
Relating Wildlife Camera Trap Data to Tick Abundance: Testing the Relationship in Different Habitats

 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
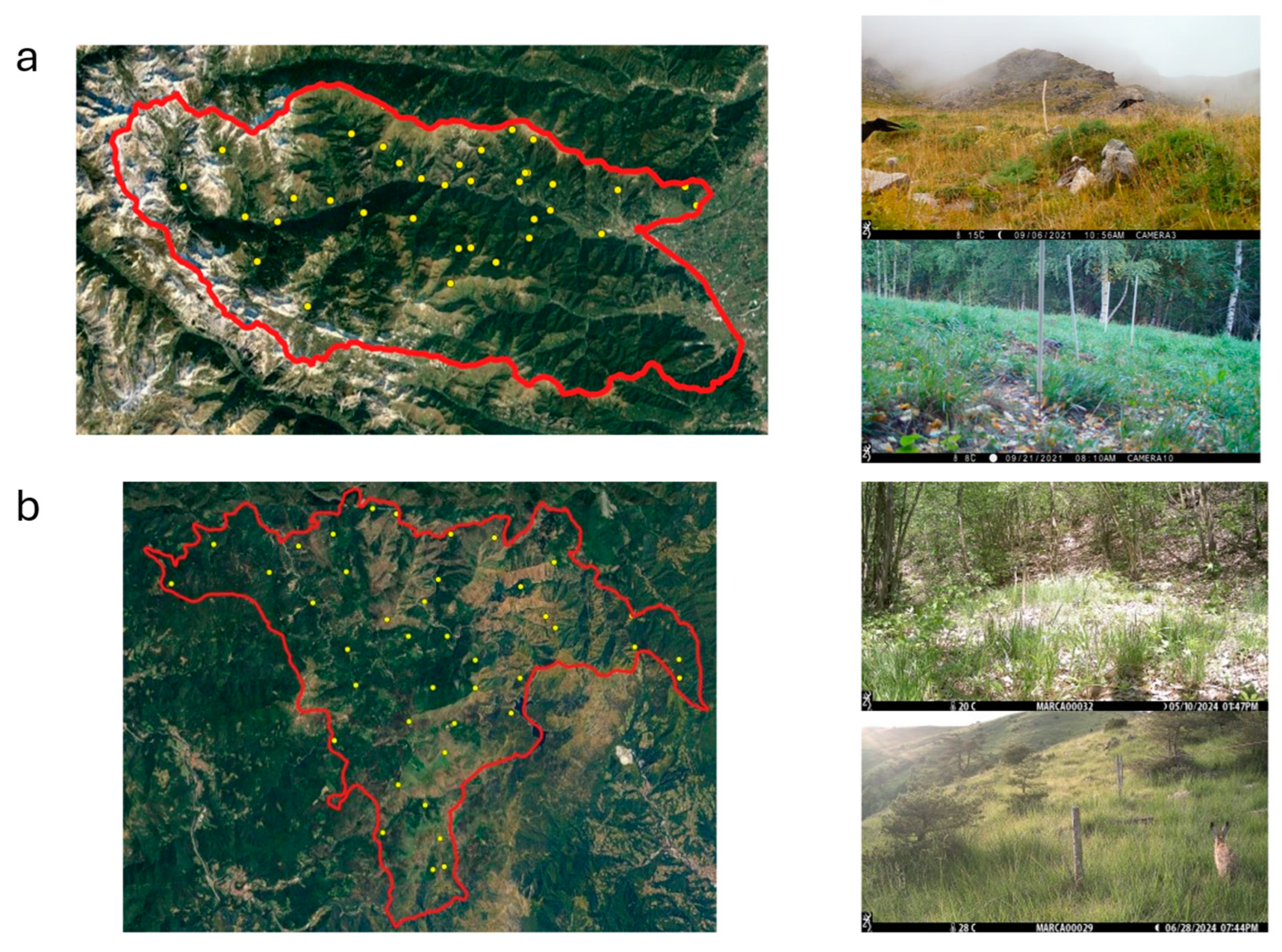
2.1. Study Areas
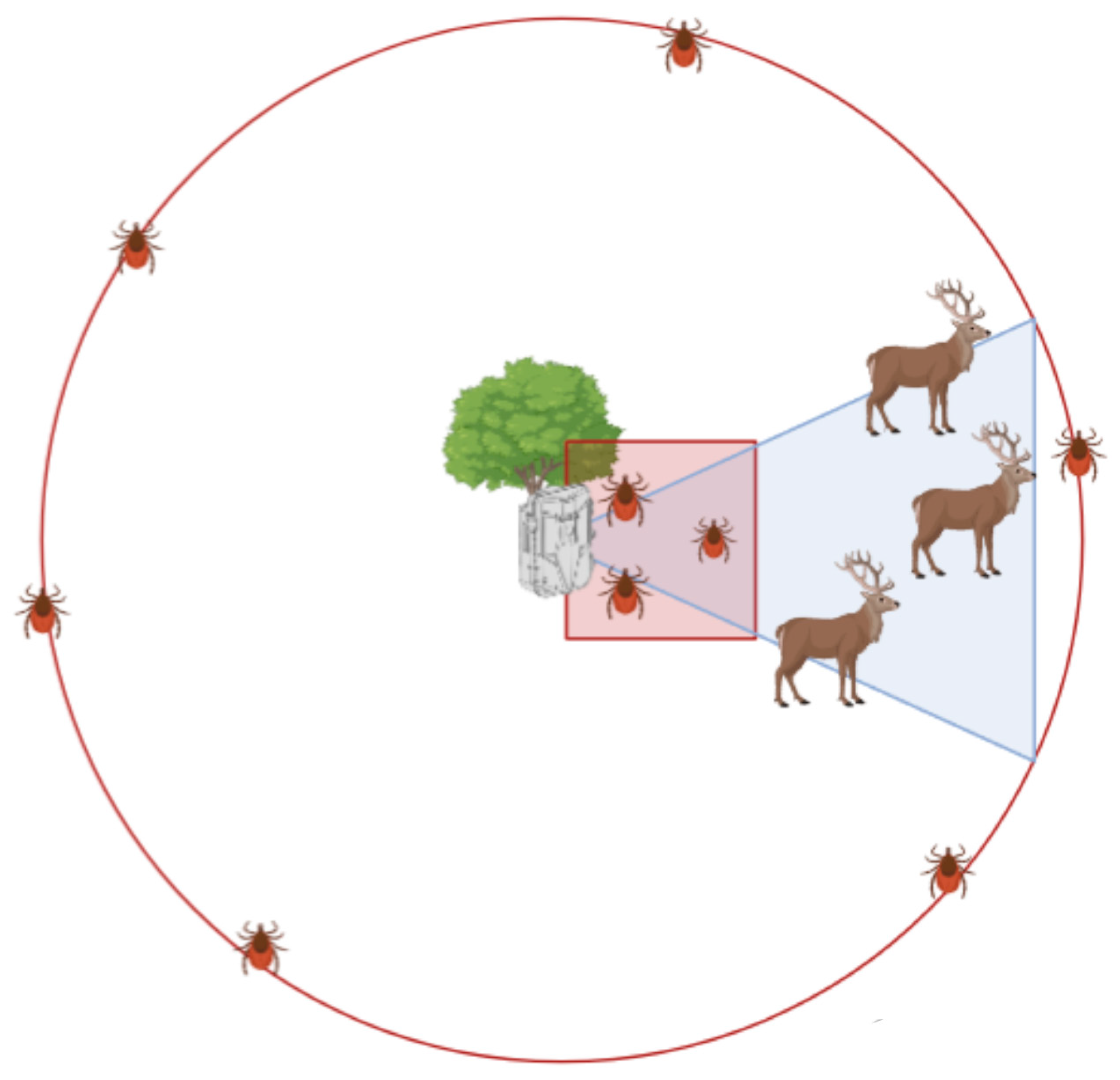
2.2. Sampling Design
2.3. Data Preparation
- (a)
- Tick abundance. Ticks collected from dragging were stored in ethanol 70% and later identified with morphological keys [19,20,21]. Ticks were identified with a Nikon SMZ1500 stereomicroscope (Nikon Corporation, Tokyo, Japan) and Zeiss Axiolab 5 microscope (Carl Zeiss AG, Oberkochen, Germany). Only ticks collected at T1 were considered in the analysis, and abundance was calculated with the sampling point referring to the total area sampled via the two dragging transects;
- (b)
- Wildlife TO. We focused on species that had good detectability in camera traps, thus extracting TO for wild boar, red deer, roe deer, red fox, badger, pine marten, and beech marten. Camera trap pictures were analyzed to extract, for each individual, the following information: sampling point, date of passage, and time spent in front of the camera (calculated as the number of seconds between the first picture and the last). As the aim of this analysis was to calculate the species TO, no individual recognition was required and, for simplicity, an individual exiting the field of view and entering it again was considered as a new individual, so that only the actual time spent in front of the camera trap was considered. In the case that more than one individual appeared in the picture, the time of each animal was calculated separately and eventually summed. As species were unevenly represented in the two study areas, we merged seconds per sampling point for mesocarnivores, and we decided to discard the red deer for its reduced presence and being localized in one study area only, thus implementing in the analysis the TO for roe deer, wild boar, and mesocarnivores;
