
Impact of Perch Provision Timing on Activity and Musculoskeletal Health of Laying Hens

 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Animals and Housing
2.3. Treatments
2.4. Activity
2.5. Musculoskeletal Health
2.5.1. Computed Tomography (CT) Image Acquisition
2.5.2. Tibiotarsal CT Image Analyses
2.5.3. Muscle Deposition
2.5.4. Tibia Breaking Strength
2.5.5. Tibia Ash Percentage
2.5.6. Bone Resorption Markers
2.5.7. Data Processing and Statistical Analysis
3. Results
3.1. Activity
3.2. Tibial Bone Mineral Density (BMD) and Cross-Sectional Area (CSA)
3.3. Muscle Deposition
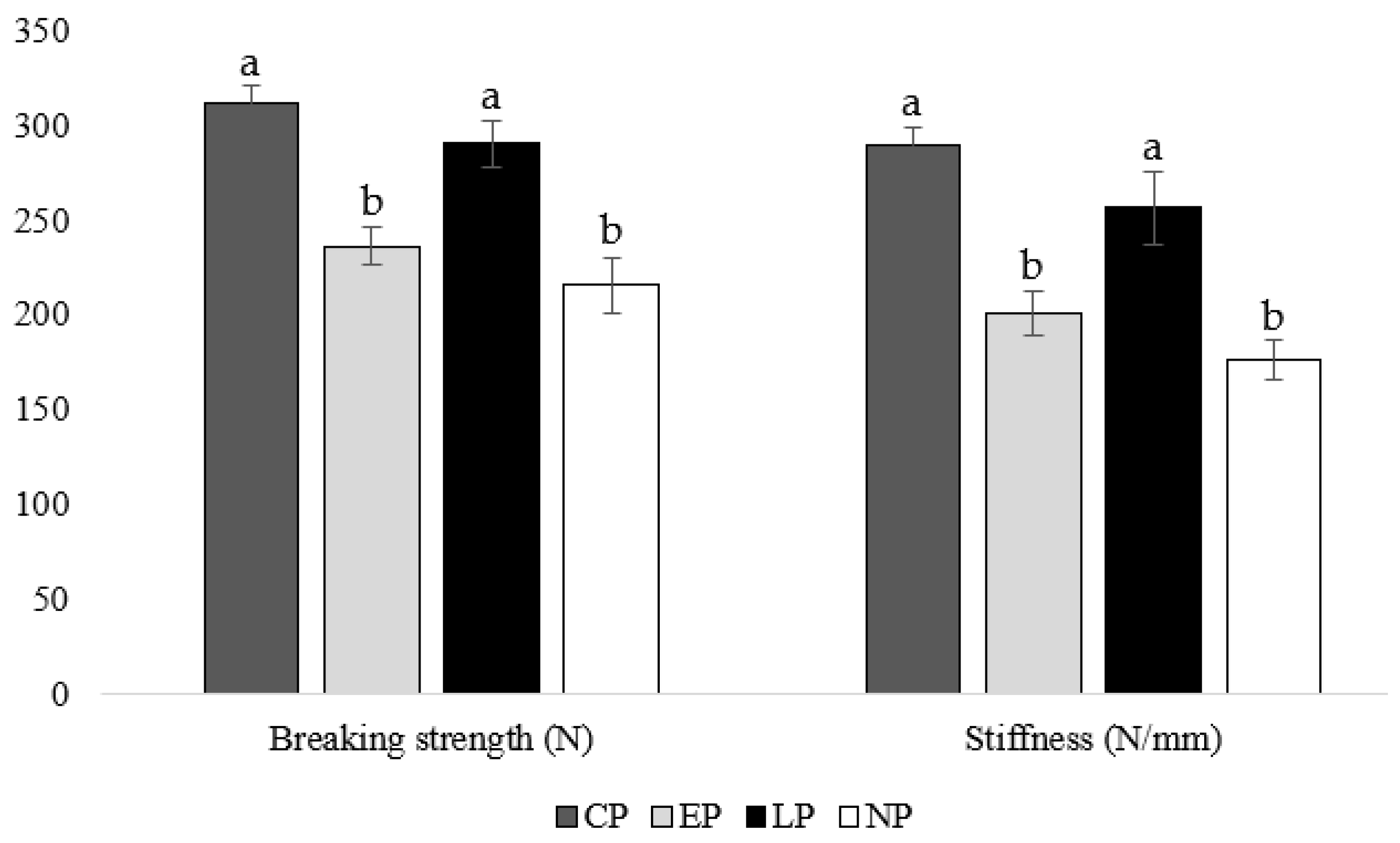
3.4. Tibia Breaking Strength
3.5. Tibia Ash Percentage
3.6. Bone Demineralization
4. Discussion
4.1. Activity
4.2. Tibial Bone Mineral Density (BMD) and Cross-Sectional Area (CSA)
4.3. Muscle Deposition
4.4. Tibia Breaking Strength
4.5. Tibia Ash Percentage
4.6. Bone Resorption
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whitehead, C.C. Overview of Bone Biology in the Egg-Laying Hen. Poult. Sci. 2004, 83, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, C.C.; Fleming, R.H. Osteoporosis in Cage Layers. Poult. Sci. 2000, 79, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Nasr, M.; Murrell, J.; Wilkins, L.; Nicol, C. The Effect of Keel Fractures on Egg-Production Parameters, Mobility and Behaviour in Individual Laying Hens. Anim. Welf. 2012, 21, 127–135. [Google Scholar] [CrossRef]
- Nasr, M.A.F.; Nicol, C.J.; Murrell, J.C. Do Laying Hens with Keel Bone Fractures Experience Pain? PLoS ONE 2012, 7, e42420. [Google Scholar] [CrossRef] [PubMed]
- Cufadar, Y.; Olgun, O.; Yildiz, A.Ö. The Effect of Dietary Calcium Concentration and Particle Size on Performance, Eggshell Quality, Bone Mechanical Properties and Tibia Mineral Contents in Moulted Laying Hens. Br. Poult. Sci. 2011, 52, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Saunders-Blades, J.L.; MacIsaac, J.L.; Korver, D.R.; Anderson, D.M. The Effect of Calcium Source and Particle Size on the Production Performance and Bone Quality of Laying Hens. Poult. Sci. 2009, 88, 338–353. [Google Scholar] [CrossRef]
- Koutoulis, K.C.; Kyriazakis, I.; Perry, G.C.; Lewis, P.D. Effect of Different Calcium Sources and Calcium Intake on Shell Quality and Bone Characteristics of Laying Hens at Sexual Maturity and End of Lay. Int. J. Poult. Sci. 2009, 8, 342–348. [Google Scholar] [CrossRef]
- Fleming, R.H.; McCormack, H.A.; Whitehead, C.C. Bone Structure and Strength at Different Ages in Laying Hens and Effects of Dietary Particulate Limestone, Vitamin K and Ascorbic Acid. Br. Poult. Sci. 1998, 39, 434–440. [Google Scholar] [CrossRef]
