
Physiological and Genetic Basis of High-Altitude Indigenous Animals’ Adaptation to Hypoxic Environments


Simple Summary
Abstract
1. Introduction
2. High-Altitude Hypoxic Adaptation and Physiological Biochemical Characteristics
2.1. Pulmonary and Cardiovascular System Characteristics
2.2. Characteristics of O2-Consuming Tissues
2.3. Oxidative Stress
2.4. Other Systems Related to Adaptation to High-Altitude Hypoxic Environments
3. Genetics Study of Hypoxia Adaptation at High Altitude
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- West, J.B.; Schoene, R.B.; Milledge, J.S. High Altitude Medicine and Physiology; Hodder Arnold: London, UK, 2007; p. 27. [Google Scholar]
- Niermeyer, S.; Zamdio, S.; Moore, L.G. The People. In High Altitude: An Exploration of Human Adaptation; Marcel Dekker: New York, NY, USA, 2001; pp. 42–100. [Google Scholar]
- Beall, C.M. Human adaptability studies at high altitude: Research designs and major concepts during fifty years of discovery. Am. J. Hum. Biol. 2013, 25, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Gujar, G.; Shashank, C.; Ponsuksili, S. Selection signatures for high altitude adaptation in livestock: A review. Gene 2024, 927, 148757. [Google Scholar] [CrossRef] [PubMed]
- Monge, C.; Leon-Velarde, F. Physiological adaptation to high altitude: Oxygen transport in mammals and birds. Physiol. Rev. 1991, 71, 1135–1172. [Google Scholar] [CrossRef] [PubMed]
- Monge Medrano, C. La Enfermedad de los Andes. Fac. Med. 1928, 11, 90–91. [Google Scholar]
- Murillo, J.P. Between the acclimatization to high altitude, the medical anthropology and the civilizing utopia. Mapping of the evolution of thought of Carlos Monge Medrano on the health-illness process of andean populations. Rev. Peru. Med. Exp. Salud Publica 2017, 34, 280–286. [Google Scholar] [CrossRef]
- Barcroft, J. Phases in Foetal Life. Glasgow Med. J. 1943, 139, 1–13. [Google Scholar]
- Snyder, L.R. Low P50 in deer mice native to high altitude. J. Appl. Physiol. 1985, 58, 193–199. [Google Scholar] [CrossRef]
- Signore, A.V.; Storz, J.F. Biochemical pedomorphosis and genetic assimilation in the hypoxia adaptation of Tibetan antelope. Sci. Adv. 2020, 6, b5447. [Google Scholar] [CrossRef]
- Chiou, K.L.; Janiak, M.C.; Schneider-Crease, I.A.; Sen, S.; Ayele, F.; Chuma, I.S.; Knauf, S.; Lemma, A.; Signore, A.V.; D’ippolito, A.M.; et al. Genomic signatures of high-altitude adaptation and chromosomal polymorphism in geladas. Nat. Ecol. Evol. 2022, 6, 630–643. [Google Scholar] [CrossRef]
- Gao, X.; Wang, S.; Wang, Y.-F.; Li, S.; Wu, S.-X.; Yan, R.-G.; Zhang, Y.-W.; Wan, R.-D.; He, Z.; Song, R.-D.; et al. Long read genome assemblies complemented by single cell RNA-sequencing reveal genetic and cellular mechanisms underlying the adaptive evolution of yak. Nat. Commun. 2022, 13, 4887. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Li, Y.; Pan, J.; Wang, D.; Chen, W.; Zheng, Z.; He, X.; Zhao, Q.; Pu, Y.; et al. EPAS1 gain-of-function mutation contributes to high-altitude adaptation in Tibetan horses. Mol. Biol. Evol. 2019, 36, 2591–2603. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Wang, H.; Liu, G.; Zhao, F.; Kijas, J.W.; Ma, Y.; Lu, J.; Zhang, L.; Cao, J.; Wu, M.; et al. Genome-wide analysis reveals adaptation to high altitudes in Tibetan sheep. Sci. Rep. 2016, 6, 26770. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Tian, S.; Jin, L.; Zhou, G.; Li, Y.; Zhang, Y.; Wang, T.; Yeung, C.K.L.; Chen, L.; Ma, J.; et al. Genomic analyses identify distinct patterns of selection in domesticated pigs and Tibetan wild boars. Nat. Genet. 2013, 45, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Sodhi, M.; Sharma, M.; Sharma, V.; Mukesh, M. Hypoxia related genes modulate in similar fashion in skin fibroblast cells of yak (Bos grunniens) adapted to high altitude and native cows (Bos indicus) adapted to tropical climate during hypoxia stress. Int. J. Biometeorol. 2024, 68, 1675–1687. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Tian, D.; Li, X.; Liu, S.; Tian, F.; Liu, D.; Wang, S.; Zhao, K. Multiomics Analyses Provide New Insight into Genetic Variation of Reproductive Adaptability in Tibetan Sheep. Mol. Biol. Evol. 2024, 41, msae058. [Google Scholar] [CrossRef]