- (c)
- (d)
- MaxSD. We registered, for each sampling point, the daily mean temperature and humidity, and calculated the daily mean SD (as in [24] per sampling point, from which we derived the maxSD per sampling point in the whole period. Mean and minimum SD were considered but ultimately discarded, based on the high collinearity with max SD and considering that the max SD was the most limiting factor to tick activity [25];
- (e)
- Altitude. Altitude was registered with a GPS device (GPSmap 60CSx, Garmin Ltd., Olathe, KS, USA) for each sampling point.
2.4. Statistical Analysis
3. Results
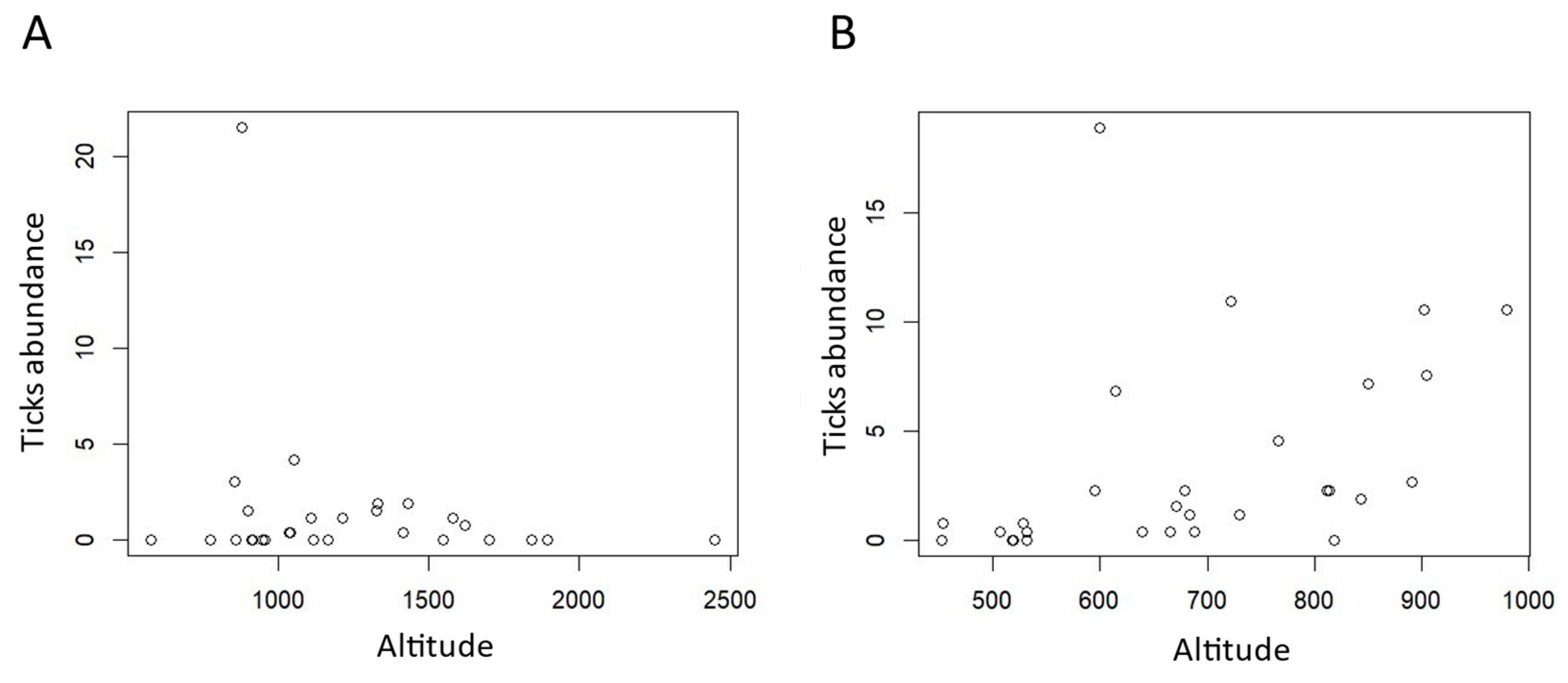
3.1. Tick Transects and Camera Trap Pictures
3.2. Temporal Occupancy Model
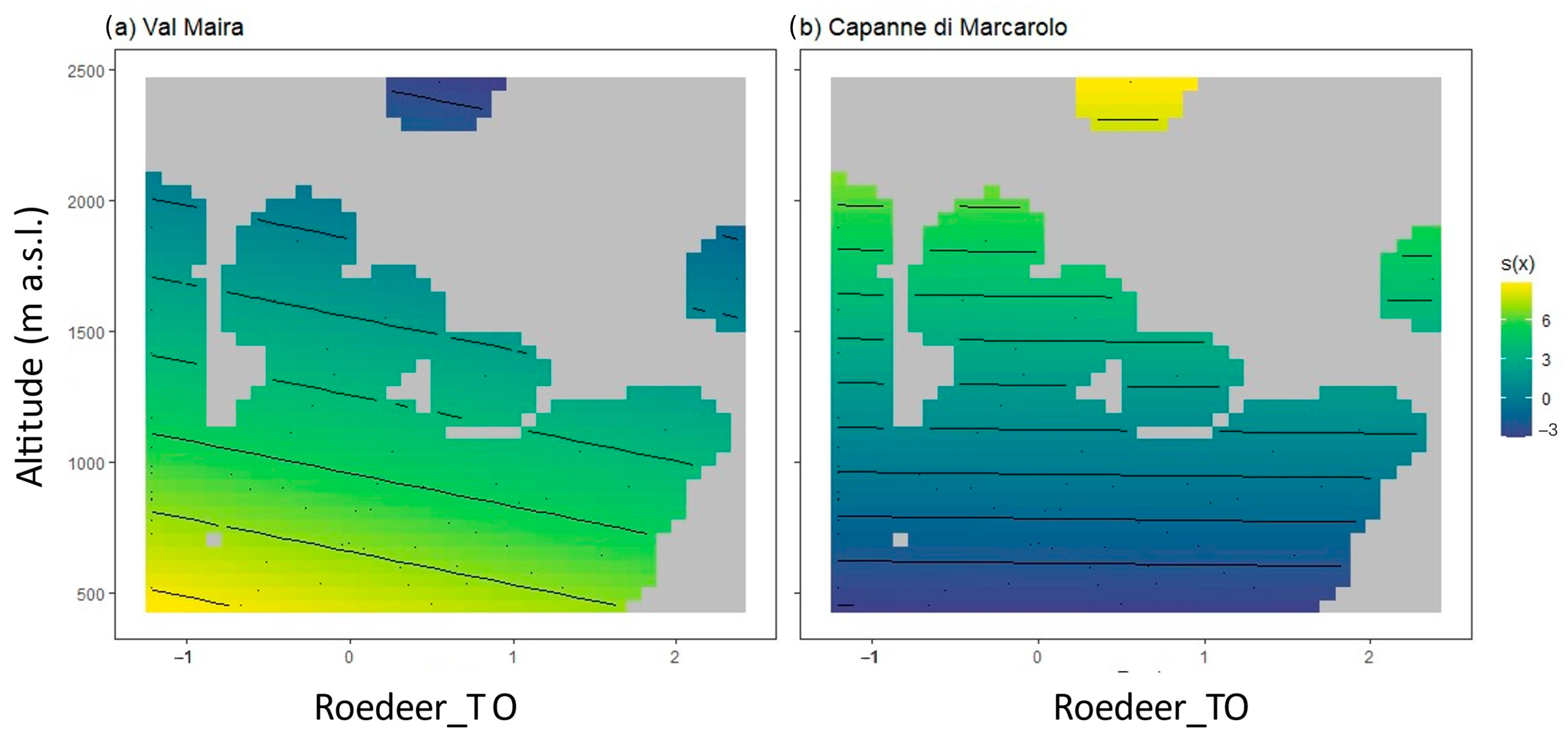
3.3. Variations of TO
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eisen, R.J.; Kugeler, K.J.; Eisen, L.; Beard, C.B.; Paddock, C.D. Tick-borne zoonoses in the United States: Persistent and emerging threats to human health. ILAR J. 2017, 58, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Vannier, E.; Krause, P.J. Babesiosis in China, an emerging threat. Lancet Infect. Dis. 2015, 15, 137–139. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.-C.; Golovljova, I.; Jaenson, T.G.T.; Jensen, J.-K.; Jensen, P.M. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasit. Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Materna, J.; Daniel, M.; Metelka, L.; Harčarik, J. The vertical distribution, density and the development of the tick Ixodes ricinus in mountain areas influenced by climate changes (The Krkonoše Mts., Czech Republic). Int. J. Med. Microbiol. 2008, 298, 25–37. [Google Scholar] [CrossRef]
- Gilbert, L. Altitudinal patterns of tick and host abundance: A potential role for climate change in regulating tick-borne diseases? Oecologia 2010, 162, 217–225. [Google Scholar] [CrossRef]
