
Quill Mites of the Subfamily Syringophilinae (Acariformes: Syringophilidae) Parasitising Starlings (Passeriformes: Sturnidae) †

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
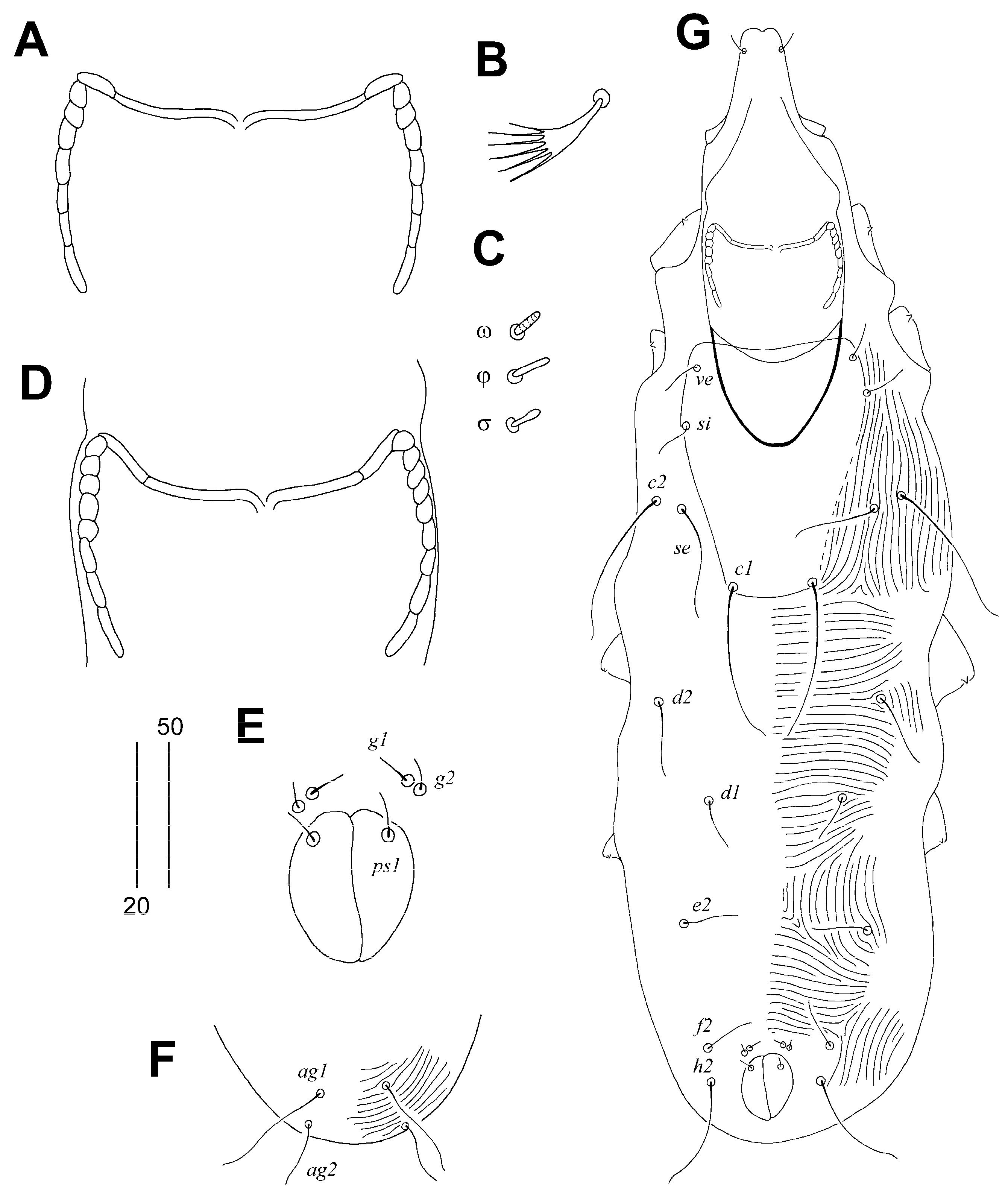
3.1. Descriptions
3.1.1. Aulonastus indonesianus Marcisova, Skoracki and Patan sp. n.
Type Material
Type Material Deposition
Additional Material
Differential Diagnosis
Etymology
3.1.2. Aulonastus anais Skoracki and Patan sp. n.
Type Material
Type Material Deposition
Differential Diagnosis
Etymology
3.1.3. Syringophiloidus poeopterus Skoracki and Patan sp. n.
Type Material
Types Deposition
Differential Diagnosis
Etymology
3.1.4. Syringophiloidus presentalis Chirov and Kravtsova, 1995
3.1.5. Syringophiloidus saponai Skoracki, Patan and Unsoeld, 2024
New Material Examined
3.1.6. Syringophiloidus graculae Fain, Bochkov and Mironov, 2000
New Material Examined
3.1.7. Syringophilopsis sturni Chirov and Kravtsova, 1995
3.1.8. Syringophilopsis parasturni Skoracki, Patan and Unsoeld, 2024
New Material Examined
3.1.9. Krantziaulonastus buczekae (Skoracki, 2002)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lavoipierre, M.M.J. The undescribed male and female of the Pigeon quill mite, Syringophilus columbae Hirst, 1920. Trans. R. Soc. Trop. Med. Hyg. 1953, 47, 7. [Google Scholar]
- Kethley, J.B. A revision of the family Syringophilidae (Prostigmata: Acarina). Contrib. Am. Entomol. Inst. 1970, 5, 1–76. [Google Scholar]
- Kethley, J.B. Population regulation in quill mites (Acari: Syringophilidae). Ecology 1971, 52, 1113–1118. [Google Scholar] [CrossRef]
- Skoracki, M. Quill mites (Acari: Syringophilidae) of the Palaearctic region. Zootaxa 2011, 2840, 1–414. [Google Scholar] [CrossRef]
- Skoracki, M.; Sikora, B.; Spicer, G.S. A review of the subfamily Picobiinae Johnston and Kethley, 1973 (Acariformes: Prostigmata: Syringophilidae). Zootaxa 2016, 4113, 1–95. [Google Scholar] [CrossRef] [PubMed]
- Hromada, M.; Klimovicova, M.; Unsöld, M.; Skoracki, M. Host-parasite relationships in the system composed by cuckoos and quill mites. Syst. Appl. Acarol. 2016, 21, 528–536. [Google Scholar] [CrossRef]
- Sikora, B.; Unsoeld, M.; Melzer, R.R.; Friedrich, S.; Skoracki, M. First Records of Picobiine Mites Associated with Birds-of-Paradise: Can Interspecific Sexual Behaviour of Hosts Play a Role in the Distribution of Quill Mite Parasites? Animals 2023, 13, 1509. [Google Scholar] [CrossRef] [PubMed]