- Beall, C.M. Adaptation to high altitude: Phenotypes and genotypes. Annu. Rev. Anthropol. 2014, 43, 251–272. [Google Scholar] [CrossRef]
- West, J.B. Physiological Effects of Chronic Hypoxia. N. Engl. J. Med. 2017, 376, 1965–1971. [Google Scholar] [CrossRef]
- Weibel, E.R. Lung morphometry: The link between structure and function. Cell Tissue Res. 2017, 367, 413–426. [Google Scholar] [CrossRef]
- Ivy, C.M.; Scott, G.R. Control of breathing and the circulation in high-altitude mammals and birds. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 186, 66–74. [Google Scholar] [CrossRef]
- Teppema, L.J.; Dahan, A. The ventilatory response to hypoxia in mammals: Mechanisms, measurement, and analysis. Physiol. Rev. 2010, 90, 675–754. [Google Scholar] [CrossRef]
- Wang, G.; He, Y.; Luo, Y. Expression of OPA1 and Mic60 genes and their association with mitochondrial cristae morphology in Tibetan sheep. Cell Tissue Res. 2019, 376, 273–279. [Google Scholar] [CrossRef]
- Ivy, C.M.; Scott, G.R. Control of breathing and ventilatory acclimatization to hypoxia in deer mice native to high altitudes. Acta Physiol. 2017, 221, 266–282. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.L.; Pranckevicius, N.A.; Nurse, C.A.; Scott, G.R. Regulation of catecholamine release from the adrenal medulla is altered in deer mice (Peromyscus maniculatus) native to high altitudes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 317, R407–R417. [Google Scholar] [CrossRef] [PubMed]
- Kiyamu, M.; Bigham, A.; Parra, E.; León-Velarde, F.; Rivera-Chira, M.; Brutsaert, T.D. Developmental and genetic components explain enhanced pulmonary volumes of female Peruvian Quechua. Am. J. Phys. Anthropol. 2012, 148, 534–542. [Google Scholar] [CrossRef]
- Penaloza, D.; Arias-Stella, J. The heart and pulmonary circulation at high altitudes: Healthy highlanders and chronic mountain sickness. Circulation 2007, 115, 1132–1146. [Google Scholar] [CrossRef] [PubMed]
- Wiener, G.; Han, J.; Long, R. The Yak, 2nd ed.; Rap Publication: Bangkok, Thailand, 2003. [Google Scholar]
- Hsia, C.C.; Carbayo, J.J.P.; Yan, X.; Bellotto, D.J. Enhanced alveolar growth and remodeling in Guinea pigs raised at high altitude. Respir. Physiol. Neurobiol. 2005, 147, 105–115. [Google Scholar] [CrossRef]
- Hsia, C.C.W.; Johnson, R.L., Jr.; McDonough, P.; Dane, D.M.; Hurst, M.D.; Fehmel, J.L.; Wagner, H.E.; Wagner, P.D. Residence at 3800-m altitude for 5 mo in growing dogs enhances lung diffusing capacity for oxygen that persists at least 2.5 years. J. Appl. Physiol. 2007, 102, 1448–1455. [Google Scholar] [CrossRef]
- Maina, J.N.; McCracken, K.G.; Chua, B.; York, J.M.; Milsom, W.K. Morphological and morphometric specializations of the lung of the Andean goose, Chloephaga melanoptera: A lifelong high-altitude resident. PLoS ONE 2017, 12, e174395. [Google Scholar] [CrossRef]
- Ding, X.; Liang, C.; Guo, X.; Wu, X.; Wang, H.; Johnson, K.; Yan, P. Physiological insight into the high-altitude adaptations in domesticated yaks (Bos grunniens) along the Qinghai-Tibetan Plateau altitudinal gradient. Livest. Sci. 2014, 162, 233–239. [Google Scholar] [CrossRef]
- West, C.M.; Wearing, O.H.; Rhem, R.G.; Scott, G.R. Pulmonary hypertension is attenuated and ventilation-perfusion matching is maintained during chronic hypoxia in deer mice native to high altitude. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 320, R800–R811. [Google Scholar] [CrossRef]