- Rennie, J.S.; Fleming, R.H.; McCormack, H.A.; McCorquodale, C.C.; Whitehead, C.C. Studies on Effects of Nutritional Factors on Bone Structure and Osteoporosis in Laying Hens. Br. Poult. Sci. 1997, 38, 417–424. [Google Scholar] [CrossRef]
- Bishop, S.C.; Fleming, R.H.; McCormack, H.A.; Flock, D.K.; Whitehead, C.C. Inheritance of Bone Characteristics Affecting Osteoporosis in Laying Hens. Br. Poult. Sci. 2000, 41, 33–40. [Google Scholar] [CrossRef]
- Hocking, P.M.; Bain, M.; Channing, C.E.; Fleming, R.; Wilson, S. Genetic Variation for Egg Production, Egg Quality and Bone Strength in Selected and Traditional Breeds of Laying Fowl. Br. Poult. Sci. 2003, 44, 365–373. [Google Scholar] [CrossRef]
- Fleming, R.H.; McCormack, H.A.; McTeir, L.; Whitehead, C.C. Relationships between Genetic, Environmental and Nutritional Factors Influencing Osteoporosis in Laying Hens. Br. Poult. Sci. 2006, 47, 742–755. [Google Scholar] [CrossRef]
- Podisi, B.K.; Knott, S.A.; Dunn, I.C.; Burt, D.W.; Hocking, P.M. Bone Mineral Density QTL at Sexual Maturity and End of Lay. Br. Poult. Sci. 2012, 53, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Enneking, S.A.; Cheng, H.W.; Jefferson-Moore, K.Y.; Einstein, M.E.; Rubin, D.A.; Hester, P.Y. Early Access to Perches in Caged White Leghorn Pullets. Poult. Sci. 2012, 91, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Hester, P.Y.; Enneking, S.A.; Haley, B.K.; Cheng, H.W.; Einstein, M.E.; Rubin, D.A. The Effect of Perch Availability during Pullet Rearing and Egg Laying on Musculoskeletal Health of Caged White Leghorn Hens. Poult. Sci. 2013, 92, 1972–1980. [Google Scholar] [CrossRef] [PubMed]
- Jendral, M.J.; Korver, D.R.; Church, J.S.; Feddes, J.J.R. Bone Mineral Density and Breaking Strength of White Leghorns Housed in Conventional, Modified, and Commercially Available Colony Battery Cages. Poult. Sci. 2008, 87, 828–837. [Google Scholar] [CrossRef]
- Gunnarsson, S.; Yngvesson, J.; Keeling, L.J.; Forkman, B. Rearing without Early Access to Perches Impairs the Spatial Skills of Laying Hens. Appl. Anim. Behav. Sci. 2000, 67, 217–228. [Google Scholar] [CrossRef]
- Casey-Trott, T.M.; Korver, D.R.; Guerin, M.T.; Sandilands, V.; Torrey, S.; Widowski, T.M. Opportunities for Exercise during Pullet Rearing, Part I: Effect on the Musculoskeletal Characteristics of Pullets. Poult. Sci. 2017, 96, 2509–2517. [Google Scholar] [CrossRef]