- Skoracki, M.; Unsoeld, M.; Kosicki, J.Z.; Melzer, R.R.; Friedrich, S.; Sikora, B. Enigmatic host-mite relationships: Unraveling the distribution of quill mites on Birds-of-Paradise. Int. J. Parasitol. 2024, 54, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Nattress, B. Horizontal transmission of Syringophilopsis kirgizorum (Acari: Cheyletoidea: Syringophilidae). Acarina 2011, 19, 270. [Google Scholar]
- Casto, S.D. Dispersal of the quill mite Syringophiloidus minor (Berlese) (Acarina: Syringophilidae). J. Med. Entomol. 1976, 13, 357–360. [Google Scholar]
- Skirnisson, K.; Nielsen, Ó.K. Quill mite infestation of rock ptarmigan Lagopus muta (Aves: Phasianidae) in relation to year and host age, sex, body condition, and density. Parasitol. Res. 2019, 118, 2643–2650. [Google Scholar] [CrossRef] [PubMed]
- Skoracki, M.; Hromada, M.; Sikora, B. Castosyringophilus meropis sp. n. (Acariformes: Syringophilidae)—A new quill mite species parasitising the world population of Merops apiaster Linnaeus (Coraciiformes: Meropidae). Folia Parasitol. 2017, 64, 24. [Google Scholar] [CrossRef] [PubMed]
- Grossi, A.; Proctor, H. The distribution of quill mites (Betasyringophiloidus seiuri) among flight feathers of the ovenbird (Seiurus aurocapilla). J. Parasitol. 2020, 106, 82–89. [Google Scholar] [CrossRef]
- Skoracki, M.; Michalik, J.; Sikora, B. Prevalence and habitat preference of quill mites (Acari, Syringophilidae) parasitising forest passerine birds in Poland. Acta Parasitol. 2010, 55, 188–193. [Google Scholar] [CrossRef]
- Szymański, P.; Niśkiewicz, M.; Budka, M.; Zampa, L.; Osiejuk, T.S.; Skoracki, M. Quill mites of the family Syringophilidae (Acariformes: Prostigmata) parasitising (Columbiformes: Columbidae) doves of the genus Turtur. Syst. Appl. Acarol. 2023, 28, 1466–1475. [Google Scholar] [CrossRef]
- Rebrassier, R.E.; Martin, E.D. Syringophilus bipectinatus, a quill mite of poultry. Science 1932, 76, 128. [Google Scholar] [CrossRef]
- Schwabe, O. A quill mite of poultry—A case report. J. Am. Vet. Med. Assoc. 1956, 129, 481–482. [Google Scholar] [PubMed]
- Hwang, J.C. Case reports of the quill mite, Syringophilus bipectinatus in poultry. Proc. Helminthol. Soc. Wash. 1959, 26, 47–50. [Google Scholar]
- Gritsenko, E.F. The biology and ecology of the quill mite Syringophilus bipectinatus Heller, 1880. In Proceedings of the 3rd International Congress of Acarology, Prague, Czechoslovakia, 31 August–6 September 1971; Milan, D., Rosicky, B., Eds.; Springer: Dordrecht, The Netherlands, 1971; pp. 515–516. [Google Scholar]