- Wei, Q.; Yu, H. Comparison of histological structure of pulmonary alveoli between 180 days old yak and plain cattle. J. Qinghai Univ. 2008, 26, 36–39. [Google Scholar]
- Maron, B.A.; Oldham, W.M.; Chan, S.Y.; Vargas, S.O.; Arons, E.; Zhang, Y.-Y.; Loscalzo, J.; Leopold, J.A. Upregulation of steroidogenic acute regulatory protein by hypoxia stimulates aldosterone synthesis in pulmonary artery endothelial cells to promote pulmonary vascular fibrosis. Circulation 2014, 130, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.R.; Ghofrani, H.-A.; Weissmann, N.; Aldashev, A.; Zhao, L. Pathophysiology and treatment of high-altitude pulmonary vascular disease. Circulation 2015, 131, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Alworth, L.C.; Harvey, S.B. Chapter 39—Anatomy, Physiology, and Behavior. In The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents; Academic Press: Boston, MA, USA, 2012; pp. 955–966. [Google Scholar]
- Wang, D.; Zhang, H.; Li, M.; Frid, M.G.; Flockton, A.R.; McKeon, B.A.; Yeager, M.E.; Fini, M.A.; Morrell, N.W.; Pullamsetti, S.S.; et al. MicroRNA-124 controls the proliferative, migratory, and inflammatory phenotype of pulmonary vascular fibroblasts. Circ. Res. 2014, 114, 67–78. [Google Scholar] [CrossRef]
- Heath, D.; Williams, D.; Harris, P.; Smith, P.; Krüger, H.; Ramirez, A. The pulmonary vasculature of the mountain-viscacha (Lagidium peruanum). The concept of adapted and acclimatized vascular smooth muscle. J. Comp. Pathol. 1981, 91, 293–301. [Google Scholar] [CrossRef]
- Wang, K.; Yang, Y.; Wang, L.; Ma, T.; Shang, H.; Ding, L.; Han, J.; Qiu, Q. Different gene expressions between cattle and yak provide insights into high-altitude adaptation. Anim. Genet. 2016, 47, 28–35. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, D.; Xu, Q.; Li, J.; Shi, W.; Yu, J.; Na, S. Histological Observation of Heart and Respiratory System in Tibet Minipigs. Chin. J. Comp. Med. 2009, 19, 61–62. [Google Scholar]
- Stembridge, M.; Williams, A.M.; Gasho, C.; Dawkins, T.G.; Drane, A.; Villafuerte, F.C.; Levine, B.D.; Shave, R.; Ainslie, P.N. The overlooked significance of plasma volume for successful adaptation to high altitude in Sherpa and Andean natives. Proc. Natl. Acad. Sci. USA 2019, 116, 16177–16179. [Google Scholar] [CrossRef]
- Rao, M.; Li, J.; Qin, J.; Zhang, J.; Gao, X.; Yu, S.; Yu, J.; Chen, G.; Xu, B.; Li, H.; et al. Left Ventricular Function during Acute High-Altitude Exposure in a Large Group of Healthy Young Chinese Men. PLoS ONE 2015, 10, e116936. [Google Scholar] [CrossRef]
- Wu, T. Challenges to humans in the low-oxygen environment of plateau. J. Med. Res. 2006, 10, 1–3. [Google Scholar]
- Beall, C.M. Two routes to functional adaptation: Tibetan and Andean high-altitude natives. Proc. Natl. Acad. Sci. USA 2007, 104 (Suppl. S1), 8655–8660. [Google Scholar] [CrossRef] [PubMed]
- León-Velarde, F.; Gamboa, A.; Chuquiza, J.A.; Esteba, W.A.; Rivera-Chira, M.; Monge, C.C. Hematological parameters in high altitude residents living at 4355, 4660, and 5500 meters above sea level. High Alt. Med. Biol. 2000, 1, 97–104. [Google Scholar] [CrossRef]
- Kong, X.; Dong, X.; Yang, S.; Qian, J.; Yang, J.; Jiang, Q.; Li, X.; Wang, B.; Yan, D.; Lu, S.; et al. Natural selection on TMPRSS6 associated with the blunted erythropoiesis and improved blood viscosity in Tibetan pigs. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2019, 233, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Gou, X.; Wang, Z.; Li, N.; Qiu, F.; Xu, Z.; Yan, D.; Yang, S.; Jia, J.; Kong, X.; Wei, Z.; et al. Whole-genome sequencing of six dog breeds from continuous altitudes reveals adaptation to high-altitude hypoxia. Genome Res. 2014, 24, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Llanos, A.J.; Riquelme, R.A.; Herrera, E.A.; Ebensperger, G.; Krause, B.; Reyes, R.V.; Sanhueza, E.M.; Pulgar, V.M.; Behn, C.; Cabello, G.; et al. Evolving in thin air—Lessons from the llama fetus in the altiplano. Respir. Physiol. Neurobiol. 2007, 158, 298–306. [Google Scholar] [CrossRef]