- Gern, L.; Cadenas, F.M.; Burri, C. Influence of some climatic factors on Ixodes ricinus ticks studied along altitudinal gradients in two geographic regions in Switzerland. Int. J. Med. Microbiol. 2008, 298, 55–59. [Google Scholar] [CrossRef]
- Rand, P.W.; Lubelczyk, C.; Lavigne, G.R.; Elias, S.; Holman, M.S.; Lacombe, E.H.; Smith, R.P. Deer density and the abundance of Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 2003, 40, 179–184. [Google Scholar] [CrossRef]
- Ginsberg, H.S.; Rulison, E.L.; Miller, J.L.; Pang, G.; Arsnoe, I.M.; Hickling, G.J.; Ogden, N.H.; LeBrun, R.A.; Tsao, J.I. Local abundance of Ixodes scapularis in forests: Effects of environmental moisture, vegetation characteristics, and host abundance. Ticks Tick. Borne. Dis. 2020, 11, 101271. [Google Scholar] [CrossRef]
- Consortium, E.; Grignolio, S.; Apollonio, M.; Brivio, F.; Vicente, J.; Acevedo, P.; Petrovic, K.; Keuling, O. Guidance on estimation of abundance and density data of wild ruminant population: Methods, challenges, possibilities. EFSA Support. Publ. 2020, 17, 1876E. [Google Scholar]
- Esser, H.J.; Herre, E.A.; Kays, R.; Liefting, Y.; Jansen, P.A. Local host-tick coextinction in neotropical forest fragments. Int. J. Parasitol. 2019, 49, 225–233. [Google Scholar] [CrossRef]
- Domínguez, L.; Miranda, R.J.; Torres, S.; Moreno, R.; Ortega, J.; Bermúdez, S.E. Hard tick (Acari: Ixodidae) survey of Oleoducto trail, Soberania National Park, Panama. Ticks Tick. Borne. Dis. 2019, 10, 830–837. [Google Scholar] [CrossRef] [PubMed]
- Iijima, H.; Watari, Y.; Furukawa, T.; Okabe, K. Importance of host abundance and microhabitat in tick abundance. J. Med. Entomol. 2022, 59, 2110–2119. [Google Scholar] [CrossRef] [PubMed]