- Skoracki, M.; Michalik, J.; Skotarczak, B.; Rymaszewska, A.; Sikora, B.; Hofman, T.; Wodecka, B.; Sawczuk, M. First detection of Anaplasma phagocytophilum in quill mites (Acari: Syringophilidae) parasitising passerine birds in Poland. Microbes Infect. 2006, 8, 303–307. [Google Scholar] [CrossRef]
- Glowska, E.; Dragun-Damian, A.; Dabert, M.; Gerth, M. New Wolbachia supergroups detected in quill mites (Acari: Syringophilidae). Infect. Genet. Evol. 2015, 30, 140–146. [Google Scholar] [CrossRef]
- Glowska, E.; Filutowska, Z.K.; Dabert, M.; Gerth, M. Microbial composition of enigmatic bird parasites: Wolbachia and Spiroplasma are the most important bacterial associates of quill mites (Acariformes: Syringophilidae). Microbiologyopen 2019, 9, e964. [Google Scholar] [CrossRef] [PubMed]
- Johnston, D.E.; Kethley, J.B. A numerical phenetic study of the quill mites of the family Syringophilidae (Acari). J. Parasitol. 1973, 59, 520–530. [Google Scholar] [CrossRef]
- Glowska, E.; Chrzanowski, M.; Kaszewska, K. Checklist of the quill mites (Acariformes: Syringophilidae) of the World. Zootaxa 2015, 3968, 1–81. [Google Scholar] [CrossRef] [PubMed]
- Zmudzinski, M.; Skoracki, M.; Sikora, B. An Updated Checklist of Quill Mites of the Family Syringophilidae (Acariformes: Prostigmata). 2023. Available online: https://figshare.com/articles/dataset/An_updated_checklist_of_quill_mites_of_the_family_Syringophilidae_Acariformes_Prostigmata_/16529574 (accessed on 27 July 2024).
- Skoracki, M.; Fajfer, M.; Hromada, M.; Hušek, J.; Sikora, B. Tinamiphilopsis temmincki sp. n., a New Quill Mite Species from Tataupa Tinamou, and the Early History of Syringophilid Mites. Animals 2023, 13, 2728. [Google Scholar] [CrossRef] [PubMed]
- Krantz, G.W.; Walter, D.E. A Manual of Acarology, 3rd ed.; Texas Tech University Press: Lubbock, TX, USA, 2009. [Google Scholar]
- Grandjean, F. Les segments post-larvaires de l’hystérosoma chez les Oribates (Acariens). Bull. Soc. Zool. Fr. 1939, 64, 273–284. [Google Scholar]
- Kethley, J.B. Acarina: Prostigmata (Actinedida). In Soil Biology Guide; Dindal, D.L., Ed.; John Wiley & Sons: New York, NY, USA, 1990; pp. 667–756. [Google Scholar]
- Grandjean, F. Observations Sur Les Acariens de La Famille Des Stigmaeidae. Arch. Sci. Phys. Nat. 1944, 26, 103–131. [Google Scholar]
- Clements, J.F.; Rasmussen, P.C.; Schulenberg, T.S.; Iliff, M.J.; Fredericks, T.A.; Gerbracht, J.A.; Lepage, D.; Spencer, A.; Billerman, S.M.; Sullivan, B.L.; et al. The eBird/Clements Checklist of Birds of the World. 2023. Available online: https://www.birds.cornell.edu/clementschecklist/download/ (accessed on 27 July 2024).
