resembling Their Copepod Prey along the Coast of Rapa Nui (Easter Island) in the South Pacific Subtropical Gyre. Sci. Total Environ. 2017, 586, 430–437. [Google Scholar] [CrossRef]
- Murray, F.; Cowie, P.R. Plastic Contamination in the Decapod Crustacean Nephrops norvegicus (Linnaeus, 1758). Mar. Pollut. Bull. 2011, 62, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Browne, M.A.; Dissanayake, A.; Galloway, T.S.; Lowe, D.M.; Thompson, R.C. Ingested Microscopic Plastic Translocates to the Circulatory System of the Mussel, Mytilus edulis (L.). Environ. Sci. Technol. 2008, 42, 5026–5031. [Google Scholar] [CrossRef] [PubMed]
- Denton, E.J. Light and Life in the Sea; Cambridge University Press: Cambridge, UK, 1990; ISBN 0-521-39207-1. [Google Scholar]
- Bauer, R.T. Shrimps: Their Diversity, Intriguing Adaptations and Varied Lifestyles; Fish & Fisheries Series; Springer International Publishing: Cham, Switzerland, 2023; Volume 42, ISBN 978-3-031-20965-9. [Google Scholar]
- Deopura, B.L.; Alagirusamy, R.; Textile Institute (Eds.) Polyesters and Polyamides; Woodhead Publishing in Textiles; CRC Press: Boca Raton, FL, USA, 2008; ISBN 978-1-84569-298-8. [Google Scholar]
- Turner, A. Paint Particles in the Marine Environment: An Overlooked Component of Microplastics. Water Res. X 2021, 12, 100110. [Google Scholar] [CrossRef]
- Lithner, D.; Larsson, Å.; Dave, G. Environmental and Health Hazard Ranking and Assessment of Plastic Polymers Based on Chemical Composition. Sci. Total Environ. 2011, 409, 3309–3324. [Google Scholar] [CrossRef]
- Berto, D.; Rampazzo, F.; Gion, C.; Noventa, S.; Ronchi, F.; Traldi, U.; Giorgi, G.; Cicero, A.M.; Giovanardi, O. Preliminary Study to Characterize Plastic Polymers Using Elemental Analyser/Isotope Ratio Mass Spectrometry (EA/IRMS). Chemosphere 2017, 176, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Shum, P.; Wäge-Recchioni, J.; Sellers, G.S.; Johnson, M.L.; Joyce, D.A. DNA Metabarcoding Reveals the Dietary Profiles of a Benthic Marine Crustacean, Nephrops Norvegicus. PLoS ONE 2023, 18, e0289221. [Google Scholar] [CrossRef] [PubMed]
- Mackas, D.; Bohrer, R. Fluorescence Analysis of Zooplankton Gut Contents and an Investigation of Diel Feeding Patterns. J. Exp. Mar. Biol. Ecol. 1976, 25, 77–85. [Google Scholar] [CrossRef]
- Kousteni, V.; Karachle, P.K.; Megalofonou, P. Diet of the Small-Spotted Catshark Scyliorhinus canicula in the Aegean Sea (Eastern Mediterranean). Mar. Biol. Res. 2017, 13, 161–173. [Google Scholar] [CrossRef]
- Fanelli, E.; Rey, J.; Torres, P.; Gil De Sola, L. Feeding Habits of Blackmouth Catshark Galeus melastomus Rafinesque, 1810 and Velvet Belly Lantern Shark Etmopterus spinax (Linnaeus, 1758) in the Western Mediterranean. J. Appl. Ichthyol. 2009, 25, 83–93. [Google Scholar] [CrossRef]
- Anastasopoulou, A.; Mytilineou, C.; Lefkaditou, E.; Dokos, J.; Smith, C.J.; Siapatis, A.; Bekas, P.; Papadopoulou, K.-N. Diet and Feeding Strategy of Blackmouth Catshark Galeus melastomus. J. Fish Biol. 2013, 83, 1637–1655. [Google Scholar] [CrossRef]
- Esposito, V.; Andaloro, F.; Bianca, D.; Natalotto, A.; Romeo, T.; Scotti, G.; Castriota, L. Diet and Prey Selectivity of the Red Mullet, Mullus barbatus (Pisces: Mullidae), from the Southern Tyrrhenian Sea: The Role of the Surf Zone as a Feeding Ground. Mar. Biol. Res. 2014, 10, 167–178. [Google Scholar] [CrossRef]
- Zicarelli, G.; Romano, C.; Gallo, S.; Valentino, C.; Pepe Bellomo, V.; Leonetti, F.L.; Giglio, G.; Neri, A.; Marsili, L.; Milazzo, C.; et al. Diet and Plastic Ingestion in the Blackmouth Catshark Galeus Melastomus, Rafinesque 1810, in Italian Waters. Animals 2023, 13, 1039. [Google Scholar] [CrossRef] [PubMed]