- Noden, B.H.; Roselli, M.A.; Loss, S.R. Factors influencing abundance of 3 tick species across a gradient of urban development intensity in the US Great Plains. J. Med. Entomol. 2024, 61, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Hofmeester, T.R.; Sprong, H.; Jansen, P.A.; Prins, H.H.T.; Van Wieren, S.E. Deer presence rather than abundance determines the population density of the sheep tick, Ixodes ricinus, in Dutch forests. Parasit. Vectors 2017, 10, 433. [Google Scholar] [CrossRef]
- Takumi, K.; Sprong, H.; Hofmeester, T.R. Impact of vertebrate communities on Ixodes ricinus-borne disease risk in forest areas. Parasit. Vectors 2019, 12, 434. [Google Scholar] [CrossRef]
- Taylor, C.L.; Hochuli, D.F.; Banks, P.B. Activity and movement of small mammal tick hosts at the urban fringes of Sydney, Australia. Wildl. Res. 2023, 50, 927–938. [Google Scholar] [CrossRef]
- Hofmeester, T.R.; Rowcliffe, J.M.; Jansen, P.A. Quantifying the availability of vertebrate hosts to ticks: A camera-trapping approach. Front. Vet. Sci. 2017, 4, 115. [Google Scholar] [CrossRef]
- ENETWILD-consortium; Guerrasio, T.; Pelayo Acevedo, P.; Apollonio, M.; Arnon, A.; Barroqueiro, C.; Belova, O.; Berdión, O.; Blanco-Aguiar, J.A.; Bijl, H. Wild ungulate density data generated by camera trapping in 37 European areas: First output of the European Observatory of Wildlife (EOW). EFSA Support. Publ. 2023, 20, 7892E. [Google Scholar]
- Manilla, G.; Iori, A. Chiave illustrata delle zecche d’Italia. I: Stadi larvali delle specie della sottofamiglia Ixodinae (Acari, Ixodoidea, Ixodidae). Parassitologia 1992, 34, 83. [Google Scholar]
- Manilla, G.; Iori, A. Chiave illustrata delle zecche d’Italia. II: Stadi ninfali delle specie della sottofamiglia Ixodinate (Acari, Ixodoidea, Ixodidae). Parassitologia 1993, 35, 37. [Google Scholar]
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Ticks of Europe and North Africa: A Guide to Species Identification; Springer: Berlin/Heidelberg, Germany, 2018; ISBN 3319637606. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Busetto, L.; Ranghetti, L. MODIStsp: An R package for automatic preprocessing of MODIS Land Products time series. Comput. Geosci. 2016, 97, 40–48. [Google Scholar] [CrossRef]
