

Actinobacillus pleuropneumoniae Serotypes by Multiplex PCR Identification and Evaluation of Lung Lesions in Pigs from Piedmont (Italy) Farms

 , , ,
, , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Gross Evaluation and Sample Collection
2.2. Histopathological Evaluation
2.3. APP Bacterial Isolation
2.4. Microbiological Investigations
2.5. DNA Extraction and Multiplex PCR
3. Results
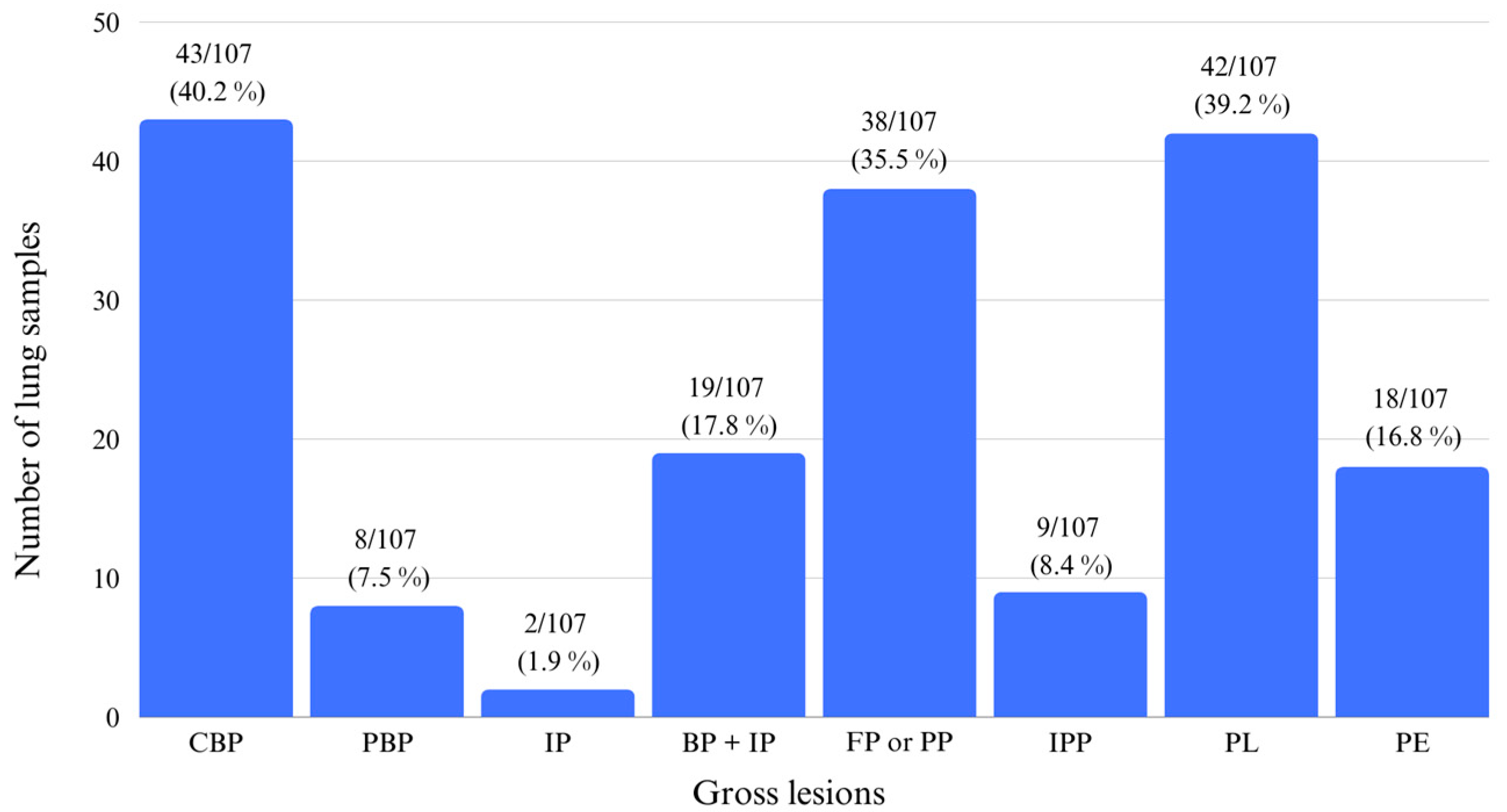
3.1. Gross Evaluation
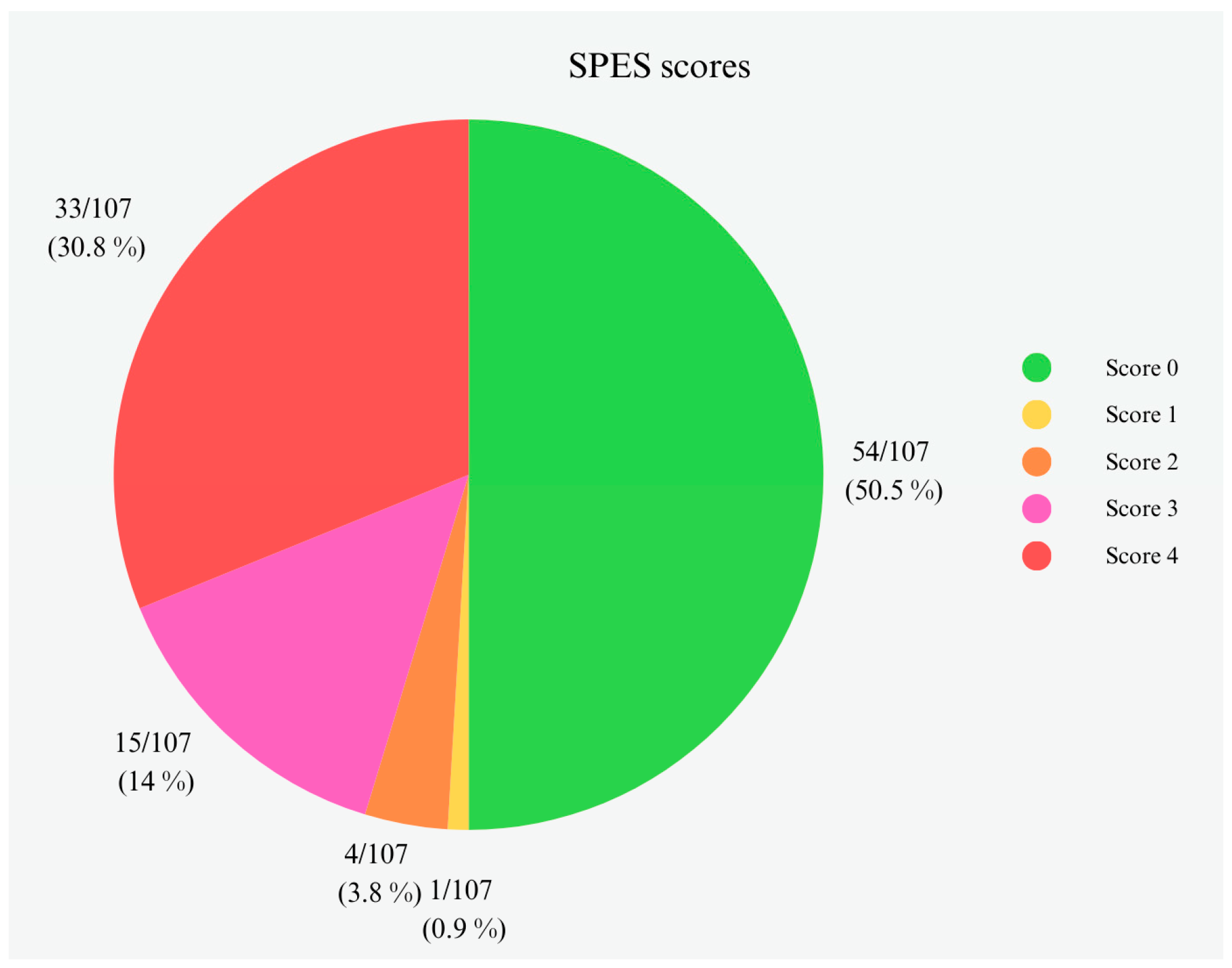
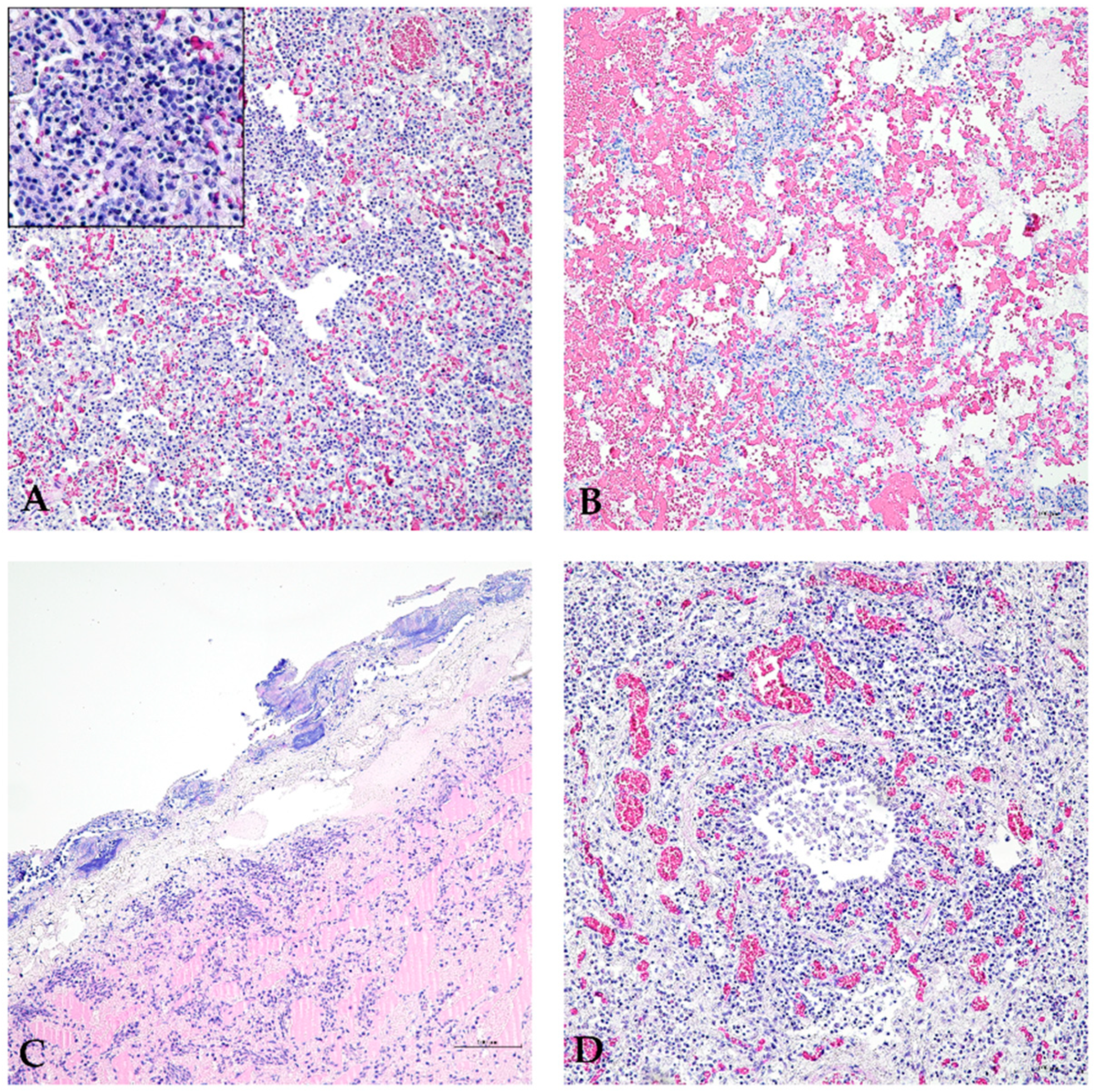
3.2. Histopathological Evaluation
3.3. Microbiological Investigations
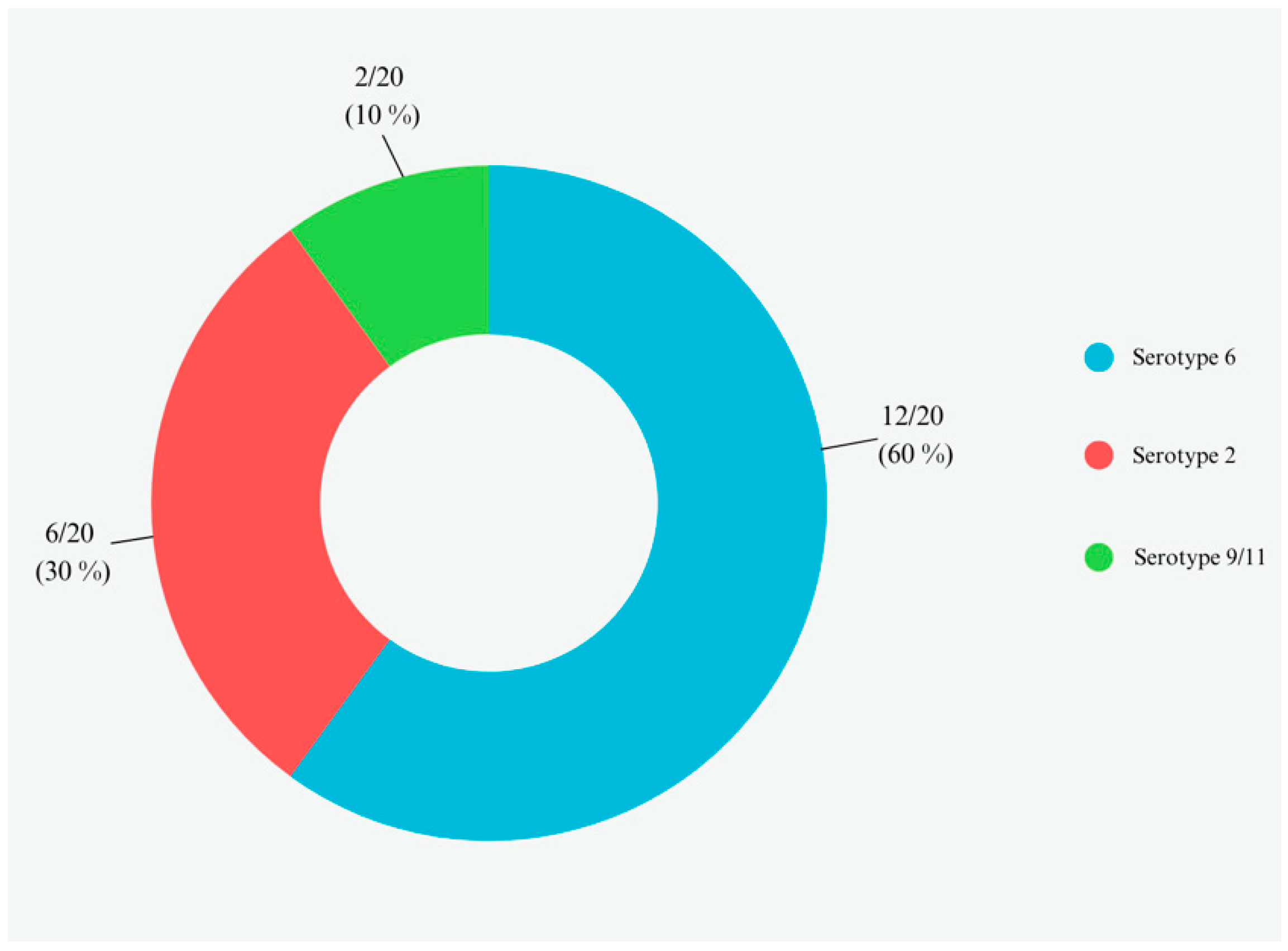
3.4. Serotypes Assignment of APP from Isolated Colonies and Lung Swabs by Multiplex PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Konze, S.A.; Abraham, W.R.; Goethe, E.; Surges, E.; Kuypers, M.M.M.; Hoeltig, D.; Meens, J.; Vogel, C.; Stiesch, M.; Valentin-Weigand, P.; et al. Link between Heterotrophic Carbon Fixation and Virulence in the Porcine Lung Pathogen Actinobacillus Pleuropneumoniae. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef] [PubMed]
- Holmer, I.; Salomonsen, C.M.; Jorsal, S.E.; Astrup, L.B.; Jensen, V.F.; Høg, B.B.; Pedersen, K. Antibiotic Resistance in Porcine Pathogenic Bacteria and Relation to Antibiotic Usage. BMC Vet. Res. 2019, 15, 449. [Google Scholar] [CrossRef] [PubMed]