- del Hoyo, J.; Collar, N.J.; Christie, D.A.; Elliott, A.; Fishpool, L.D.C.; Boesman, P.; Kirwan, G.M. HBW and BirdLife International Illustrated Checklist of the Birds of the World. Volume 2: Passerines; Lynx Edicions and BirdLife International: Barcelona, Spain; Cambridge, UK, 2016. [Google Scholar]
- Holt, B.G.; Lessard, J.P.; Borregaard, M.K.; Fritz, S.A.; Araújo, M.B.; Dimitrov, D.; Fabre, P.H.; Graham, C.H.; Graves, G.R.; Jønsson, K.A.; et al. An update of Wallace’s zoogeographic regions of the world. Science 2013, 339, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Ficetola, G.; Mazel, F.; Thuiller, W. Global determinants of zoogeographical boundaries. Nat. Ecol. Evol. 2017, 1, 0089. [Google Scholar] [CrossRef] [PubMed]
- Skoracki, M.; Sikora, B.; Unsoeld, M.; Hromada, M. Mite Fauna of the Family Syringophilidae (Acariformes: Prostigmata) Parasitizing Darwin’s Finches in Galápagos Archipelago. Diversity 2022, 14, 585. [Google Scholar] [CrossRef]
- Skoracki, M.; Patan, M.; Unsoeld, M.; Hromada, M.; Kwieciński, Z.; Marcisova, I. Diversity of Quill Mites of the Family Syringophilidae (Acariformes: Prostigmata) Parasitizing Starlings of the Genus Lamprotornis (Passeriformes: Sturnidae). Diversity 2024, 16, 51. [Google Scholar] [CrossRef]
- Chirov, P.A.; Kravtsova, N.T. A new genus and new species of mites of the family Syringophilidae. Parazitologiya 1995, 29, 370–379. [Google Scholar]
- Fain, A.; Bochkov, A.V.; Mironov, S.V. New genera and species of quill mites of the family Syringophilidae (Acari: Prostigmata). Bull. Inst. R. Sci. Nat. Belg. 2000, 70, 33–70. [Google Scholar]
- Bochkov, A.V.; Mironov, S.V. Quill mites of the family Syringophilidae Lavoipierre, 1953 (Acariformes: Prostigmata) parasitic on birds (Aves) of the fauna of the former USSR. Acarina 1998, 6, 3–16. [Google Scholar]
- Skoracki, M. Quill mites of the genus Syringophilopsis (Acari, Syringophilidae) from passeriform birds of Poland with descriptions of five new species. Acta Parasitol. 2004, 49, 45–62. [Google Scholar]
- Skoracki, M. Three new species of quill mites of the genus Aulonastus Kethley, 1970 (Acari, Prostigmata, Syringophilidae) from passerine birds. Acta Parasitol. 2002, 47, 300–305. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Starlings (Sturnidae), version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Fjeldså, J.; Christidis, L.; Ericson, P.G.P. The Largest Avian Radiation: The Evolution of Perching Birds, or the Order Passeriformes; Lynx Edicions: Barcelona, Spain, 2020. [Google Scholar]
- Skoracki, M.; Spicer, G.S.; Oconnor, B.M. A systematic review of the subfamily Syringophilinae (Acari: Syringophilidae) of the Nearctic region. Part 1: Quill mites associated with passerines (Aves: Passeriformes). Zootaxa 2016, 4084, 451–494. [Google Scholar] [CrossRef] [PubMed]
- Glowska, E.; Laniecka, I.; Ostrowska, K.; Gebhard, C.A.; Olechnowicz, J.; Dabert, M. Evaluation of Genetic Diversity in Quill Mites of the Genus Syringophiloidus Kethley, 1970 (Prostigmata: Syringophilidae) with Six New-to-Science Species. Animals 2023, 13, 3877. [Google Scholar] [CrossRef]
- Sibley, C.G.; Monroe, B.L. Distribution and Taxonomy of Birds of the World; Yale University Press: New Haven, CT, USA, 1990. [Google Scholar]
- Zuccon, D.; Cibois, A.; Pasquet, E.; Ericson, P.G.P. Nuclear and mitochondrial sequence data reveal the major lineages of starlings, mynas and related taxa. Mol. Phylogenet. Evol. 2006, 41, 333–344. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mite Species | Host Species | Locality | References |
