
Caveolin 1 Regulates the Tight Junctions between Sertoli Cells and Promotes the Integrity of Blood–Testis Barrier in Yak via the FAK/ERK Signaling Pathway
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. Identification via RNA-seq and Bioinformatics Analysis of miRNAs and mRNAs in Yak Testis and Cryptorchidism
2.3. Hematoxylin–Eosin Staining
2.4. Plasmid Construction, Cloning and Double Digests
2.5. Cell Primary Culture and Identification
2.6. Cell Transfection
2.7. Quantitative Reverse Transcription-PCR
2.8. Immunofluorescence and Immunohistochemistry
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
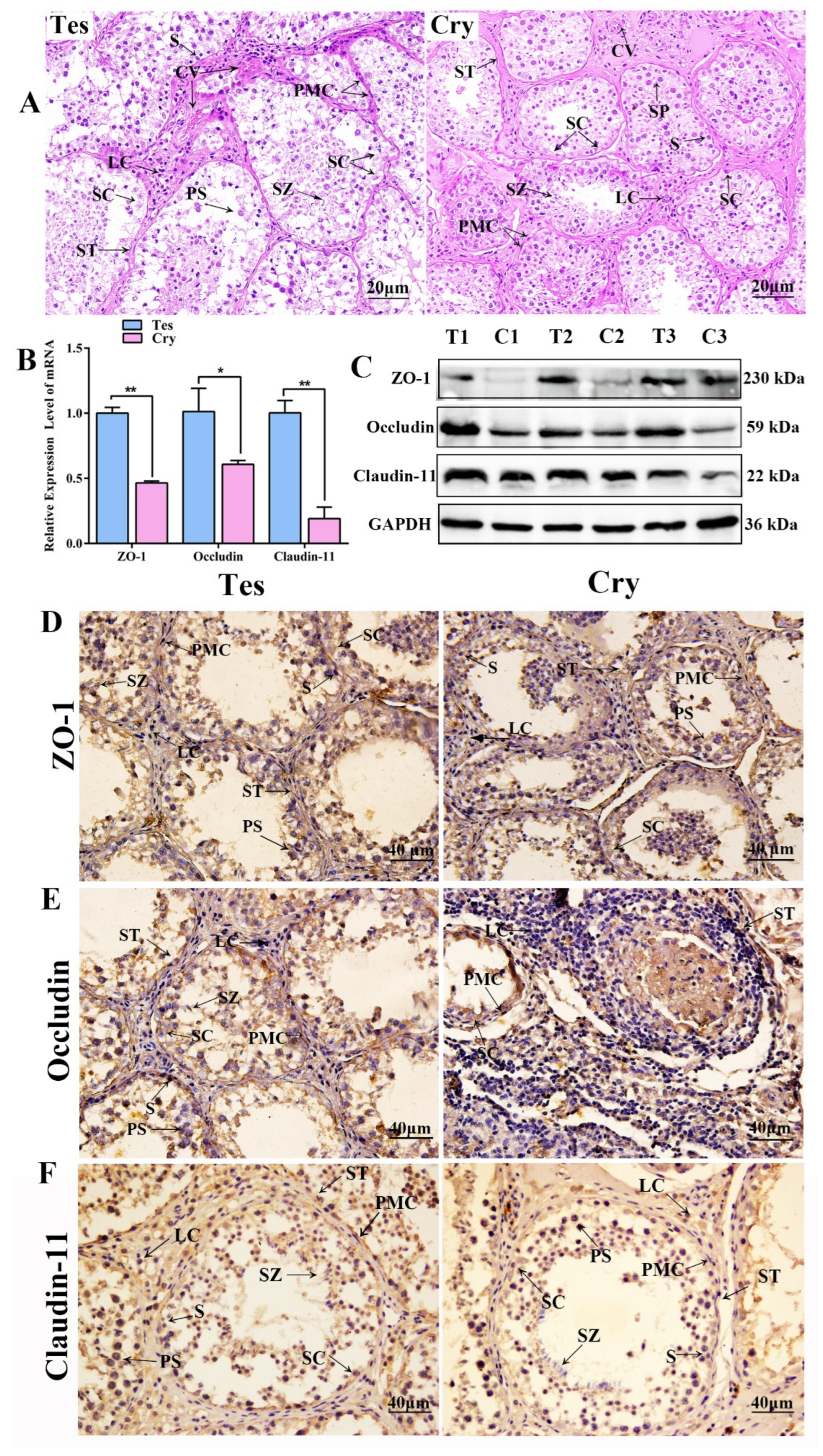
3.1. Cell TJs and BTB Were Detected in yak Testis and Cryptorchidism
3.2. Identification and Functional Enrichment Analysis of Differentially Expressed Genes in the Normal Testis and Cryptorchidism of Yak
3.3. Identifying Differentially Expressed microRNA of Small RNA Sequencing
3.4. Validation of DERs and DEGs
3.5. The Efficiency of p-EGFP-CAV1 Was Detected in Yak Sertoli Cells
3.6. Detection of Silencing Efficiency of the CAV1 Gene
3.7. Up-Regulated CAV1 Promoted TJs and BTBs via FAK/ERK Pathway of Sertoli Cells
3.8. Down-Regulating CAV1 Suppression of TJs and BTB via FAK/ERK Pathway of Sertoli Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goda, Y.; Mizutani, S.; Mizutani, Y.; Kitahara, G.; Siswandi, R.; Wakabayashi, K.; Torisu, S.; Kaneko, Y.; Hidaka, Y.; Osawa, T.; et al. Usefulness of computed tomography for cryptorchidism in bulls. J. Vet. Med. Sci. 2022, 84, 1430–1436. [Google Scholar] [CrossRef] [PubMed]
- St Jean, G.; Gaughan, E.M.; Constable, P.D. Cryptorchidism in North American cattle: Breed predisposition and clinical findings. Theriogenology 1992, 38, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Mieusset, R.; Bujan, L.E.; Massat, G.; Mansat, A.; Pontonnier, F. Inconstant ascending testis as a potential risk factor for spermatogenesis in infertile men with no history of cryptorchism. Hum. Reprod. 1997, 12, 974–979. [Google Scholar] [CrossRef] [PubMed]
- Kellaway, R.C.; Seamark, R.F.; Farrant, R.K. Sterilisation of cattle by induced cryptorchidism. Aust. Vet. J. 1971, 47, 547–550. [Google Scholar] [CrossRef] [PubMed]
- Goto, A.; Yoshida, N.; Nakada, K.; Inoue, Y.; Hisaeda, K.; Inaba, T.; Domoto, N.; Ishiguro, Y.; Itoh, M.; Takahashi, E.; et al. Efficiency of immunocastration with an anti-gonadotropin-releasing hormone vaccine on cryptorchid bulls. J. Vet. Med. Sci. 2023, 85, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Yefimova, M.G.; Buschiazzo, A.; Burel, A.; Lavault, M.T.; Pimentel, C.; Jouve, G.; Jaillard, S.; Jegou, B.; Bourmeyster, N.; Ravel, C. Autophagy is increased in cryptorchid testis resulting in abnormal spermatozoa. Asian J. Androl. 2019, 21, 570–576. [Google Scholar] [CrossRef]