- Giacominelli-Stuffler, R.; Marruchella, G.; Storelli, M.M.; Sabatucci, A.; Angelucci, C.B.; Maccarrone, M. 5-Lipoxygenase and Cyclooxygenase-2 in the Lungs of Pigs Naturally Affected by Enzootic Pneumonia and Porcine Pleuropneumonia. Res. Vet. Sci. 2012, 93, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Srijuntongsiri, G.; Mhoowai, A.; Samngamnim, S.; Assavacheep, P.; Bossé, J.T.; Langford, P.R.; Posayapisit, N.; Leartsakulpanich, U.; Songsungthong, W. Novel DNA Markers for Identification of Actinobacillus Pleuropneumoniae. Microbiol. Spectr. 2022, 10, e01311-21. [Google Scholar] [CrossRef] [PubMed]
- Stringer, O.W.; Li, Y.; Bossé, J.T.; Langford, P.R. JMM Profile: Actinobacillus Pleuropneumoniae: A Major Cause of Lung Disease in Pigs but Difficult to Control and Eradicate. J. Med. Microbiol. 2022, 71, 001483. [Google Scholar] [CrossRef] [PubMed]
- Sárközi, R.; Makrai, L.; Fodor, L. Isolation of Biotype 1 Serotype 12 and Detection of Actinobacillus Pleuropneumoniae from Wild Boars. Pathogens 2022, 11, 505. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, J.; Liu, L.; Zhang, R.; Hao, X.; Han, Q.; Wang, J.; Yuan, W. Direct Detection of Actinobacillus Pleuropneumoniae in Swine Lungs and Tonsils by Real-Time Recombinase Polymerase Amplification Assay. Mol. Cell. Probes 2019, 45, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Sassu, E.L.; Bossé, J.T.; Tobias, T.J.; Gottschalk, M.; Langford, P.R.; Hennig-Pauka, I. Update on Actinobacillus Pleuropneumoniae—Knowledge, Gaps and Challenges. Transbound. Emerg. Dis. 2018, 65, 72–90. [Google Scholar] [CrossRef] [PubMed]
- Stringer, O.W.; Bossé, J.T.; Lacouture, S.; Gottschalk, M.; Fodor, L.; Angen, Ø.; Velazquez, E.; Penny, P.; Lei, L.; Langford, P.R.; et al. Proposal of Actinobacillus Pleuropneumoniae Serovar 19, and Reformulation of Previous Multiplex PCRs for Capsule-Specific Typing of All Known Serovars. Vet. Microbiol. 2021, 255, 109021. [Google Scholar] [CrossRef] [PubMed]