- Casey-Trott, T.M.; Korver, D.R.; Guerin, M.T.; Sandilands, V.; Torrey, S.; Widowski, T.M. Opportunities for Exercise during Pullet Rearing, Part II: Long-Term Effects on Bone Characteristics of Adult Laying Hens at the End-of-Lay. Poult. Sci. 2017, 96, 2518–2527. [Google Scholar] [CrossRef]
- Wood-Gush, D.G.M. The Behaviour of the Domestic Fowl; Heinemann Educational Books Ltd.: Portsmouth, NW, USA, 1971; ISBN 9780435. [Google Scholar]
- Olsson, I.A.S.; Keeling, L.J. Night-Time Roosting in Laying Hens and the Effect of Thwarting Access to Perches. Appl. Anim. Behav. Sci. 2000, 68, 243–256. [Google Scholar] [CrossRef]
- Olsson, I.A.S.A.S.; Keeling, L.J.J. The Push-Door for Measuring Motivation in Hens: Laying Hens Are Motivated to Perch at Night. Anim. Welf. 2002, 11, 11–19. [Google Scholar] [CrossRef]
- Wilkins, L.J.; Brown, S.N.; Zimmerman, P.H.; Leeb, C.; Nicol, C.J. Investigation of Palpation as a Method for Determining the Prevalence of Keel and Furculum Damage in Laying Hens. Vet. Rec. 2004, 155, 547–549. [Google Scholar] [CrossRef]
- Harlander-Matauschek, A.; Rodenburg, T.B.; Sandilands, V.; Tobalske, B.W.; Toscano, M.J. Causes of Keel Bone Damage and Their Solutions in Laying Hens. Worlds Poult. Sci. J. 2015, 71, 461–472. [Google Scholar] [CrossRef]
- Whitehead, C.C. Bone Biology and Skeletal Disorders in Poultry; Poultry Science Symposium Series; Carfax Publishing Co.: Centreville, VA, USA, 1992. [Google Scholar]
- Moinard, C.; Statham, P.; Haskell, M.J.; McCorquodale, C.; Jones, R.B.; Green, P.R. Accuracy of Laying Hens in Jumping Upwards and Downwards between Perches in Different Light Environments. Appl. Anim. Behav. Sci. 2004, 85, 77–92. [Google Scholar] [CrossRef]
- Appleby, M.C.; Duncan, I.J.H. Development of Perching in Hens. Biol. Behav. 1989, 14, 157–168. [Google Scholar]
- Regmi, P.; Deland, T.S.; Steibel, J.P.; Robison, C.I.; Haut, R.C.; Orth, M.W.; Karcher, D.M. Effect of Rearing Environment on Bone Growth of Pullets. Poult. Sci. 2015, 94, 502–511. [Google Scholar] [CrossRef]
- Regmi, P.; Nelson, N.; Steibel, J.P.; Anderson, K.E.; Karcher, D.M. Comparisons of Bone Properties and Keel Deformities between Strains and Housing Systems in End-of-Lay Hens. Poult. Sci. 2016, 95, 2225–2234. [Google Scholar] [CrossRef] [PubMed]
- Casey-Trott, T.M.; Guerin, M.T.; Sandilands, V.; Torrey, S.; Widowski, T.M. Rearing System Affects Prevalence of Keel-Bone Damage in Laying Hens: A Longitudinal Study of Four Consecutive Flocks. Poult. Sci. 2017, 96, 2029–2039. [Google Scholar] [CrossRef]