- Pinart, E.; Bonet, S.; Briz, M.D.; Pastor, L.M.; Sancho, S.; García, N.; Badia, E. Morphologic and histochemical study of blood capillaries in boar testes: Effects of abdominal cryptorchidism. Teratology 2001, 63, 42–51. [Google Scholar] [CrossRef]
- Batra, N.V.; DeMarco, R.T.; Bayne, C.E. A narrative review of the history and evidence-base for the timing of orchidopexy for cryptorchidism. J. Pediatr. Urol. 2021, 17, 239–245. [Google Scholar] [CrossRef]
- Cortes, D.; Holt, R.; de Knegt, V.E. Hormonal Aspects of the Pathogenesis and Treatment of Cryptorchidism. Eur. J. Pediatr. Surg. 2016, 26, 409–417. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Mruk, D.D. The blood-testis barrier and its implications for male contraception. Pharmacol. Rev. 2012, 64, 16–64. [Google Scholar] [CrossRef]
- Mruk, D.D.; Cheng, C.Y. The Mammalian Blood-Testis Barrier: Its Biology and Regulation. Endocr. Rev. 2015, 36, 564–591. [Google Scholar] [CrossRef] [PubMed]
- Mital, P.; Hinton, B.T.; Dufour, J.M. The blood-testis and blood-epididymis barriers are more than just their tight junctions. Biol. Reprod. 2011, 84, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Gerber, J.; Heinrich, J.; Brehm, R. Blood-testis barrier and Sertoli cell function: Lessons from SCCx43KO mice. Reprod. (Camb. Engl.) 2016, 151, R15–R27. [Google Scholar] [CrossRef]
- Kato, T.; Mizuno, K.; Nishio, H.; Moritoki, Y.; Kamisawa, H.; Kurokawa, S.; Nakane, A.; Maruyama, T.; Ando, R.; Hayashi, Y.; et al. Disorganization of claudin-11 and dysfunction of the blood-testis barrier during puberty in a cryptorchid rat model. Andrology 2020, 8, 1398–1408. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Kim, S.O.; Cho, K.A.; Song, S.H.; Kang, T.W.; Park, K.; Kwon, D. Loss of Caveolin 1 is Associated With the Expression of Aquaporin 1 and Bladder Dysfunction in Mice. Int. Neurourol. J. 2015, 19, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, Z.; Yang, Y.; Dai, Y.; Gao, X. Polypyrimidine tract-binding protein 1 regulates the Sertoli cell blood-testis barrier by promoting the expression of tight junction proteins. Exp. Ther. Med. 2021, 22, 847. [Google Scholar] [CrossRef] [PubMed]
- Ishii, K.; Kanatsu-Shinohara, M.; Toyokuni, S.; Shinohara, T. FGF2 mediates mouse spermatogonial stem cell self-renewal via upregulation of Etv5 and Bcl6b through MAP2K1 activation. Development 2012, 139, 1734–1743. [Google Scholar] [CrossRef]