- Miranda-de La Lama, G.C.; Villarroel, M. Behavioural biology of South American domestic camelids: An overview from a welfare perspective. Small Ruminant Res. 2023, 220, 106918. [Google Scholar] [CrossRef]
- Gonzalez, N.C.; Kuwahira, I. Systemic Oxygen Transport with Rest, Exercise, and Hypoxia: A Comparison of Humans, Rats, and Mice. Compr. Physiol. 2018, 8, 1537–1573. [Google Scholar]
- Wu, T.; Wang, X.; Wei, C.; Cheng, H.; Wang, X.; Li, Y.; Dong, G.; Zhao, H.; Young, P.; Li, G.; et al. Hemoglobin levels in Qinghai-Tibet: Different effects of gender for Tibetans vs. Han. J. Appl. Physiol. 2005, 98, 598–604. [Google Scholar] [CrossRef]
- Chen, Q.-H.; Ge, R.-L.; Wang, X.-Z.; Chen, H.-X.; Wu, T.-Y.; Kobayashi, T.; Yoshimura, K. Exercise performance of Tibetan and Han adolescents at altitudes of 3417 and 4300 m. J. Appl. Physiol. 1997, 83, 661–667. [Google Scholar] [CrossRef]
- Storz, J.F. Hemoglobin-oxygen affinity in high-altitude vertebrates: Is there evidence for an adaptive trend? J. Exp. Biol. 2016, 219 Pt 20, 3190–3203. [Google Scholar] [CrossRef]
- Miao, B.; Wang, Z.; Li, Y. Genomic Analysis Reveals Hypoxia Adaptation in the Tibetan Mastiff by Introgression of the Gray Wolf from the Tibetan Plateau. Mol. Biol. Evol. 2017, 34, 734–743. [Google Scholar] [PubMed]
- Tufts, D.M.; Natarajan, C.; Revsbech, I.G.; Projecto-Garcia, J.; Hoffmann, F.G.; Weber, R.E.; Fago, A.; Moriyama, H.; Storz, J.F. Epistasis constrains mutational pathways of hemoglobin adaptation in high-altitude pikas. Mol. Biol. Evol. 2015, 32, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Storz, J.F.; Runck, A.M.; Moriyama, H.; Weber, R.E.; Fago, A. Genetic differences in hemoglobin function between highland and lowland deer mice. J. Exp. Biol. 2010, 213 Pt 15, 2565–2574. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Guan, Y.; Signore, A.V.; Natarajan, C.; DuBay, S.G.; Cheng, Y.; Han, N.; Song, G.; Qu, Y.; Moriyama, H.; et al. Divergent and parallel routes of biochemical adaptation in high-altitude passerine birds from the Qinghai-Tibet Plateau. Proc. Natl. Acad. Sci. USA 2018, 115, 1865–1870. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, C.; Jendroszek, A.; Kumar, A.; Weber, R.E.; Tame, J.R.H.; Fago, A.; Storz, J.F. Molecular basis of hemoglobin adaptation in the high-flying bar-headed goose. PLoS Genet. 2018, 14, e1007331. [Google Scholar] [CrossRef]
- Storz, J.F. Gene Duplication and Evolutionary Innovations in Hemoglobin-Oxygen Transport. Physiology 2016, 31, 223–232. [Google Scholar] [CrossRef]
- Blunt, M.H.; Huisman, T.H.; Lewis, J.P. The production of haemoglobin C in adult sheep and goats. Aust. J. Exp. Biol. Med. 1969, 47, 601–611. [Google Scholar] [CrossRef]
- Blunt, M.H.; Perry, M.; Lane, R. The production of haemoglobin C by sheep at simulated high altitude. Res. Vet. Sci. 1970, 11, 191–193. [Google Scholar] [CrossRef]
- Samanta, D.; Semenza, G.L. Metabolic adaptation of cancer and immune cells mediated by hypoxia-inducible factors. Biochim. Biophys. Acta-Rev. Cancer 2018, 1870, 15–22. [Google Scholar] [CrossRef]
- Yan, Z.; Yang, J.; Wei, W.-T.; Zhou, M.-L.; Mo, D.-X.; Wan, X.; Ma, R.; Wu, M.-M.; Huang, J.-H.; Liu, Y.-J.; et al. A time-resolved multi-omics atlas of transcriptional regulation in response to high-altitude hypoxia across whole-body tissues. Nat. Commun. 2024, 15, 3970. [Google Scholar] [CrossRef]
