
Zoonotic Tick-Borne Pathogens in Ticks from Vegetation and Alpine Ibex (Capra ibex) in the Maritime Alps, Italy
 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Tick Collection
2.3. Laboratory Analyses
2.4. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garcia-Vozmediano, A.; Krawczyk, A.I.; Sprong, H.; Rossi, L.; Ramassa, E.; Tomassone, L. Ticks climb the mountains: Ixodid tick infestation and infection by tick-borne pathogens in Western Alps. Ticks Tick Borne Dis. 2020, 11, 101489. [Google Scholar] [CrossRef] [PubMed]
- Tomassone, L.; Berriatua, E.; de Sousa, R.; Duscher, G.G.; Mihalca, A.; Sprong, H.; Silaghi, C.; Zintl, A. Neglected vector-borne zoonoses in Europe: Into the wild. Vet. Parasitol. 2018, 251, 17–26. [Google Scholar] [CrossRef]
- Peralbo-Moreno, A.; Baz-Flores, S.; Cuadrado-Matías, R.; Barroso, P.; Triguero-Ocaña, R.; Jiménez-Ruiz, S.; Herraiz, C.; Ruiz-Rodríguez, C.; Acevedo, P.; Ruiz-Fons, F. Environmental factors driving fine-scale ixodid tick abundance patterns. Sci. Total Environ. 2022, 20, 158633. [Google Scholar] [CrossRef] [PubMed]
- Mustoni, A.; Pedrotti, L.; Zanon, E.; Tosi, G. Gli Ungulati Delle Alpi: Biologia, Riconoscimento E Gestione; Immagine, N., Ed.; Cles: Trento, Italy, 2002; p. 538. [Google Scholar]
- Messner, M.; Kayikci, F.N.; Shahi-Barogh, B.; Harl, J.; Messner, C.; Fuehrer, H.P. Screening of wild ruminants from the Kaunertal and other alpine regions of Tyrol (Austria) for vector-borne pathogens. Parasitol. Res. 2019, 118, 2735–2740. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L.; Meneguz, P.G. Fauna ixodologica in ruminanti selvatici del Piemonte. Ann. Fac. Med. Vet. Torino 1989, 33, 335–345. [Google Scholar]
- ARPA Piemonte. Assessment Climatico Della Provincia di Cuneo—CclimaTT, Cambiamenti Climatici nel Territorio Transfrontaliero”, programma Interreg VA Italia-Francia Alcotra 2014–2020. 2019. Available online: http://www.cclimatt.eu/fileadmin/cclimatt/contents/file/LOW_relazione_provinciaCN_assessment.pdf (accessed on 2 July 2024).
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2014: Synthesis Report. In Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2014. [Google Scholar]
- Comino, E.; Fiorucci, A.; Rosso, M.; Terenziani, A.; Treves, A. Vegetation and Glacier Trends in the area of the Maritime Alps Natural Park (Italy): MaxEnt application to predict habitat development. Climate 2021, 9, 54. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2018. Available online: http://qgis.osgeo.org (accessed on 2 July 2024).
- Moine, R. ProjectAlcotra LEMED-IBEX, Monitoring and Management of Capra ibex from lake Leman (Geneva) to Mediterranean Sea. In Progetto ALCOTRA 1664—LEMED-IBEX “Monitoraggio e gestione dello Stambecco dal lago di Gienvra (Léman) al Mediterraneo”—Conferenza Finale—Abstract Book, 2020, 3. Available online: https://it.marittimemercantour.eu/media/14cbfc3e.pdf (accessed on 26 July 2024).