- Carpentieri, P.; Colloca, F.; Cardinale, M.; Belluscio, A.; Ardizzone, G. Feeding Habits of European Hake (Merluccius merluccius) in the Central Mediterranean Sea. Fish. Bull. 2005, 103, 411–416. [Google Scholar]
- Bello, G. The Feeding Ecology of the Velvet Belly, Etmopterus spinax (Chondrichthyes: Squalidae), of the Adriatic; Società Italiana Scienze Naturali: Milano, Italy, 1998. [Google Scholar]
- Valente, T.; Sbrana, A.; Scacco, U.; Jacomini, C.; Bianchi, J.; Palazzo, L.; De Lucia, G.A.; Silvestri, C.; Matiddi, M. Exploring Microplastic Ingestion by Three Deep-Water Elasmobranch Species: A Case Study from the Tyrrhenian Sea. Environ. Pollut. 2019, 253, 342–350. [Google Scholar] [CrossRef]
- Wang, W.; Ge, J.; Yu, X. Bioavailability and Toxicity of Microplastics to Fish Species: A Review. Ecotoxicol. Environ. Saf. 2020, 189, 109913. [Google Scholar] [CrossRef] [PubMed]
- Alomar, C.; Deudero, S. Evidence of Microplastic Ingestion in the Shark Galeus Melastomus Rafinesque, 1810 in the Continental Shelf off the Western Mediterranean Sea. Environ. Pollut. 2017, 223, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Giani, D.; Baini, M.; Galli, M.; Casini, S.; Fossi, M.C. Microplastics Occurrence in Edible Fish Species (Mullus barbatus and Merluccius merluccius) Collected in Three Different Geographical Sub-Areas of the Mediterranean Sea. Mar. Pollut. Bull. 2019, 140, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.V.B.; Justino, A.K.S.; Eduardo, L.N.; Schmidt, N.; Martins, J.R.; Ménard, F.; Fauvelle, V.; Mincarone, M.M.; Lucena-Frédou, F. Influencing Factors for Microplastic Intake in Abundant Deep-Sea Lanternfishes (Myctophidae). Sci. Total Environ. 2023, 867, 161478. [Google Scholar] [CrossRef]
- Vecchi, S.; Bianchi, J.; Scalici, M.; Fabroni, F.; Tomassetti, P. Field Evidence for Microplastic Interactions in Marine Benthic Invertebrates. Sci. Rep. 2021, 11, 20900. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | δ15N | δ13C | NR | CR | TA | CD | SEAc |
|---|---|---|---|---|---|---|---|
| P. longirostris | 7.7 ± 0.07 | −18.4 ± 0.04 | 1.624 | 1.002 | 0.502 | 0.275 | 0.168 |
| A. foliacea | 7.8 ± 0.06 | −18.1 ± 0.05 | 1.046 | 1.014 | 0.543 | 0.339 | 0.199 |
| Species | tW (g) | cL (cm) | Sex (F%) | FI μ (%) | HSI (%) | Items/Ind. |
|---|---|---|---|---|---|---|
| P. longirostris | 11.1 ± 0.44 | 2.9 ± 0.04 | 80% | 1.9% | 2.9% | 1.44 ± 0.62 |
| A. foliacea | 13.1 ± 0.36 | 3.3 ± 0.03 | 56% | 4.1% | 6.5% | 2.56 ± 0.73 |
| All individuals | 12.1 ± 0.3 | 3.1 ± 0.03 | 68% | 2.9% | 4.7% | 2.04 ± 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciaralli, L.; Valente, T.; Monfardini, E.; Libralato, G.; Manfra, L.; Berto, D.; Rampazzo, F.; Gioacchini, G.; Chemello, G.; Piermarini, R.; et al. Rose or Red, but Still under Threat: Comparing Microplastics Ingestion between Two Sympatric Marine Crustacean Species (Aristaeomorpha foliacea and Parapenaeus longirostris). Animals 2024, 14, 2212. https://doi.org/10.3390/ani14152212
Ciaralli L, Valente T, Monfardini E, Libralato G, Manfra L, Berto D, Rampazzo F, Gioacchini G, Chemello G, Piermarini R, et al. Rose or Red, but Still under Threat: Comparing Microplastics Ingestion between Two Sympatric Marine Crustacean Species (Aristaeomorpha foliacea and Parapenaeus longirostris). Animals. 2024; 14(15):2212. https://doi.org/10.3390/ani14152212
Chicago/Turabian StyleCiaralli, Laura, Tommaso Valente, Eleonora Monfardini, Giovanni Libralato, Loredana Manfra, Daniela Berto, Federico Rampazzo, Giorgia Gioacchini, Giulia Chemello, Raffaella Piermarini, and et al. 2024. "Rose or Red, but Still under Threat: Comparing Microplastics Ingestion between Two Sympatric Marine Crustacean Species (Aristaeomorpha foliacea and Parapenaeus longirostris)" Animals 14, no. 15: 2212. https://doi.org/10.3390/ani14152212