- Sands, B.O.; Bryer, K.E.; Wall, R. Climate and the seasonal abundance of the tick Dermacentor reticulatus. Med. Vet. Entomol. 2021, 35, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Perret, J.-L.; Rais, O.; Gern, L. Influence of climate on the proportion of Ixodes ricinus nymphs and adults questing in a tick population. J. Med. Entomol. 2004, 41, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Peterson, R.A. Finding Optimal Normalizing Transformations via best Normalize. R J. 2021, 13, 294–313. [Google Scholar] [CrossRef]
- Wood, S.N. Generalized Additive Models: An Introduction with R; CRC Press: Boca Raton, FL, USA, 2017; ISBN 1498728340. [Google Scholar]
- Elias, S.P.; Gardner, A.M.; Maasch, K.A.; Birkel, S.D.; Anderson, N.T.; Rand, P.W.; Lubelczyk, C.B.; Smith, R.P., Jr. A generalized additive model correlating blacklegged ticks with white-tailed deer density, temperature, and humidity in Maine, USA, 1990–2013. J. Med. Entomol. 2021, 58, 125–138. [Google Scholar] [CrossRef]
- McKight, P.E.; Najab, J. The Corsini Encyclopedia of Psychology; Kruskal-Wallis Test; John Wiley & Sons: New York, NY, USA, 2010. [Google Scholar]
- Salman, M.; Tarrés-Call, J. Ticks and Tick-Borne Diseases: Geographical Distribution and Control Strategies in the Euro-Asia Region; CABI: Wallingford, UK, 2013; ISBN 1845938542. [Google Scholar]
- Accorsi, A.; Schiavetti, I.; Listorti, V.; Dellepiane, M.; Masotti, C.; Ercolini, C.; Guardone, L.; Razzuoli, E. Hard Ticks (Ixodidae) from Wildlife in Liguria, Northwest Italy: Tick Species Diversity and Tick-Host Associations. Insects 2022, 13, 199. [Google Scholar] [CrossRef]