- Sipos, W.; Cvjetković, V.; Dobrokes, B.; Sipos, S. Evaluation of the Efficacy of a Vaccination Program against Actinobacillus Pleuropneumoniae Based on Lung-scoring at Slaughter. Animals 2021, 11, 2778. [Google Scholar] [CrossRef] [PubMed]
- Baraldi, T.G.; Cruz, N.R.N.; Pereira, D.A.; Galdeano, J.V.B.; Gatto, I.R.H.; Silva, A.F.D.; Panzardi, A.; Linhares, D.C.L.; Mathias, L.A.; De Oliveira, L.G. Antibodies against Actinobacillus Pleuropneumoniae, Mycoplasma Hyopneumoniae and Influenza Virus and Their Relationships with Risk Factors, Clinical Signs and Lung Lesions in Pig Farms with One-Site Production Systems in Brazil. Prev. Vet. Med. 2019, 171, 104748. [Google Scholar] [CrossRef] [PubMed]
- Stringer, O.W.; Bossé, J.T.; Lacouture, S.; Gottschalk, M.; Fodor, L.; Angen, Ø.; Velazquez, E.; Penny, P.; Lei, L.; Langford, P.R.; et al. Rapid Detection and Typing of Actinobacillus Pleuropneumoniae Serovars Directly from Clinical Samples: Combining FTA® Card Technology with Multiplex PCR. Front. Vet. Sci. 2021, 8, 728660. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, R.; Roychoudhury, P.; Kumar, S.; Dutta, S.; Konwar, N.; Subudhi, P.K.; Dutta, T.K. Rapid Detection of Actinobacillus Pleuropneumoniae Targeting the apxIVA Gene for Diagnosis of Contagious Porcine Pleuropneumonia in Pigs by Polymerase Spiral Reaction. Lett. Appl. Microbiol. 