- Hochachka, P.; Stanley, C.; Merkt, J.; Sumar-Kalinowski, J. Metabolic meaning of elevated levels of oxidative enzymes in high altitude adapted animals: An interpretive hypothesis. Respir. Physiol. 1983, 52, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Hardy, K.M.; Dillaman, R.M.; Locke, B.R.; Kinsey, S.T. A skeletal muscle model of extreme hypertrophic growth reveals the influence of diffusion on cellular design. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1855–R1867. [Google Scholar] [CrossRef] [PubMed]
- Murray, A.J.; Montgomery, H.E. How wasting is saving: Weight loss at altitude might result from an evolutionary adaptation. Bioessays 2014, 36, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.R.; Elogio, T.S.; Lui, M.A.; Storz, J.F.; Cheviron, Z.A. Adaptive Modifications of Muscle Phenotype in High-Altitude Deer Mice Are Associated with Evolved Changes in Gene Regulation. Mol. Biol. Evol. 2015, 32, 1962–1976. [Google Scholar] [CrossRef]
- Mahalingam, S.; McClelland, G.B.; Scott, G.R. Evolved changes in the intracellular distribution and physiology of muscle mitochondria in high-altitude native deer mice. J. Physiol. 2017, 595, 4785–4801. [Google Scholar] [CrossRef]
- Scott, G.R.; Egginton, S.; Richards, J.G.; Milsom, W.K. Evolution of muscle phenotype for extreme high altitude flight in the bar-headed goose. Proc. R. Soc. B-Biol. Sci. 2009, 276, 3645–3653. [Google Scholar] [CrossRef]
- Hoppeler, H.; Howald, H.; Conley, K.; Lindstedt, S.L.; Claassen, H.; Vock, P.; Weibel, E.R. Endurance training in humans: Aerobic capacity and structure of skeletal muscle. J. Appl. Physiol. 1985, 59, 320–327. [Google Scholar] [CrossRef]
- Jacobs, R.A.; Lundby, A.M.; Fenk, S.; Gehrig, S.; Siebenmann, C.; Flück, D.; Kirk, N.; Hilty, M.P.; Lundby, C. Twenty-eight days of exposure to 3454 m increases mitochondrial volume density in human skeletal muscle. J. Physiol. 2016, 594, 1151–1166. [Google Scholar] [CrossRef]
- Murray, A.J.; Horscroft, J.A. Mitochondrial function at extreme high altitude. J. Physiol. 2016, 594, 1137–1149. [Google Scholar] [CrossRef]
- Levett, D.Z.; Radford, E.J.; Menassa, D.A.; Graber, E.F.; Morash, A.J.; Hoppeler, H.; Clarke, K.; Martin, D.S.; Ferguson-Smith, A.C.; Montgomery, H.E.; et al. Acclimatization of skeletal muscle mitochondria to high-altitude hypoxia during an ascent of Everest. FASEB J. 2012, 26, 1431–1441. [Google Scholar] [CrossRef]
- Welch, K.C.; Altshuler, D.L.; Suarez, R.K. Oxygen consumption rates in hovering hummingbirds reflect substrate-dependent differences in P/O ratios: Carbohydrate as a ‘premium fuel’. J. Exp. Biol. 2007, 210, 2146. [Google Scholar] [CrossRef] [PubMed]
- Ge, R.-L.; Simonson, T.S.; Cooksey, R.C.; Tanna, U.; Qin, G.; Huff, C.D.; Witherspoon, D.J.; Xing, J.; Zhengzhong, B.; Prchal, J.T.; et al. Metabolic insight into mechanisms of high-altitude adaptation in Tibetans. Mol. Genet. Metab. 2012, 106, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Horscroft, J.A.; Kotwica, A.O.; Laner, V.; West, J.A.; Hennis, P.J.; Levett, D.Z.H.; Howard, D.J.; Fernandez, B.O.; Burgess, S.L.; Ament, Z.; et al. Metabolic basis to Sherpa altitude adaptation. Proc. Natl. Acad. Sci. USA 2017, 114, 6382–6387. [Google Scholar] [CrossRef] [PubMed]
- Lau, D.S.; Connaty, A.D.; Mahalingam, S.; Wall, N.; Cheviron, Z.A.; Storz, J.F.; Scott, G.R.; McClelland, G.B. Acclimation to hypoxia increases carbohydrate use during exercise in high-altitude deer mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R400–R411. [Google Scholar] [CrossRef]
- Schippers, M.-P.; Ramirez, O.; Arana, M.; Pinedo-Bernal, P.; McClelland, G.B. Increase in carbohydrate utilization in high-altitude Andean mice. Curr. Biol. 2012, 22, 2350–2354. [Google Scholar] [CrossRef]