- Castillo-Contreras, R.; Magen, L.; Birtles, R.; Varela-Castro, L.; Hall, J.L.; Conejero, C.; Aguilar, X.F.; Colom-Cadena, A.; Lavín, S.; Mentaberre, G. Ticks on wild boar in the metropolitan area of Barcelona (Spain) are infected with spotted fever group rickettsiae. Transbound. Emerg. Dis. 2022, 69, e82–e95. [Google Scholar] [CrossRef]
- Fabri, N.D.; Sprong, H.; Hofmeester, T.R.; Heesterbeek, H.; Donnars, B.F.; Widemo, F.; Ecke, F.; Cromsigt, J.P.G.M. Wild ungulate species differ in their contribution to the transmission of Ixodes ricinus-borne pathogens. Parasit. Vectors 2021, 14, 360. [Google Scholar] [CrossRef]
- Palencia, P.; Vada, R.; Zanet, S.; Calvini, M.; De Giovanni, A.; Gola, G.; Ferroglio, E. Not just pictures: Utility of camera trapping in the context of African Swine fever and wild boar management. Transbound. Emerg. Dis. 2023, 2023, 7820538. [Google Scholar] [CrossRef]
- Danielová, V.; Rudenko, N.; Daniel, M.; Holubová, J.; Materna, J.; Golovchenko, M.; Schwarzová, L. Extension of Ixodes ricinus ticks and agents of tick-borne diseases to mountain areas in the Czech Republic. Int. J. Med. Microbiol. 2006, 296, 48–53. [Google Scholar] [CrossRef]
- Shchuchinova, L.D.; Kozlova, I.V.; Zlobin, V.I. Influence of altitude on tick-borne encephalitis infection risk in the natural foci of the Altai Republic, Southern Siberia. Ticks Tick. Borne. Dis. 2015, 6, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.-X.; Lai, S.-J.; Yang, Y.; Li, X.-L.; Liu, K.; Yao, H.-W.; Zhou, H.; Li, Y.; Wang, L.-P.; Mu, D. Mapping the distribution of tick-borne encephalitis in mainland China. Ticks Tick. Borne. Dis. 2017, 8, 631–639. [Google Scholar] [CrossRef] [PubMed]
- De Pelsmaeker, N.; Korslund, L.; Steifetten, Ø. High-elevational occurrence of two tick species, Ixodes ricinus and I. trianguliceps, at their northern distribution range. Parasit. Vectors 2021, 14, 161. [Google Scholar] [CrossRef] [PubMed]