2022, 75, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.S.; Matias, D.N.; Poor, A.P.; Dutra, M.C.; Moreno, L.Z.; Parra, B.M.; Silva, A.P.S.; Matajira, C.E.C.; de Moura Gomes, V.T.; Barbosa, M.R.F.; et al. Causes of Sow Mortality and Risks to Post-Mortem Findings in a Brazilian Intensive Swine Production System. Animals 2022, 12, 1804. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Won, H.; Shin, M.K.; Oh, M.W.; Shim, S.; Yoon, I.; Yoo, H.S. Development of Actinobacillus Pleuropneumoniae ApxI, ApxII, and ApxIII-Specific ELISA Methods for Evaluation of Vaccine Efficiency. J. Vet. Sci. 2019, 20, e2. [Google Scholar] [CrossRef] [PubMed]
- Sørenson, V.; Jorsal, S.E.; Mousing, J. Diseases of the Respiratory System. In Diseases of Swine (9 ed., pp. 149–177); Blackwell Publishing Professional: Ames, IA, USA, 2006; pp. 149–178. ISBN 978-0-8138-1703-3. [Google Scholar]
- Madec, F.; Derrien, H. Frequency, Intensity and Localization of Pulmonary Lesions in Bacon Pigs: Results of a First Series of Observations at the Slaughter-House. Ann. De Zootech. 1981, 30, 380. [Google Scholar] [CrossRef]
- Madec, F.; Kobish, M. A Survey of Pulmonary Lesions in Bacon Pigs (Observations Made at the Slaughterhouse). Ann. de Zootech. 1982, 31, 341. [Google Scholar] [CrossRef]
- Dottori, M.; Nigrelli, A.D.; Bonilauri, P.; Merialdi, G.; Gozio, S.; Cominotti, F. Proposta di un nuovo sistema di punteggiatura delle pleuriti suine in sede di macellazione. La griglia S.P.E.S. (Slaughterhouse Pleuritis Evaluation System). Large Anim. Rev. 2007, 13, 161–165. [Google Scholar]
- Scollo, A.; Abbas, M.; Contiero, B.; Gottardo, F. Undocked Tails, Mycoplasma-like Lesions and Gastric Ulcers in Slaughtering Pigs: What Connection? Animals 2023, 13, 305. [Google Scholar] [CrossRef] [PubMed]