- Vits, A.; Weitzenbürger, D.; Hamann, H.; Distl, O. Production, Egg Quality, Bone Strength, Claw Length, and Keel Bone Deformities of Laying Hens Housed in Furnished Cages with Different Group Sizes. Poult. Sci. 2005, 84, 1511–1519. [Google Scholar] [CrossRef]
- Hy-Line Brown. Hy-Line Management Guide. Available online: https://www.hyline.com/filesimages/Hy-Line-Products/Hy-Line-Product-PDFs/Brown/BRN%20COM%20ENG.pdf (accessed on 25 September 2023).
- Harrison, C.; Jones, J.; Bridges, W.; Anderson, G.; Ali, A.; Mercuri, J. Associations among Computed Tomographic Measures of Bone and Muscle Quality and Biomechanical Measures of Tibiotarsal Bone Quality in Laying Hens. Am. J. Vet. Res. 2023, 84, 1–9. [Google Scholar] [CrossRef]
- ANSI/ASAE S459 MAR1992 (R2017). Shear and Three-Point Bending Test of Animal Bone. Available online: https://webstore.ansi.org/preview-pages/ASABE/preview_ANSI+ASAE+S459+MAR1992+(R2017).pdf (accessed on 25 September 2023).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. J. Math. Methods Biosci. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Brantsæter, M.; Nordgreen, J.; Rodenburg, T.B.; Tahamtani, F.M.; Popova, A.; Janczak, A.M. Exposure to Increased Environmental Complexity during Rearing Reduces Fearfulness and Increases Use of Three-Dimensional Space in Laying Hens (Gallus gallus domesticus). Front. Vet. Sci. 2016, 3, 14. [Google Scholar] [CrossRef]
- Tactacan, G.B.; Guenter, W.; Lewis, N.J.; Rodriguez-Lecompte, J.C.; House, J.D. Performance and Welfare of Laying Hens in Conventional and Enriched Cages. Poult. Sci. 2009, 88, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Regmi, P.; Smith, N.; Nelson, N.; Haut, R.C.; Orth, M.W.; Karcher, D.M. Housing Conditions Alter Properties of the Tibia and Humerus during the Laying Phase in Lohmann White Leghorn Hens. Poult. Sci. 2016, 95, 198–206. [Google Scholar] [CrossRef]
- Hester, P.Y.; Enneking, S.A.; Jefferson-Moore, K.Y.; Einstein, M.E.; Cheng, H.W.; Rubin, D.A. The Effect of Perches in Cages during Pullet Rearing and Egg Laying on Hen Performance, Foot Health, and Plumage. Poult. Sci. 2013, 92, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Moinard, C.; Morisse, J.P.; Faure, J.M. Effect of Cage Area, Cage Height and Perches on Feather Condition, Bone Breakage and Mortality of Laying Hens. Br Poult. Sci. 1998, 39, 198–202. [Google Scholar] [CrossRef]
- Widowski, T.M.; Caston, L.J.; Hunniford, M.E.; Cooley, L.; Torrey, S. Effect of Space Allowance and Cage Size on Laying Hens Housed in Furnished Cages, Part I: Performance and Well-Being. Poult. Sci. 2017, 96, 3805–3815. [Google Scholar] [CrossRef]