- Cringoli, G.; Iori, A.; Rinaldi, L.; Veneziano, V.; Genchi, C. Zecche. In Mappe Parassitologiche; Ronaldo Editore: Napoli, Italy, 2005; p. 263. [Google Scholar]
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Ticks of Europe and North Africa: A Guide to Species Identification; Springer Nature: Cham, Switzerland, 2017; p. 404. [Google Scholar]
- Rijpkema, S.G.; Molkenboer, M.J.; Schouls, L.M.; Jongejan, F.; Schellekens, J.F. Simultaneous detection and genotyping of three genomic groups of Borrelia burgdorferi sensu lato in Dutch Ixodes ricinus ticks by characterization of the amplified intergenic spacer region between 5S and 23S rRNA genes. J. Clin. Microbiol. 1995, 3, 3091–3095. [Google Scholar] [CrossRef] [PubMed]
- Labruna, M.B.; Whitworth, T.; Bouyer, D.H.; Mcbride, J.W.; Pinter, A.; Popov, V.; Gennari, S.M.; Walker, D.H. Rickettsia species infecting Amblyomma cooperi ticks from an area in the state of Sao Paulo, Brazil, where Brazilian Spotted Fever is endemic. J. Clin. Microbiol. 2004, 42, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B.D. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J. Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef]
- Jahfari, S.; Coipan, E.C.; Fonville, M.; van Leeuwen, A.D.; Hengeveld, P.; Heylen, D.; Heyman, P.; van Maanen, C.; Butler, C.M.; Földvári, G.; et al. Circulation of four Anaplasma phagocytophilum ecotypes in Europe. Parasit. Vectors 2014, 7, 365. [Google Scholar] [CrossRef]
- Alberti, A.; Zobba, R.; Chessa, B.; Addis, M.F.; Sparagano, O.; Pinna Parpaglia, M.L.; Cubeddu, T.; Pintori, G.; Pittau, M. Equine and canine Anaplasma phagocytophilum strains isolated on the island of Sardinia (Italy) are phylogenetically related to pathogenic strains from the United States. Appl. Environ. Microbiol. 2005, 71, 6418–6422. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Jaarsma, R.I.; Sprong, H.; Takumi, K.; Kazimirova, M.; Silaghi, C.; Mysterud, A.; Rudolf, I.; Beck, R.; Földvári, G.; Tomassone, L.; et al. Anaplasma phagocytophilum evolves in geographical and biotic niches of vertebrates and ticks. Parasit Vectors. 2019, 12, 328. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.; Posada, D.; Crandall, K. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1660. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: http://www.r-project.org/index.html (accessed on 2 July 2024).
- Jaenson, T.G.; Jaenson, D.G.; Eisen, L.; Petersson, E.; Lindgren, E. Changes in the geographical distribution and abundance of the tick Ixodes ricinus during the past 30 years in Sweden. Parasit Vectors 2012, 10, 8. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasit Vectors 2013, 6, 1. [Google Scholar] [CrossRef]