- Amatucci, L.; Luise, D.; Luppi, A.; Virdis, S.; Prosperi, A.; Cirelli, A.; Bosco, C.; Trevisi, P. Evaluation of Carcass Quality, Body and Pulmonary Lesions Detected at the Abattoir in Heavy Pigs Subjected or Not to Tail Docking. Porc. Health Manag. 2023, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- Papatsiros, V.G.; Stylianaki, I.; Tsokana, C.N.; Papakonstantinou, G.; Christophorou, M.; Papaioannou, N.; Athanasiou, L.V. Histopathological Lesions of Actinobacillus Pleuropneumoniae Serotype 8 in Infected Pigs. Vet. Res. Forum. 2023, 14, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Kardos, G.; Sárközi, R.; Laczkó, L.; Marton, S.; Makrai, L.; Bányai, K.; Fodor, L. Genetic Diversity of Actinobacillus Pleuropneumoniae Serovars in Hungary. Vet. Sci. 2022, 9, 511. [Google Scholar] [CrossRef]
- Sárközi, R.; Makrai, L.; Fodor, L. Actinobacillus Pleuropneumoniae Serotypes in Hungary. Acta Vet. Hung. 2018, 66, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, J.; Valls, L.; Martínez, E.; Riera, P. Isolation Rates, Serovars, and Toxin Genotypes of Nicotinamide Adenine Dinucleotide-Independent Actinobacillus Pleuropneumoniae among Pigs Suffering from Pleuropneumonia in Spain. J. Vet. Diagn. Investig. 2009, 21, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Schuwerk, L.; Hoeltig, D.; Waldmann, K.-H.; Valentin-Weigand, P.; Rohde, J. Sero- and Apx-Typing of German Actinobacillus Pleuropneumoniae Field Isolates from 2010 to 2019 Reveals a Predominance of Serovar 2 with Regular Apx-Profile. Vet. Res. 2021, 52, 10. [Google Scholar] [CrossRef] [PubMed]