- Bender, F.; Montoya, M.; Monardes, V.; Leyton, L.; Quest, A.F. Caveolae and caveolae-like membrane domains in cellular signaling and disease: Identification of downstream targets for the tumor suppressor protein caveolin-1. Biol. Res. 2002, 35, 151–167. [Google Scholar] [CrossRef]
- Razani, B.; Schlegel, A.; Lisanti, M.P. Caveolin proteins in signaling, oncogenic transformation and muscular dystrophy. J. Cell Sci. 2000, 113 Pt 12, 2103–2109. [Google Scholar] [CrossRef]
- Minshall, R.D.; Malik, A.B. Transport across the endothelium: Regulation of endothelial permeability. Handb. Exp. Pharmacol. 2006, 176/I, 107–144. [Google Scholar] [CrossRef]
- Nighot, P.K.; Leung, L.; Ma, T.Y. Chloride channel ClC- 2 enhances intestinal epithelial tight junction barrier function via regulation of caveolin-1 and caveolar trafficking of occludin. Exp. Cell Res. 2017, 352, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Hoop, C.L.; Sivanandam, V.N.; Kodali, R.; Srnec, M.N.; van der Wel, P.C. Structural characterization of the caveolin scaffolding domain in association with cholesterol-rich membranes. Biochemistry 2012, 51, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Saxena, S.; Liu, Y.; Joshi, B.; Wong, T.H.; Shankar, J.; Foster, L.J.; Bernatchez, P.; Nabi, I.R. The phospho-caveolin-1 scaffolding domain dampens force fluctuations in focal adhesions and promotes cancer cell migration. Mol. Biol. Cell 2017, 28, 2190–2201. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Xue, X.; You, K.; Fu, J. Caveolin-1 regulates the expression of tight junction proteins during hyperoxia-induced pulmonary epithelial barrier breakdown. Respir. Res. 2016, 17, 50. [Google Scholar] [CrossRef]
- Marchiando, A.M.; Shen, L.; Graham, W.V.; Weber, C.R.; Schwarz, B.T.; Austin, J.R., 2nd; Raleigh, D.R.; Guan, Y.; Watson, A.J.; Montrose, M.H.; et al. Caveolin-1-dependent occludin endocytosis is required for TNF-induced tight junction regulation in vivo. J. Cell Biol. 2010, 189, 111–126. [Google Scholar] [CrossRef]
- Li, T.; Luo, R.; Wang, X.; Wang, H.; Zhao, X.; Guo, Y.; Jiang, H.; Ma, Y. Unraveling Stage-Dependent Expression Patterns of Circular RNAs and Their Related ceRNA Modulation in Ovine Postnatal Testis Development. Front. Cell Dev. Biol. 2021, 9, 627439. [Google Scholar] [CrossRef]
- Gao, Y.; Lu, W.; Jian, L.; Machaty, Z.; Luo, H. Vitamin E promotes ovine Sertoli cell proliferation by regulation of genes associated with cell division and the cell cycle. Anim. Biotechnol. 2022, 33, 392–400. [Google Scholar] [CrossRef]