- Sharma, M.K.; White, D.; Chen, C.; Kim, W.K.; Adhikari, P. Effects of the Housing Environment and Laying Hen Strain on Tibia and Femur Bone Properties of Different Laying Phases of Hy-Line Hens. Poult. Sci. 2021, 100, 100933. [Google Scholar] [CrossRef]
- Silversides, F.G.; Singh, R.; Cheng, K.M.; Korver, D.R. Comparison of Bones of 4 Strains of Laying Hens Kept in Conventional Cages and Floor Pens. Poult. Sci. 2012, 91, 1–7. [Google Scholar] [CrossRef]
- Yilmaz Dikmen, B.; İpek, A.; Şahan, Ü.; Petek, M.; Sözcü, A. Egg Production and Welfare of Laying Hens Kept in Different Housing Systems (Conventional, Enriched Cage, and Free Range). Poult. Sci. 2016, 95, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Neijat, M.; Casey-Trott, T.M.; Robinson, S.; Widowski, T.M.; Kiarie, E. Effects of Rearing and Adult Laying Housing Systems on Medullary, Pneumatic and Radius Bone Attributes in 73-Wk Old Lohmann LSL Lite Hens. Poult. Sci. 2019, 98, 2840–2845. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Week | Treatment | Overall Activity (g) | Vertical Activity (g) | Horizontal Activity (g) | Average Daily Vertical Displacement/Bird (F) |
|---|---|---|---|---|---|
| 24 | CP | 1.42 ± 0.11 a | 0.59 ± 0.06 a | 0.83 ± 0.09 a | 24.52 ± 2.96 a |
| EP | 1.33 ± 0.13 a | 0.18 ± 0.01 b | 1.15 ± 0.11 b | 3.69 ± 0.69 b | |
| LP | 1.39 ± 0.19 a | 0.51 ± 0.07 a | 0.88 ± 0.07 a | 23.58 ± 4.58 a | |
| NP | 1.29 ± 0.21 b | 0.11 ± 0.03 b | 1.18 ± 0.06 b | 1.13 ± 0.96 b | |
| p-value | 0.034 | 0.029 | 0.022 | 0.031 | |
| 36 | CP | 1.44 ± 0.16 a | 0.56 ± 0.06 a | 0.88 ± 0.11 a | 33.25 ± 4.21 a |
| EP | 1.45 ± 0.21 a | 0.12 ± 0.03 b | 1.33 ± 0.16 b | 3.56 ± 1.25 b | |
| LP | 1.41 ± 0.19 a | 0.55 ± 0.09 a | 0.86 ± 0.09 a | 29.87 ± 5.25 a | |
| NP | 1.31 ± 0.21 a | 0.11 ± 0.01 b | 1.20 ± 0.16 b | 2.03 ± 1.03 b | |
| p-value | 0.096 | 0.032 | 0.036 | 0.028 | |
| 40 | CP | 1.35 ± 0.22 a | 0.53 ± 0.07 a | 0.82 ± 0.10 a | 28.85 ± 6.69 a |
| EP | 1.39 ± 0.23 a | 0.13 ± 0.06 b | 1.26 ± 0.09 b | 4.03 ± 2.36 b | |
| LP | 1.36 ± 0.29 a | 0.59 ± 0.03 a | 0.77 ± 0.06 a | 26.85 ± 4.52 a | |
| NP | 1.32 ± 0.27 a | 0.09 ± 0.01 b | 1.23 ± 0.17 b | 1.63 ± 0.85 b | |
| p-value | 0.325 | 0.031 | 0.029 | 0.035 |
| Parameter/ Treatment | Bone Cross-Sectional Area (mm2) | |||||
| Total | Cortical | |||||
| Proximal | Middle | Distal | Proximal | Middle | Distal | |
| CP | 70.2 ± 1.1 a | 53.8 ± 1.3 a | 55.5 ± 0.7 a | 37.1 ± 2.4 a | 29.5 ± 2.1 a | 29.2 ± 1.8 a |