- Šatrán, P.; Nedbalcová, K. Prevalence of Serotypes, Production of Apx Toxins, and Antibiotic Resistance in Strains of Actinobacillus Pleuropneumoniae Isolated in the Czech Republic. Vet. Med. 2002, 47, 92–98. [Google Scholar] [CrossRef]
- Kucerova, Z.; Jaglic, Z.; Ondriasova, R.; Nedbalcova, K. Serotype Distribution of Actinobacillus Pleuropneumoniae Isolated from Porcine Pleuropneumonia in the Czech Republicduring Period 2003–2004. Vet. Med. 2005, 50, 355–360. [Google Scholar] [CrossRef]
- Li, Y.; Bossé, J.T.; Williamson, S.M.; Maskell, D.J.; Tucker, A.W.; Wren, B.W.; Rycroft, A.N.; Langford, P.R. Actinobacillus Pleuropneumoniae Serovar 8 Predominates in England and Wales. Vet. Rec. 2016, 179, 276. [Google Scholar] [CrossRef] [PubMed]
- Lacouture, S.; Gottschalk, M. Distribution of Actinobacillus Pleuropneumoniae (from 2015 to June 2020) and Glaesserella Parasuis (from 2017 to June 2020) Serotypes Isolated from Diseased Pigs in Quebec. Can. Vet. J. 2020, 61, 1261–1263. [Google Scholar] [PubMed]
- Gottschalk, M.; Lacouture, S. Canada: Distribution of Streptococcus Suis (from 2012 to 2014) and Actinobacillus Pleuropneumoniae (from 2011 to 2014) Serotypes Isolated from Diseased Pigs. Can. Vet. J. 2015, 56, 1093–1094. [Google Scholar] [PubMed]
- Bossé, J.T.; Li, Y.; Sárközi, R.; Fodor, L.; Lacouture, S.; Gottschalk, M.; Casas Amoribieta, M.; Angen, Ø.; Nedbalcova, K.; Holden, M.T.G.; et al. Proposal of Serovars 17 and 18 of Actinobacillus Pleuropneumoniae Based on Serological and Genotypic Analysis. Vet. Microbiol. 