- Salama, S.A.; Arab, H.H.; Hassan, M.H.; Al Robaian, M.M.; Maghrabi, I.A. Cadmium-induced hepatocellular injury: Modulatory effects of γ-glutamyl cysteine on the biomarkers of inflammation, DNA damage, and apoptotic cell death. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. (GMS) 2019, 52, 74–82. [Google Scholar] [CrossRef]
- França, L.R.; Hess, R.A.; Dufour, J.M.; Hofmann, M.C.; Griswold, M.D. The Sertoli cell: One hundred fifty years of beauty and plasticity. Andrology 2016, 4, 189–212. [Google Scholar] [CrossRef]
- Oatley, J.M.; Brinster, R.L. The germline stem cell niche unit in mammalian testes. Physiol. Rev. 2012, 92, 577–595. [Google Scholar] [CrossRef]
- Wu, S.; Yan, M.; Ge, R.; Cheng, C.Y. Crosstalk between Sertoli and Germ Cells in Male Fertility. Trends Mol. Med. 2020, 26, 215–231. [Google Scholar] [CrossRef] [PubMed]
- Chojnacka, K.; Zarzycka, M.; Mruk, D.D. Biology of the Sertoli Cell in the Fetal, Pubertal, and Adult Mammalian Testis. Results Probl. Cell Differ. 2016, 58, 225–251. [Google Scholar] [CrossRef] [PubMed]
- Luaces, J.P.; Toro-Urrego, N.; Otero-Losada, M.; Capani, F. What do we know about blood-testis barrier? current understanding of its structure and physiology. Front. Cell Dev. Biol. 2023, 11, 1114769. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; You, X.; Zhu, K.; Zhao, X.; Yuan, D.; Wang, T.; Dun, Y.; Wu, J.; Ren, D.; Zhang, C.; et al. Changes in the tight junctions of the testis during aging: Role of the p38 MAPK/MMP9 pathway and autophagy in Sertoli cells. Exp. Gerontol. 2022, 161, 111729. [Google Scholar] [CrossRef] [PubMed]
- Ning, W.; Liao, X.; Dong, X.; Wang, Y.; Yang, X.; Xu, J.; Yi, S.; Yang, Z. Protective effect of astragaloside IV on cadmium-induced spermatogenesis microenvironment damage in rats. Syst. Biol. Reprod. Med. 2022, 68, 203–212. [Google Scholar] [CrossRef]
- Dolati, P.; Khodabandeh, Z.; Zamiri, M.J.; Jamhiri, I.; Mehrabani, D. The Effect of Lead Acetate and Quercetin on the Tight and Gap Junctions in the Mouse Testis. Biol. Trace Elem. Res. 2020, 198, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shen, S.; Ruan, X.; Liu, X.; Zheng, J.; Liu, Y.; Yang, C.; Wang, D.; Liu, L.; Ma, J.; et al. Biosynthetic CircRNA_001160 induced by PTBP1 regulates the permeability of BTB via the CircRNA_001160/miR-195-5p/ETV1 axis. Cell Death Dis. 2019, 10, 960. [Google Scholar] [CrossRef]
- Stammler, A.; Lüftner, B.U.; Kliesch, S.; Weidner, W.; Bergmann, M.; Middendorff, R.; Konrad, L. Highly Conserved Testicular Localization of Claudin-11 in Normal and Impaired Spermatogenesis. PLoS ONE 2016, 11, e0160349. [Google Scholar] [CrossRef]
