
Research on the Changes in Distribution and Habitat Suitability of the Chinese Red Panda Population

Abstract
:Simple Summary
Abstract
1. Instruction
2. Material and Methods
2.1. Study Area and Species Distribution Data
2.2. Environmental Factors
2.3. Dynamic Assessment of Habitat Suitability at Multiple Spatiotemporal Scales
2.4. Analysis of Habitat Suitability Decline in the Liangshan Mountains
3. Results
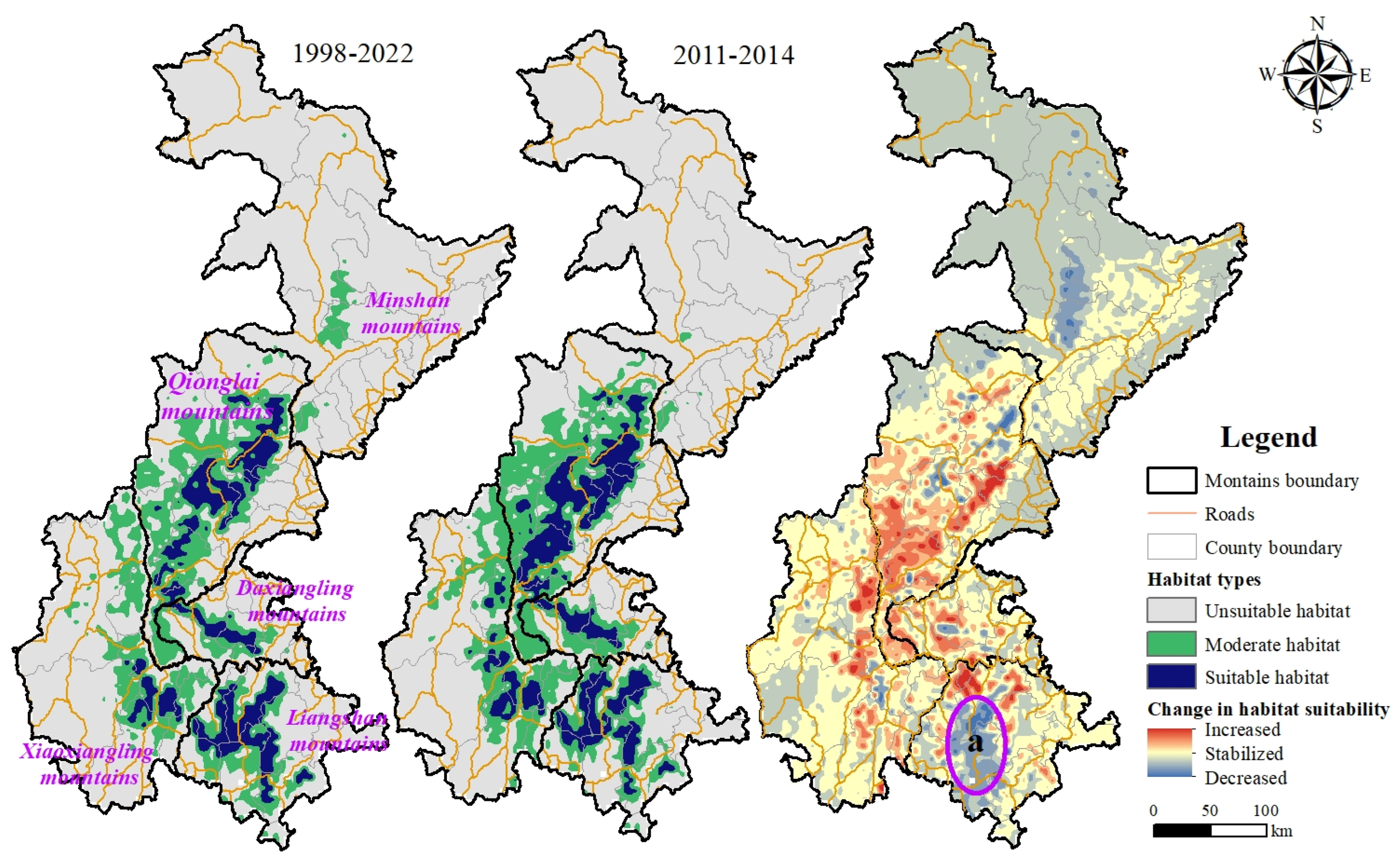
3.1. Habitat Distribution and Habitat Suitability Dynamic Change at Different Time Scales
3.2. Analysis of Habitat Suitability Change in the Liangshan Mountains
4. Discussion
4.1. Habitat Change at Different Time Scales
4.2. The Causes of Habitat Suitability Decline
5. Conclusions
6. Protection Management Proposal
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, Y.; Thapa, A.; Fan, H.; Ma, T.; Wu, Q.; Ma, S.; Zhang, D.; Wang, B.; Li, M.; Yan, L. Genomic evidence for two phylogenetic species and long-term population bottlenecks in red pandas. Sci. Adv. 2020, 6, eaax5751. [Google Scholar] [CrossRef]
- Choudhury, A. An overview of the status and conservation of the red panda Ailurus fulgens in India, with reference to its global status. Oryx 2001, 35, 250–259. [Google Scholar] [CrossRef]
- Chinese State Forestry Administration. Results of the Fourth National Survey on the Giant Panda; Science Press: Beijing, China, 2015. [Google Scholar]
- Thapa, A.; Aryal Prakash, C.; Singh Paras, B.; Shah Karan, B. The endangered red panda in Himalayas: Potential distribution and ecological habitat associates. Glob. Ecol. Conserv. 2020, 21, e00890. [Google Scholar] [CrossRef]
- Wei, F.; Feng, Z.; Wang, Z.; Hu, J. Current distribution, status and conservation of wild red pandas Ailurus fulgens in China. Biol. Conserv. 1999, 89, 285–291. [Google Scholar] [CrossRef]
- Wei, F.; Wang, Z.; Feng, Z.; Ming, L.; Ang, Z. Seasonal energy utilization in bamboo by the red panda (Ailurus fulgens). Zoo Biol. 2000, 19, 27–33. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, J.; Han, Z.; Wei, F. Activity patterns of wild red pandas in Fengtongzhai Nature Reserve, China. Ital. J. Zool. 2011, 78, 398–404. [Google Scholar] [CrossRef]
- Wei, W.; Fuwen, W.; Jingchu, H.; Zuojian, F.; Guang, Y. Habitat selection by red panda in Mabian Dafending Reserve. Acta Theriol. Sin. 1998, 18, 15–20. [Google Scholar]
- Wei, F.; Feng, Z.; Wang, Z.; Hu, J. Habitat use and separation between the giant panda and the red panda. J. Mammal. 2000, 81, 448–455. [Google Scholar] [CrossRef]