2018, 217, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Ogawa, T.; Fukamizu, D.; Morinaga, Y.; Kusumoto, M. Nucleotide Sequence Analysis of a DNA Region Involved in Capsular Polysaccharide Biosynthesis Reveals the Molecular Basis of the Nontypeability of Two Actinobacillus Pleuropneumoniae Isolates. J. Vet. Diagn. Investig. 2016, 28, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Blackall, P.J.; Klaasen, H.L.B.M.; Van Den Bosch, H.; Kuhnert, P.; Frey, J. Proposal of a New Serovar of Actinobacillus Pleuropneumoniae: Serovar 15. Vet. Microbiol. 2002, 84, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bossé, J.T.; Stringer, O.W.; Hennig-Pauka, I.; Mortensen, P.; Langford, P.R. Detection of Novel Actinobacillus Pleuropneumoniae Serovars by Multiplex PCR: A Cautionary Tale. Microbiol. Spectr. 2023, 11, e04461-22. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pulmonary Lobe | Score (Mean ± sd) |
|---|---|
| Left cranial | 1.99 ± 1.41 |
| Left medium | 2.38 ± 1.4 |
| Left caudal | 2.17 ± 1.2 |
| Right cranial | 2.2 ± 1.32 |
| Right medium | 2.64 ± 1.32 |
| Right accessory | 2.42 ± 1.52 |
| Right caudal | 2.26 ± 1.22 |
| Microorganisms | Number of Infected Individuals (%) |
|---|---|
| Porcine reproductive and respiratory syndrome virus (PRRSV) | 31 (29%) |
| Mesomycoplasma hyopneumoniae | 11 (10.3%) |
| Escherichia coli | 10 (9.3%) |
| Streptococcus suis | 10 (9.3%) |
| Pasteurella multocida | 9 (8.4%) |
| Actinobacillus lignieresii | 5 (4.7%) |
| Trueperella pyogenes | 4 (3.7%) |
| Bordetella bronchiseptica | 3 (2.8%) |
| Staphylococcus aureus | 3 (2.8%) |