| EP | 70.1 ± 1.3 a | 53.9 ± 1.0 a | 55.3 ± 0.6 a | 30.7 ± 2.1 b | 23.9 ± 1.6 b | 24.0 ± 1.5 b |
| LP | 69.9 ± 1.1 a | 54.5 ± 0.8 a | 56.0 ± 0.7 a | 31.5 ± 2.1 b | 25.4 ± 1.6 b | 24.2 ± 1.5 b |
| NP | 69.9 ± 1.1 a | 54.9 ± 0.7 a | 56.0 ± 1.1 a | 26.3 ± 1.7 c | 21.6 ± 1.4 c | 20.9 ± 1.4 b |
| p-value | 0.235 | 0.185 | 0.635 | 0.021 | 0.019 | 0.024 |
| Parameter/ Treatment | Bone Mineral Density (mg/cm3) | |||||
| Total | Cortical | |||||
| Proximal | Middle | Distal | Proximal | Middle | Distal | |
| CP | 515.7 ± 13.7 a | 730.6 ± 11.0 a | 806.8 ± 11.0 a | 1028.8 ± 23.6 a | 1746.8 ± 16.9 a | 1370.7 ± 889.5 a |
| EP | 428.1 ± 15.4 a | 591.8 ± 22.5 b | 661.6 ± 14.6 b | 740.7 ± 20.9 b | 1257.7 ± 17.9 b | 1000.6 ± 22.0 b |
| LP | 438.4 ± 14.0 a | 628.3 ± 18.8 b | 669.7 ± 17.4 b | 761.3 ± 21.0 b | 1380.0 ± 22.6 b | 1165.1 ± 19.6 b |
| NP | 356.3 ± 11.4 b | 502.6 ± 13.1 c | 545.8 ± 20.7 c | 726.9 ± 23.3 c | 976.5 ± 19.1 c | 889.5 ± 23.1 c |
| p-value | 0.034 | 0.003 | 0.021 | 0.013 | 0.025 | 0.022 |
| Treatment | Biceps Brachii (g) | Triceps Brachii (g) | Pectoralis Major (g) | Pectoralis Minor (g) | Leg Muscle Group (g) |
|---|---|---|---|---|---|
| CP | 4.25 ± 0.26 a | 3.88 ± 0.31 a | 124.58 ± 4.85 a | 62.58 ± 5.55 a | 141.85 ± 7.98 a |
| EP | 3.65 ± 0.29 b | 3.59 ± 0.22 a | 112.55 ± 5.25 b | 56.85 ± 1.14 a | 124.55 ± 6.52 b |
| LP | 4.18 ± 0.38 a | 3.67 ± 0.29 a | 119.93 ± 4.99 a | 59.22 ± 3.55 a | 138.57 ± 5.88 a |
| NP | 3.52 ± 0.21 b | 3.53 ± 0.25 a | 107.58 ± 3.78 b | 55.85 ± 5.03 a | 120.79 ± 6.85 b |
| p-value | 0.026 | 0.259 | 0.031 | 0.523 | 0.028 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anderson, M.G.; Johnson, A.M.; Harrison, C.; Arguelles-Ramos, M.; Ali, A. Impact of Perch Provision Timing on Activity and Musculoskeletal Health of Laying Hens. Animals 2024, 14, 265. https://doi.org/10.3390/ani14020265
Anderson MG, Johnson AM, Harrison C, Arguelles-Ramos M, Ali A. Impact of Perch Provision Timing on Activity and Musculoskeletal Health of Laying Hens. Animals. 2024; 14(2):265. https://doi.org/10.3390/ani14020265
Chicago/Turabian StyleAnderson, Mallory G., Alexa M. Johnson, Cerano Harrison, Mireille Arguelles-Ramos, and Ahmed Ali. 2024. "Impact of Perch Provision Timing on Activity and Musculoskeletal Health of Laying Hens" Animals 14, no. 2: 265. https://doi.org/10.3390/ani14020265
APA StyleAnderson, M. G., Johnson, A. M., Harrison, C., Arguelles-Ramos, M., & Ali, A. (2024). Impact of Perch Provision Timing on Activity and Musculoskeletal Health of Laying Hens. Animals, 14(2), 265. https://doi.org/10.3390/ani14020265