- Földvári, G.; Široký, P.; Szekeres, S.; Majoros, G.; Sprong, H. Dermacentor reticulatus: A vector on the rise. Parasit Vectors 2016, 9, 314. [Google Scholar] [CrossRef]
- Uiterwijk, M.; Ibáñez-Justicia, A.; van de Vossenberg, B.; Jacobs, F.; Overgaauw, P.; Nijsse, R.; Dabekaussen, C.; Stroo, A.; Sprong, H. Imported Hyalomma ticks in the Netherlands 2018–2020. Parasit Vectors 2021, 14, 244. [Google Scholar] [CrossRef] [PubMed]
- Pintore, M.D.; Ceballos, L.; Iulini, B.; Tomassone, L.; Pautasso, A.; Corbellini, D.; Rizzo, F.; Mandola, M.L.; Bardelli, M.; Peletto, S.; et al. Detection of Invasive Borrelia burgdorferi strains in North-Eastern Piedmont, Italy. Zoonoses Public Health 2014, 62, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Vozmediano, A.; Giglio, G.; Ramassa, E.; Nobili, F.; Rossi, L.; Tomassone, L. Dermacentor marginatus and Dermacentor reticulatus, and Their Infection by SFG Rickettsiae and Francisella-Like Endosymbionts, in Mountain and Periurban Habitats of Northwestern Italy. Vet. Sci. 2020, 7, 157. [Google Scholar] [CrossRef] [PubMed]
- Richomme, C.; Gauthier, D.; Fromont, E. Contact rates and exposure to inter-species disease transmission in mountain ungulates. Epidemiol Infect. 2006, 134, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Michel, A.O.; Mathis, A.; Ryser-Degiorgis, M.P. Babesia spp. in European wild ruminant species: Parasite diversity and risk factors for infection. Vet. Res. 2014, 45, 65. [Google Scholar] [CrossRef] [PubMed]
- Silaghi, C.; Hamel, D.; Thiel, C.; Pfister, K.; Passos, L.M.; Rehbein, S. Genetic variants of Anaplasma phagocytophilum in wild caprine and cervid ungulates from the Alps in Tyrol, Austria. Vector Borne Zoonotic Dis. 2011, 11, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Kauffmann, M.; Rehbein, S.; Hamel, D.; Lutz, W.; Heddergott, M.; Pfister, K.; Silaghi, C. Anaplasma phagocytophilum and Babesia spp. in roe deer (Capreolus capreolus), fallow deer (Dama dama) and mouflon (Ovis musimon) in Germany. Mol. Cell Probes. 2017, 31, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Grassi, L.; Franzo, G.; Martini, M.; Mondin, A.; Cassini, R.; Drigo, M.; Pasotto, D.; Vidorin, E.; Menandro, M.L. Ecotyping of Anaplasma phagocytophilum from Wild Ungulates and Ticks Shows Circulation of Zoonotic Strains in Northeastern Italy. Animals 2021, 11, 310. [Google Scholar] [CrossRef] [PubMed]
- Jaenson, T.G.; Tälleklint, L. Incompetence of roe deer as reservoirs of the Lyme borreliosis spirochete. J. Med. Entomol. 1992, 29, 813–817. [Google Scholar] [CrossRef]
- Telford, S.R., 3rd; Mather, T.N.; Moore, S.I.; Wilson, M.L.; Spielman, A. Incompetence of deer as reservoirs of the Lyme disease spirochete. Am. J. Trop. Med. Hyg. 1988, 39, 105–109. [Google Scholar] [CrossRef]