- González-Muñoz, E.; López-Iglesias, C.; Calvo, M.; Palacín, M.; Zorzano, A.; Camps, M. Caveolin-1 loss of function accelerates glucose transporter 4 and insulin receptor degradation in 3T3-L1 adipocytes. Endocrinology 2009, 150, 3493–3502. [Google Scholar] [CrossRef]
- Yun, J.H.; Park, S.J.; Jo, A.; Kang, J.L.; Jou, I.; Park, J.S.; Choi, Y.H. Caveolin-1 is involved in reactive oxygen species-induced SHP-2 activation in astrocytes. Exp. Mol. Med. 2011, 43, 660–668. [Google Scholar] [CrossRef]
- Liu, P.; Rudick, M.; Anderson, R.G. Multiple functions of caveolin-1. J. Biol. Chem. 2002, 277, 41295–41298. [Google Scholar] [CrossRef] [PubMed]
- Quest, A.F.; Leyton, L.; Párraga, M. Caveolins, caveolae, and lipid rafts in cellular transport, signaling, and disease. Biochem. Cell Biol. 2004, 82, 129–144. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Ghosh, S.; Wang, Z.; Hunter, T. Downregulation of caveolin-1 function by EGF leads to the loss of E-cadherin, increased transcriptional activity of beta-catenin, and enhanced tumor cell invasion. Cancer Cell 2003, 4, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Nag, S.; Manias, J.L.; Kapadia, A.; Stewart, D.J. Molecular Changes Associated with the Protective Effects of Angiopoietin-1 During Blood-Brain Barrier Breakdown Post-Injury. Mol. Neurobiol. 2017, 54, 4232–4242. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Wang, X.; Wan, B.; Wang, D.; Li, M.; Cheng, K.; Luo, Q.; Wang, D.; Lu, Y.; Zhu, L. Caveolin-1 accelerates hypoxia-induced endothelial dysfunction in high-altitude cerebral edema. Cell Commun. Signal. CCS 2022, 20, 160. [Google Scholar] [CrossRef] [PubMed]
- Roy-Luzarraga, M.; Hodivala-Dilke, K. Molecular Pathways: Endothelial Cell FAK-A Target for Cancer Treatment. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2016, 22, 3718–3724. [Google Scholar] [CrossRef] [PubMed]
- Kleinschmidt, E.G.; Schlaepfer, D.D. Focal adhesion kinase signaling in unexpected places. Curr. Opin. Cell Biol. 2017, 45, 24–30. [Google Scholar] [CrossRef]
- Li, S.Y.; Mruk, D.D.; Cheng, C.Y. Focal adhesion kinase is a regulator of F-actin dynamics: New insights from studies in the testis. Spermatogenesis 2013, 3, e25385. [Google Scholar] [CrossRef]
- Siu, M.K.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Adhering junction dynamics in the testis are regulated by an interplay of beta 1-integrin and focal adhesion complex-associated proteins. Endocrinology 2003, 144, 2141–2163. [Google Scholar] [CrossRef]