- Zubriková, D.; Blaňarová, L.; Hrkľová, G.; Syrota, Y.; Macko, J.; Blahútová, D.; Blažeková, V.; Stanko, M.; Švirlochová, K.; Víchová, B. The Impact of Altitude on Tick-Borne Pathogens at Two Mountain Ranges in Central Slovakia. Pathogens 2024, 13, 586. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Vozmediano, A.; Krawczyk, A.I.; Sprong, H.; Rossi, L.; Ramassa, E.; Tomassone, L. Ticks climb the mountains: Ixodid tick infestation and infection by tick-borne pathogens in the Western Alps. Ticks Tick. Borne. Dis. 2020, 11, 101489. [Google Scholar] [CrossRef]
- Jouda, F.; Perret, J.-L.; Gern, L. Ixodes ricinus density, and distribution and prevalence of Borrelia burgdorferi sensu lato infection along an altitudinal gradient. J. Med. Entomol. 2004, 41, 162–169. [Google Scholar] [CrossRef]
- Kiewra, D.; Kryza, M.; Szymanowski, M. Influence of selected meteorological variables on the questing activity of Ixodes ricinus ticks in Lower Silesia, SW Poland. J. Vector Ecol. 2014, 39, 138–145. [Google Scholar] [CrossRef]
- Rosà, R.; Andreo, V.; Tagliapietra, V.; Baráková, I.; Arnoldi, D.; Hauffe, H.C.; Manica, M.; Rosso, F.; Blaňarová, L.; Bona, M. Effect of climate and habitat type on the spatio-temporal variability of tick-borne bacteria in Europe. Int. J. Environ. Res. Public Health 2018, 15, 732. [Google Scholar] [CrossRef]
- Kjaer, L.J.; Soleng, A.; Edgar, K.S.; Lindstedt, H.E.H.; Paulsen, K.M.; Andreassen, A.K.; Korslund, L.; Kjelland, V.; Slettan, A.; Stuen, S.; et al. Predicting and mapping human risk of exposure to Ixodes ricinus nymphs using climatic and environmental data, Denmark, Norway and Sweden, 2016. Eurosurveillance 2019, 24, 35–45. [Google Scholar] [CrossRef]
- Alonso-Carné, J.; García-Martín, A.; Estrada-Peña, A. Modelling the phenological relationships of questing immature Ixodes ricinus (Ixodidae) using temperature and NDVI data. Zoonoses Public Health 2016, 63, 40–52. [Google Scholar] [CrossRef]