|---|---|---|---|
| Aulonastus indonesianus Marcisova, Skoracki and Patan sp. n. | Common Hill Myna, Gracula religiosa Linnaeus | OR: Indonesia (Java) | Current study |
| “ | White-necked Myna, Streptocitta albicollis (Vieillot) | OR: Indonesia (Celebes) | Current study |
| Aulonastus anais Skoracki and Patan sp. n. | Golden Myna, Mino anais (Lesson) | PM: Papua New Guinea | Current study |
| Krantziaulonastus buczekae Skoracki, 2002 | Common Starling, Sturnus vulgaris Linnaeus | PA: Poland | [4,41] |
| Syringophiloidus poeopterus Skoracki and Patan sp. n. | Abbott’s Starling, Poeoptera femoralis (Richmond) | ET: Tanzania | Current study |
| Syringophiloidus presentalis Chirov and Kravtsova, 1995 | Common Starling, Sturnus vulgaris Linnaeus | PA: Poland, Slovakia, France, Kyrgyzstan | [4,37] |
| Syringophiloidus saponai Skoracki, Patan and Unsoeld, 2024 | Greater Blue-eared Glossy-Starling, Lamprotornis chalybaeus Hemprich and Ehrenberg | ET: Kenya, Tanzania, Ethiopia | [36] |
| “ | Superb Starling, Lamprotornis superbus Rüppell | ET: Tanzania, Kenya | [36] |
| “ | Lesser Blue-eared Glossy-Starling, Lamprotornis chloropterus Swainson | ET: Tanzania | [36] |
| “ | Ashy Starling, Lamprotornis unicolor (Shelley) | ET: Tanzania | [36] |
| “ | Pale-winged Starling, Onychognathus nabouroup (Daudin) | ET: Namibia | Current study |
| “ | Red-winged Starling, Onychognathus morio (Linnaeus) | ET: Tanzania | Current study |
| Syringophiloidus graculae Fain, Bochkov and Mironov, 2000 | Common Hill Myna, Gracula religiosa Linnaeus | OR: SE Asia | [38] |
| “ | Golden-crested Myna, Ampeliceps coronatus Blyth | OR: Indochina | Current study |
| “ | Coleto, Sarcops calvus (Linnaeus) | OR: Philippines | Current study |
| Syringophilopsis sturni Chirov and Kravtsova, 1995 | Common Starling, Sturnus vulgaris Linnaeus | PA: Kyrgyzstan, Kazakhstan, Poland, Ukraine | [4,37,39,40] |
| Syringophilopsis parasturni Skoracki, Patan and Unsoeld, 2024 | Chestnut-bellied Starling, Lamprotornis pulcher (Müller) | ET: Senegal | [36] |
| “ | Bronze-tailed Glossy-Starling, Lamprotornis chalcurus Nordmann | ET: Senegal | [36] |
| “ | Black-bellied Glossy-Starling, Notopholia corusca (Nordmann) | ET: Tanzania | current study |
| “ | Abbott’s Starling, Poeoptera femoralis (Richmond) | ET: Tanzania | current study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcisova, I.; Skoracki, M.; Patan, M.; Hromada, M.; Sikora, B. Quill Mites of the Subfamily Syringophilinae (Acariformes: Syringophilidae) Parasitising Starlings (Passeriformes: Sturnidae). Animals 2024, 14, 2239. https://doi.org/10.3390/ani14152239
Marcisova I, Skoracki M, Patan M, Hromada M, Sikora B. Quill Mites of the Subfamily Syringophilinae (Acariformes: Syringophilidae) Parasitising Starlings (Passeriformes: Sturnidae). Animals. 2024; 14(15):2239. https://doi.org/10.3390/ani14152239
Chicago/Turabian StyleMarcisova, Iva, Maciej Skoracki, Milena Patan, Martin Hromada, and Bozena Sikora. 2024. "Quill Mites of the Subfamily Syringophilinae (Acariformes: Syringophilidae) Parasitising Starlings (Passeriformes: Sturnidae)" Animals 14, no. 15: 2239. https://doi.org/10.3390/ani14152239
APA StyleMarcisova, I., Skoracki, M., Patan, M., Hromada, M., & Sikora, B. (2024). Quill Mites of the Subfamily Syringophilinae (Acariformes: Syringophilidae) Parasitising Starlings (Passeriformes: Sturnidae). Animals, 14(15), 2239. https://doi.org/10.3390/ani14152239