- Kurtenbach, K.; Sewell, H.S.; Ogden, N.H.; Randolph, S.E.; Nuttall, P.A. Serum complement sensitivity as a key factor in Lyme disease ecology. Infect. Immun. 1998, 66, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Pearson, P.; Rich, C.; Feehan, M.J.R.; Ditchkoff, S.S.; Rich, S.M. White-Tailed Deer Serum Kills the Lyme Disease Spirochete, Borrelia burgdorferi. Vector Borne Zoonotic Dis. 2023, 23, 303–305. [Google Scholar] [CrossRef] [PubMed]

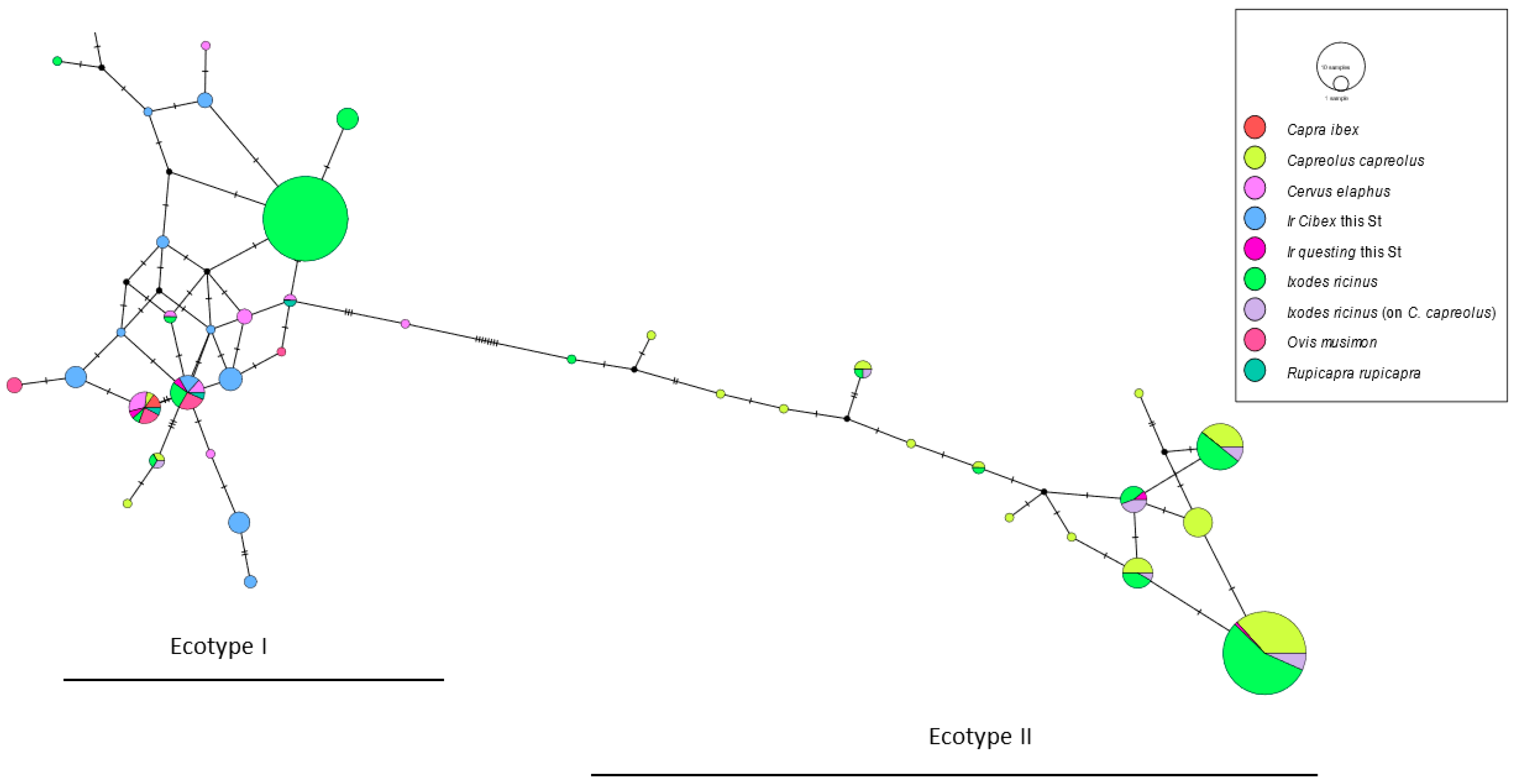

{kind=link}
{kind=link}
| Habitat Type | Altitudinal Range (m a.s.l.) | ||||||
|---|---|---|---|---|---|---|---|
| Beechwood | Sprucewood | Stone-Pinewood | a (<1000) | b (1000–1500) | c (>1500) | ||
| I. ricinus mean no. (min–max) | Nymphs | 4.2 (0–31) | 2.3 (0–13) | 0.3 (0–3) | 5.5 (0–31) | 4.7 (0–26) | 1.0 (0–9) |
| Adults | 0.9 (0–7) | 1 (0–19) | 0.08 (0–3) | 1.3 (0–17) | 1.6 (0–19) | 0.3 (0–7) | |
| Prevalence of Tick-Borne Pathogens (%, 95% CI) | ||||
|---|---|---|---|---|
| Tick-borne pathogens | Questing Ixodes ricinus (n) | I. ricinus on Capra ibex (n) | ||
| Nymphs (n = 24) | Females (n = 68) | Males (n = 38) | Females (n = 87) | |
| Borrelia burgdorferi s.l. | 20.8 (7.1–42.1) | 29.4 (19.0–41.7) | 0 (0–9.3) | 0 (0–4.1) |
| 27.2 (18.4–37.4) | 0 (0–2.9) | |||
| Rickettsia spp. | 16.7 (4.7–37.4) | 22.1 (12.9–33.8) | 13.1 (4.4–28.1) | 27.6 (18.5–38.2) |
| 20.6 (12.9–30.4) | 23.2 (16.1–31.6) | |||
| Anaplasma phagocytophilum | 0 (0–14.2) | 5.9 (1.6–14.4) | 5.2 (0.6–17.7) | 42.5 (32.0–53.6) |
| 4.3% (1.2–10.8) | 31.2% (23.2–40.1) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menzano, A.; Tizzani, P.; Farber, M.D.; Garcia-Vozmediano, A.; Martinelli, L.; Rossi, L.; Tomassone, L. Zoonotic Tick-Borne Pathogens in Ticks from Vegetation and Alpine Ibex (Capra ibex) in the Maritime Alps, Italy. Animals 2024, 14, 2251. https://doi.org/10.3390/ani14152251
Menzano A, Tizzani P, Farber MD, Garcia-Vozmediano A, Martinelli L, Rossi L, Tomassone L. Zoonotic Tick-Borne Pathogens in Ticks from Vegetation and Alpine Ibex (Capra ibex) in the Maritime Alps, Italy. Animals. 2024; 14(15):2251. https://doi.org/10.3390/ani14152251
Chicago/Turabian StyleMenzano, Arianna, Paolo Tizzani, Marisa Diana Farber, Aitor Garcia-Vozmediano, Laura Martinelli, Luca Rossi, and Laura Tomassone. 2024. "Zoonotic Tick-Borne Pathogens in Ticks from Vegetation and Alpine Ibex (Capra ibex) in the Maritime Alps, Italy" Animals 14, no. 15: 2251. https://doi.org/10.3390/ani14152251
APA StyleMenzano, A., Tizzani, P., Farber, M. D., Garcia-Vozmediano, A., Martinelli, L., Rossi, L., & Tomassone, L. (2024). Zoonotic Tick-Borne Pathogens in Ticks from Vegetation and Alpine Ibex (Capra ibex) in the Maritime Alps, Italy. Animals, 14(15), 2251. https://doi.org/10.3390/ani14152251