- Vucelja, M.; Krčmar, S.; Habuš, J.; Perko, V.M.; Boljfetić, M.; Bjedov, L.; Margaletić, J. Altitudinal distribution, seasonal dynamics and Borrelia burgdorferi sensu lato infections in hard ticks (Acari: Ixodidae) in different forest communities in Inland Croatia. Sustainability 2023, 15, 4862. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Val Maira | Capanne di Marcarolo | |
|---|---|---|
| Larvae | 143 | 735 |
| Nymphs | 49 | 21 |
| Adults | 6 | 3 |
| Ixodes ricinus | 190 | 350 |
| Haemaphysalis punctata | 7 | 406 |
| Haemaphysalis concinna | 0 | 3 |
| Dermacentor reticulatus | 1 | 0 |
| Val Maira | Capanne di Marcarolo | |
|---|---|---|
| Mesocarnivores | 2644 (26/38) | 511 (20/41) |
| Red deer | 818 (8/38) | Not present |
| Roe deer | 7607 (20/38) | 6361 (26/41) |
| Wild boar | 3021 (16/38) | 2432 (18/41) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vada, R.; Zanet, S.; Occhibove, F.; Fantini, E.; Palencia, P.; Ferroglio, E. Relating Wildlife Camera Trap Data to Tick Abundance: Testing the Relationship in Different Habitats. Animals 2024, 14, 2749. https://doi.org/10.3390/ani14182749
Vada R, Zanet S, Occhibove F, Fantini E, Palencia P, Ferroglio E. Relating Wildlife Camera Trap Data to Tick Abundance: Testing the Relationship in Different Habitats. Animals. 2024; 14(18):2749. https://doi.org/10.3390/ani14182749
Chicago/Turabian StyleVada, Rachele, Stefania Zanet, Flavia Occhibove, Enrica Fantini, Pablo Palencia, and Ezio Ferroglio. 2024. "Relating Wildlife Camera Trap Data to Tick Abundance: Testing the Relationship in Different Habitats" Animals 14, no. 18: 2749. https://doi.org/10.3390/ani14182749
APA StyleVada, R., Zanet, S., Occhibove, F., Fantini, E., Palencia, P., & Ferroglio, E. (2024). Relating Wildlife Camera Trap Data to Tick Abundance: Testing the Relationship in Different Habitats. Animals, 14(18), 2749. https://doi.org/10.3390/ani14182749