- Smith, B.E.; Braun, R.E. Germ cell migration across Sertoli cell tight junctions. Science 2012, 338, 798–802. [Google Scholar] [CrossRef]
- Wong, C.H.; Xia, W.; Lee, N.P.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Regulation of ectoplasmic specialization dynamics in the seminiferous epithelium by focal adhesion-associated proteins in testosterone-suppressed rat testes. Endocrinology 2005, 146, 1192–1204. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Breton, S. The MAPK/ERK-Signaling Pathway Regulates the Expression and Distribution of Tight Junction Proteins in the Mouse Proximal Epididymis. Biol. Reprod. 2016, 94, 22. [Google Scholar] [CrossRef] [PubMed]
- Chiba, K.; Kondo, Y.; Yamaguchi, K.; Miyake, H.; Fujisawa, M. Inhibition of claudin-11 and occludin expression in rat Sertoli cells by mono-(2-ethylhexyl) phthalate through p44/42 mitogen-activated protein kinase pathway. J. Androl. 2012, 33, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.L.; Chen, L.; Hess, R.A.; Müller, R.; Gonzalez, F.J.; Peters, J.M. Peroxisome Proliferator-activated Receptor-D (PPARD) Coordinates Mouse Spermatogenesis by Modulating Extracellular Signal-regulated Kinase (ERK)-dependent Signaling. J. Biol. Chem. 2015, 290, 23416–23431. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yin, Y.; Wang, G.; Liu, Z.; Liu, L.; Sun, F. Interleukin-6 disrupts blood-testis barrier through inhibiting protein degradation or activating phosphorylated ERK in Sertoli cells. Sci. Rep. 2014, 4, 4260. [Google Scholar] [CrossRef]
- Jiang, X.; Zhu, C.; Li, X.; Sun, J.; Tian, L.; Bai, W. Cyanidin-3- O-glucoside at Low Doses Protected against 3-Chloro-1,2-propanediol Induced Testis Injury and Improved Spermatogenesis in Male Rats. J. Agric. Food Chem. 2018, 66, 12675–12684. [Google Scholar] [CrossRef]
- Akar, F.; Yildirim, O.G.; Yucel Tenekeci, G.; Tunc, A.S.; Demirel, M.A.; Sadi, G. Dietary high-fructose reduces barrier proteins and activates mitogenic signalling in the testis of a rat model: Regulatory effects of kefir supplementation. Andrologia 2022, 54, e14342. [Google Scholar] [CrossRef]
- Wong, C.H.; Cheng, C.Y. Mitogen-activated protein kinases, adherens junction dynamics, and spermatogenesis: A review of recent data. Dev. Biol. 2005, 286, 1–15. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene Sequence (5′–3′) | Tm (°C) |
|---|---|---|
| ZO-1 | F: GAGGACAGTTACGACGAGG | 59 |
| R: CAGGGTGACTTTGGTGGG | ||
| Occludin | F: GAACGAGAAGCGACCGTATCC | 59 |
| R: CTTGCTCTGCCCGCCTTG | ||