- Murray, A.J.; Montgomery, H.E.; Feelisch, M.; Grocott, M.P.; Martin, D.S. Metabolic adjustment to high-altitude hypoxia: From genetic signals to physiological implications. Biochem. Soc. Trans. 2018, 46, 599–607. [Google Scholar] [CrossRef]
- Holden, J.E.; Stone, C.K.; Clark, C.M.; Brown, W.D.; Nickles, R.J.; Stanley, C.; Hochachka, P.W. Enhanced cardiac metabolism of plasma glucose in high-altitude natives: Adaptation against chronic hypoxia. J. Appl. Physiol. 1995, 79, 222–228. [Google Scholar] [CrossRef]
- Dosek, A.; Ohno, H.; Acs, Z.; Taylor, A.W.; Radak, Z. High altitude and oxidative stress. Respir. Physiol. Neurobiol. 2007, 158, 128–131. [Google Scholar] [CrossRef]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef]
- Chandel, N.S.; McClintock, D.S.; Feliciano, C.E.; Wood, T.M.; Melendez, J.A.; Rodriguez, A.M.; Schumacker, P.T. Reactive oxygen species generated at mitochondrial complex III stabilize hypoxia-inducible factor-1alpha during hypoxia: A mechanism of O2 sensing. J. Biol. Chem. 2000, 275, 25130–25138. [Google Scholar] [CrossRef]
- Gelfi, C.; De Palma, S.; Ripamonti, M.; Wait, R.; Eberini, I.; Bajracharya, A.; Marconi, C.; Schneider, A.; Hoppeler, H.; Cerretelli, P. New aspects of altitude adaptation in Tibetans: A proteomic approach. FASEB J. 2004, 18, 612–614. [Google Scholar] [CrossRef] [PubMed]
- Dawson, N.J.; Lyons, S.A.; Henry, D.A.; Scott, G.R. Effects of chronic hypoxia on diaphragm function in deer mice native to high altitude. Acta Physiol. 2018, 223, e13030. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Bátkai, S.; Kunos, G. The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol. Rev. 2006, 58, 389–462. [Google Scholar] [CrossRef] [PubMed]
- Alarcón-Yaquetto, D.E.; Caballero, L.; Gonzales, G.F. Association Between Plasma N-Acylethanolamides and High Hemoglobin Concentration in Southern Peruvian Highlanders. High Alt. Med. Biol. 2017, 18, 322–329. [Google Scholar] [CrossRef]
- van Doorslaer de Ten Ryen, S.; Dalle, S.; Terrasi, R.; Koppo, K.; Muccioli, G.G.; Deldicque, L. Regulation of the endocannabinoid system by endurance and resistance exercise in hypoxia in human skeletal muscle. J. Appl. Physiol. 2023, 134, 569–580. [Google Scholar] [CrossRef]
- Alonso-Alconada, D.; Álvarez, F.J.; Goñi-De-Cerio, F.; Hilario, E.; Álvarez, A. Cannabinoid-mediated Modulation of Oxidative Stress and Early Inflammatory Response after Hypoxia-Ischemia. Int. J. Mol. Sci. 2020, 21, 1283. [Google Scholar] [CrossRef]
- Benyó, Z.; Ruisanchez, É.; Leszl-Ishiguro, M.; Sándor, P.; Pacher, P. Endocannabinoids in cerebrovascular regulation. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H785–H801. [Google Scholar] [CrossRef]
- Tree, K.C.; Scotto di Perretolo, M.; Peyronnet, J.; Cayetanot, F. In utero cannabinoid exposure alters breathing and the response to hypoxia in newborn mice. Eur. J. Neurosci. 2014, 40, 2196–2204. [Google Scholar] [CrossRef]
- Lindborg, K.A.; Teachey, M.K.; Jacob, S.; Henriksen, E.J. Effects of in vitro antagonism of endocannabinoid-1 receptors on the glucose transport system in normal and insulin-resistant rat skeletal muscle. Diabetes Obes. Metab. 2010, 12, 722–730. [Google Scholar] [CrossRef]
- Simonson, T.S.; Yang, Y.; Huff, C.D.; Yun, H.; Qin, G.; Witherspoon, D.J.; Bai, Z.; Lorenzo, F.R.; Xing, J.; Jorde, L.B.; et al. Genetic evidence for high-altitude adaptation in Tibet. Science 2010, 329, 72–75. [Google Scholar] [CrossRef]