| Staphylococcus spp. | 1 (1.7%) |
| Escherichia spp. | 1 (1.7%) |
| Swine Influenza Virus | 1 (1.7%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuccato, M.; Divari, S.; Ciaramita, S.; Sereno, A.; Campelli, D.; Biolatti, P.G.; Biolatti, B.; Meliota, F.; Bollo, E.; Cannizzo, F.T. Actinobacillus pleuropneumoniae Serotypes by Multiplex PCR Identification and Evaluation of Lung Lesions in Pigs from Piedmont (Italy) Farms. Animals 2024, 14, 2255. https://doi.org/10.3390/ani14152255
Cuccato M, Divari S, Ciaramita S, Sereno A, Campelli D, Biolatti PG, Biolatti B, Meliota F, Bollo E, Cannizzo FT. Actinobacillus pleuropneumoniae Serotypes by Multiplex PCR Identification and Evaluation of Lung Lesions in Pigs from Piedmont (Italy) Farms. Animals. 2024; 14(15):2255. https://doi.org/10.3390/ani14152255
Chicago/Turabian StyleCuccato, Matteo, Sara Divari, Silvia Ciaramita, Alessandra Sereno, Domenico Campelli, Pier Giuseppe Biolatti, Bartolomeo Biolatti, Francesco Meliota, Enrico Bollo, and Francesca Tiziana Cannizzo. 2024. "Actinobacillus pleuropneumoniae Serotypes by Multiplex PCR Identification and Evaluation of Lung Lesions in Pigs from Piedmont (Italy) Farms" Animals 14, no. 15: 2255. https://doi.org/10.3390/ani14152255
APA StyleCuccato, M., Divari, S., Ciaramita, S., Sereno, A., Campelli, D., Biolatti, P. G., Biolatti, B., Meliota, F., Bollo, E., & Cannizzo, F. T. (2024). Actinobacillus pleuropneumoniae Serotypes by Multiplex PCR Identification and Evaluation of Lung Lesions in Pigs from Piedmont (Italy) Farms. Animals, 14(15), 2255. https://doi.org/10.3390/ani14152255