| CAV1 | F: GCAACATCTACAAGCCCAACA | 56 |
| R: GCAGACAGCAAACGGTAAAAC | ||
| SRC | F: CCTCAGGCATGGCTTATGT | 54 |
| R: ACCCGTCCCTTTGTTGTG | ||
| FAK | F: GACAGTTACAACGAGGGCGTCAA | 58 |
| R: GGCGGGCAGAACAGGAATG | ||
| ERK1 | F: CAACACCACCTGCGACCTTA | 56 |
| R: GGGATGGGGAGCCTAGAATA | ||
| TGFB1 | F: GGTGGAATACGGCAACAAA | 55 |
| R: GTGGGCACTGAGGCGAAA | ||
| HOXD10 | F: TGGACAGACCCGAACAGAT | 55 |
| R: GATGGGACCTCAGCAGAAAT | ||
| LCP2 | F: CAAGTCCAGCGGTTTCC | 55 |
| R: GGGTGGTGGCTCGTAAT | ||
| GAPDH | F: GCTGGTGCTGAGTATGTGGTG | 58 |
| R: GCTGACAATCTTGAGGGTGTTG | ||
| CD27 | F: GGGAGGATGGTGTTGTCAGA | 56 |
| R: TTCAGCATAAGGTAAGTGGGAG | ||
| JNK1 | F: GTTGACATTTGGTCAGTTGGG | 58 |
| R: GGGAAAAGTACATCAGGGAAGA | ||
| ITGAM | F: AGTCTGCCTCCAAGTTCGC | 55 |
| R: TTCAGGGTCTCGCATTTCT | ||
| TLR4 | F: CTGCCTTCACTACAGGGACTT | 56 |
| R: TGGGACACCACGACAATAAC | ||
| LPXN | F: CCCTTCCGTTCCTGATGACA | 56 |
| R: TGCGGCTGCTGAGGTTTTA | ||
| Claudin | F: TATGGCTACGGGGCTTTAC | 57 |
| R: AACTGTGAGCAGCAGGAGAAT | ||
| miR-150-x | F: AGTGGTCGTATCCAGTGCG | 60 |
| miR-1247-x | F: CCCGGAGTCGTATCCAGTG | 57 |
| miR-2713-z | F: CGGGCATCATTGAAGGTC | 60 |
| miR-12308-y | F: TAAATCCCTGCTCTGACACG | 56 |
| miR-2432-z | F: TAGAGGTCGTCGTATCCAGTG | 58 |
| miR-2431-y | F: GTGCGAATACCTCGGACC | 58 |
| novel-m0399-3p | F: GAGCGTCGTATCCAGTGCG | 62 |
| novel-m0230-3p | F: AGCGTGAGCAGGAGCAGC | 60 |
| U6 | R: CGAGGATGTGAAGACACCAAGAC | 59 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Q.; Li, T.; Zhang, Y.; Zhao, X.; Wang, Q.; Yuan, L. Caveolin 1 Regulates the Tight Junctions between Sertoli Cells and Promotes the Integrity of Blood–Testis Barrier in Yak via the FAK/ERK Signaling Pathway. Animals 2024, 14, 183. https://doi.org/10.3390/ani14020183
Yan Q, Li T, Zhang Y, Zhao X, Wang Q, Yuan L. Caveolin 1 Regulates the Tight Junctions between Sertoli Cells and Promotes the Integrity of Blood–Testis Barrier in Yak via the FAK/ERK Signaling Pathway. Animals. 2024; 14(2):183. https://doi.org/10.3390/ani14020183
Chicago/Turabian StyleYan, Qiu, Tianan Li, Yong Zhang, Xingxu Zhao, Qi Wang, and Ligang Yuan. 2024. "Caveolin 1 Regulates the Tight Junctions between Sertoli Cells and Promotes the Integrity of Blood–Testis Barrier in Yak via the FAK/ERK Signaling Pathway" Animals 14, no. 2: 183. https://doi.org/10.3390/ani14020183
APA StyleYan, Q., Li, T., Zhang, Y., Zhao, X., Wang, Q., & Yuan, L. (2024). Caveolin 1 Regulates the Tight Junctions between Sertoli Cells and Promotes the Integrity of Blood–Testis Barrier in Yak via the FAK/ERK Signaling Pathway. Animals, 14(2), 183. https://doi.org/10.3390/ani14020183