- Yi, X.; Liang, Y.; Huerta-Sanchez, E.; Jin, X.; Cuo, Z.X.P.; Pool, J.E.; Xu, X.; Jiang, H.; Vinckenbosch, N.; Korneliussen, T.S.; et al. Sequencing of 50 human exomes reveals adaptation to high altitude. Science 2010, 329, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Bigham, A.; Bauchet, M.; Pinto, D.; Mao, X.; Akey, J.M.; Mei, R.; Scherer, S.W.; Julian, C.G.; Wilson, M.J.; Herráez, D.L.; et al. Identifying signatures of natural selection in Tibetan and Andean populations using dense genome scan data. PLoS Genet. 2010, 6, e1001116. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, F.R.; Huff, C.; Myllymäki, M.; Olenchock, B.; Swierczek, S.; Tashi, T.; Gordeuk, V.; Wuren, T.; Ri-Li, G.; McClain, D.A.; et al. A genetic mechanism for Tibetan high-altitude adaptation. Nat. Genet. 2014, 46, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Jin, Z.-B.; Chen, J.; Huang, X.-F.; Li, X.-M.; Liang, Y.-B.; Mao, J.-Y.; Chen, X.; Zheng, Z.; Bakshi, A.; et al. Genetic signatures of high-altitude adaptation in Tibetans. Proc. Natl. Acad. Sci. USA 2017, 114, 4189–4194. [Google Scholar] [CrossRef]
- Bigham, A.W.; Mao, X.; Mei, R.; Brutsaert, T.; Wilson, M.J.; Julian, C.G.; Parra, E.J.; Akey, J.M.; Moore, L.G.; Shriver, M.D. Identifying positive selection candidate loci for high-altitude adaptation in Andean populations. Hum. Genom. 2009, 4, 79–90. [Google Scholar] [CrossRef]
- Bigham, A.W.; Julian, C.G.; Wilson, M.J.; Vargas, E.; Browne, V.A.; Shriver, M.D.; Moore, L.G. Maternal PRKAA1 and EDNRA genotypes are associated with birth weight, and PRKAA1 with uterine artery diameter and metabolic homeostasis at high altitude. Physiol. Genom. 2014, 46, 687–697. [Google Scholar] [CrossRef]
- Huerta-Sánchez, E.; DeGiorgio, M.; Pagani, L.; Tarekegn, A.; Ekong, R.; Antao, T.; Cardona, A.; Montgomery, H.E.; Cavalleri, G.L.; Robbins, P.A.; et al. Genetic signatures reveal high-altitude adaptation in a set of Ethiopian populations. Mol. Biol. Evol. 2013, 30, 1877–1888. [Google Scholar] [CrossRef]
- Udpa, N.; Ronen, R.; Zhou, D.; Liang, J.; Stobdan, T.; Appenzeller, O.; Yin, Y.; Du, Y.; Guo, L.; Cao, R.; et al. Whole genome sequencing of Ethiopian highlanders reveals conserved hypoxia tolerance genes. Genome Biol. 2014, 15, R36. [Google Scholar] [CrossRef]
- Stobdan, T.; Zhou, D.; Ao-Ieong, E.; Ortiz, D.; Ronen, R.; Hartley, I.; Gan, Z.; McCulloch, A.D.; Bafna, V.; Cabrales, P.; et al. Endothelin receptor B, a candidate gene from human studies at high altitude, improves cardiac tolerance to hypoxia in genetically engineered heterozygote mice. Proc. Natl. Acad. Sci. USA 2015, 112, 10425–10430. [Google Scholar] [CrossRef]
- Wu, H.; Liu, Y.-H.; Wang, G.-D.; Yang, C.-T.; Otecko, N.O.; Liu, F.; Wu, S.-F.; Wang, L.; Yu, L.; Zhang, Y.-P. Identifying molecular signatures of hypoxia adaptation from sex chromosomes: A case for Tibetan Mastiff based on analyses of X chromosome. Sci. Rep. 2016, 6, 35004. [Google Scholar] [CrossRef]
- Song, S.; Yao, N.; Yang, M.; Liu, X.; Dong, K.; Zhao, Q.; Pu, Y.; He, X.; Guan, W.; Yang, N.; et al. Exome sequencing reveals genetic differentiation due to high-altitude adaptation in the Tibetan cashmere goat (Capra hircus). BMC Genomics 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, R.M.; Velotta, J.P.; Ivy, C.M.; Jones, M.R.; Muir, S.M.; Bradburd, G.S.; Storz, J.F.; Scott, G.R.; Cheviron, Z.A. Physiological and genomic evidence that selection on the transcription factor Epas1 has altered cardiovascular function in high-altitude deer mice. PLoS Genet. 2019, 15, e1008420. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Luosang, J.; Hua, S.; He, J.; Ciren, A.; Wang, W.; Tong, X.; Liang, Y.; Wang, J.; Zheng, X. High altitude adaptation and phylogenetic analysis of Tibetan horse based on the mitochondrial genome. J. Genet. Genom. 2007, 34, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.-D.; Ding, X.-D.; Wang, S.; Wójcik, J.M.; Zhang, Y.; Tokarska, M.; Li, Y.; Wang, M.-S.; Faruque, O.; Nielsen, R.; et al. Pervasive introgression facilitated domestication and adaptation in the Bos species complex. Nat. Ecol. Evol. 2018, 2, 1139–1145. [Google Scholar] [CrossRef]
- Qiu, Q.; Zhang, G.; Ma, T.; Qian, W.; Wang, J.; Ye, Z.; Cao, C.; Hu, Q.; Kim, J.; Larkin, D.M.; et al. The yak genome and adaptation to life at high altitude. Nat. Genet. 2012, 44, 946–949. [Google Scholar] [CrossRef]
- Wang, M.-S.; Li, Y.; Peng, M.-S.; Zhong, L.; Wang, Z.-J.; Li, Q.-Y.; Tu, X.-L.; Dong, Y.; Zhu, C.-L.; Wang, L.; et al. Genomic Analyses Reveal Potential Independent Adaptation to High Altitude in Tibetan Chickens. Mol. Biol. Evol. 2015, 32, 1880–1889. [Google Scholar] [CrossRef]
- Ai, H.; Yang, B.; Li, J.; Xie, X.; Chen, H.; Ren, J. Population history and genomic signatures for high-altitude adaptation in Tibetan pigs. BMC Genom. 2014, 15, 834. [Google Scholar] [CrossRef]
- Yang, J.; Li, W.-R.; Lv, F.-H.; He, S.-G.; Tian, S.-L.; Peng, W.-F.; Sun, Y.-W.; Zhao, Y.-X.; Tu, X.-L.; Zhang, M.; et al. Whole-Genome Sequencing of Native Sheep Provides Insights into Rapid Adaptations to Extreme Environments. Mol. Biol. Evol. 2016, 33, 2576–2592. [Google Scholar] [CrossRef]
- Gorkhali, N.A.; Dong, K.; Yang, M.; Song, S.; Kader, A.; Shrestha, B.S.; He, X.; Zhao, Q.; Pu, Y.; Li, X.; et al. Genomic analysis identified a potential novel molecular mechanism for high-altitude adaptation in sheep at the Himalayas. Sci. Rep. 2016, 6, 29963. [Google Scholar] [CrossRef]
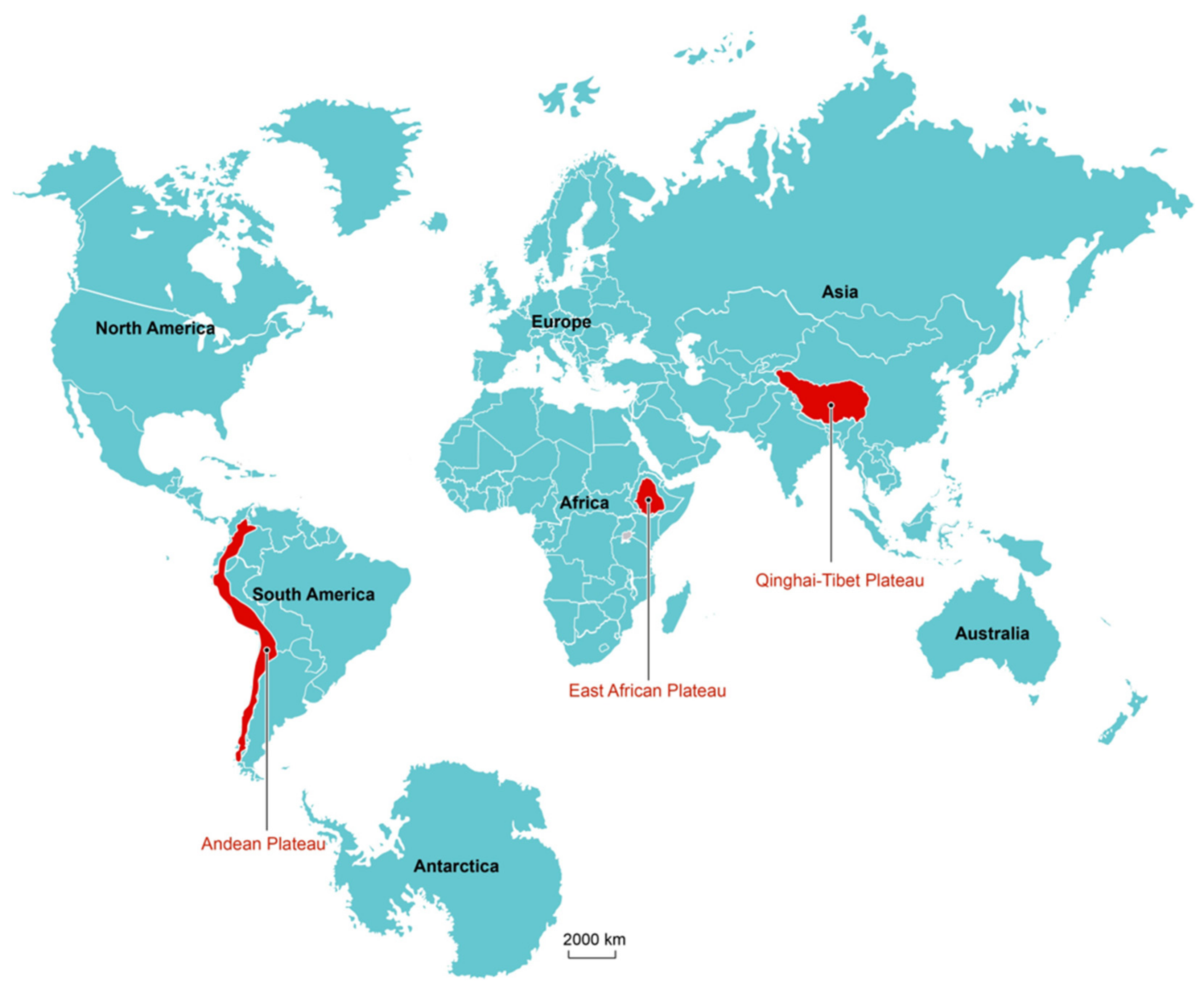
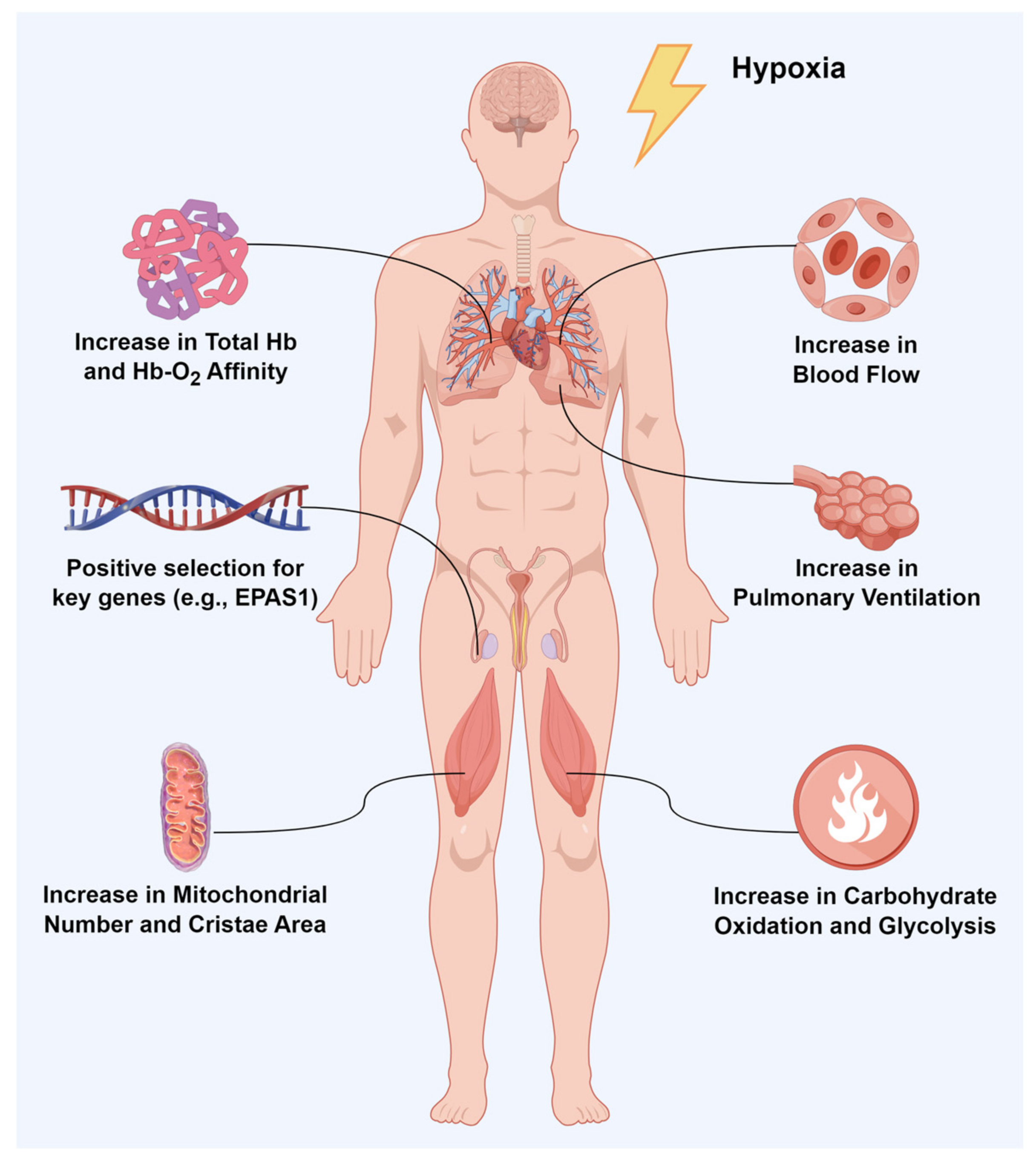

{kind=link}
{kind=link}
| Research Object | Positively Selected Genes | Main Functions of These Genes in High-Altitude Hypoxia Adaptation |
|---|---|---|
| Tibetans [94,95,96,97,98] | EGLN1, EPAS1, PPARα, HMOX2 | Degraded HIF-α (EGLN1); Associated with blood traits (EPAS1, PPARα, HMOX2) |
| Andean natives [96,99,100] | EGLN1, EDNRA, PRKAA1, NOS2A | Associated with greater birth, weight, and uterine artery diameters (EDNRA, PRKAA1) |
| Amharas and Oromos [101,102,103] | BHLHE41, CIC, LIPE, PAFAH1B3, EDNRB | Involved in the initiation of hypoxia response through the HIF pathway (BHLHE41); Associated with enhanced hypoxic tolerance (CIC, LIPE, PAFAH1B3, EDNRB) |
| Tibetan mastiffs [48,55,104] | EPAS1, HBB, AMOT | Reduced blood flow resistance (EPAS1); Increased Hb–O2 affinity (HBB); Blood pressure regulation (AMOT) |
| Tibetan cashmere goats [105] | EPAS1 | Hypoxia adaptation |
| Deer mice [106] | EPAS1 | Associated with cardiovascular function and transcriptional responses to hypoxia |
| Tibetan horse [13,107] | EPAS1, NADH6 | Promoted blood circulation and O2 transport (EPAS1); Associated with energy metabolism (NADH6) |
| Yak [108,109] | EGLN1, ADAM17, ARG2 | Reduced Hb concentration and hematocrit (EGLN1); Involved in hypoxic stress response (ADAM17, ARG2) |
| Tibetan chickens [110] | RYR2 | Associated with hypoxia tolerance |
| Tibetan pig [111] | RGCC, GRIN2B | Involved in hypoxia-induced anti-angiogenesis (RGCC); Neural response (GRIN2B) |
| Tibetan sheep [112,113] | SOCS2, FGF7 | Associated with energy metabolism (SOCS2); Inhibition of hypoxia-induced lung injury (FGF7) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, P.; Li, S.; He, Z.; Ma, X. Physiological and Genetic Basis of High-Altitude Indigenous Animals’ Adaptation to Hypoxic Environments. Animals 2024, 14, 3031. https://doi.org/10.3390/ani14203031
Zhao P, Li S, He Z, Ma X. Physiological and Genetic Basis of High-Altitude Indigenous Animals’ Adaptation to Hypoxic Environments. Animals. 2024; 14(20):3031. https://doi.org/10.3390/ani14203031
Chicago/Turabian StyleZhao, Pengfei, Shaobin Li, Zhaohua He, and Xiong Ma. 2024. "Physiological and Genetic Basis of High-Altitude Indigenous Animals’ Adaptation to Hypoxic Environments" Animals 14, no. 20: 3031. https://doi.org/10.3390/ani14203031
APA StyleZhao, P., Li, S., He, Z., & Ma, X. (2024). Physiological and Genetic Basis of High-Altitude Indigenous Animals’ Adaptation to Hypoxic Environments. Animals, 14(20), 3031. https://doi.org/10.3390/ani14203031