- Wei, F.; Feng, Z.; Wang, Z.; Li, M. Feeding strategy and resource partitioning between giant and red pandas. Mammalia 1999, 63, 417–430. [Google Scholar] [CrossRef]
- Ledig, F.T. Human impacts on genetic diversity in forest ecosystems. Oikos 1992, 63, 87–108. [Google Scholar] [CrossRef]
- Sánchez-Cordero, V.; Illoldi-Rangel, P.; Escalante, T.; Figueroa, F.; Rodríguez, G.; Linaje, M.; Sarkar, S. Deforestation and biodiversity conservation in Mexico. Endanger. Spec. New Res. 2009, 1, 279–297. [Google Scholar]
- Fox, J.; Yonzon, P.; Podger, N. Mapping conflicts between biodiversity and human needs in Langtang National Park, Nepal. Conserv. Biol. 1996, 10, 562–569. [Google Scholar] [CrossRef]
- Yonzon, P.B. Ecology and Conservation of the Red Panda in the Nepal Himalayas; The University of Maine: Orono, ME, USA, 1989. [Google Scholar]
- Glatston, A.; Wei, F.; Than, Z.; Sherpa, A. Ailurus fulgens. The IUCN Red List of Threatened Species. 2015. Available online: https://www.iucnredlist.org/species/714/110023718 (accessed on 16 December 2023).
- Yang, H.; Huang, Q.; Zhang, J.; Songer, M.; Liu, J. Range-wide assessment of the impact of china’s nature reserves on giant panda habitat quality. Sci. Total Environ. 2021, 769, 145081. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, S.; Saha, G.K.; Khan, J.A. Ecology of the red panda Ailurus fulgens in the Singhalila National Park, Darjeeling, India. Biol. Conserv. 2001, 98, 11–18. [Google Scholar] [CrossRef]
- Wei, F.; Feng, Z.; Wang, Z.; Zhou, A.; Hu, J. Use of the nutrients in bamboo by the red panda (Ailurus fulgens). J. Zool. 1999, 248, 535–541. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, J.; Yang, J.; Li, M.; Wei, F. Food habits and space-use of red pandas Ailurus fulgens in the Fengtongzhai Nature Reserve, China: Food effects and behavioural responses. ActaTheriol. 2009, 54, 225–234. [Google Scholar] [CrossRef]
- Thapa, A.; Hu, Y.; Wei, F. The endangered red panda (Ailurus fulgens): Ecology and conservation approaches across the entire range. Biol. Conserv. 2018, 220, 112–121. [Google Scholar] [CrossRef]
- Zhu, G.; Liu, G.; Bu, W.; Gao, Y. Ecological niche modeling and its applications in biodiversity conservation. Biodivers. Sci. 2013, 21, 90–98. [Google Scholar]
- Bhatta, M.; Zander, K.K.; Austin, B.J.; Garnett, S.T. Societal recognition of ecosystem service flows from red panda habitats in Western Nepal. Mt. Res. Dev. 2020, 40, R50. [Google Scholar] [CrossRef]
- Lowther, D.A. The first painting of the red panda (Ailurus fulgens) in Europe? Natural history and artistic patronage in early nineteenth-century India. Arch. Nat. Hist. 2021, 48, 368–376. [Google Scholar] [CrossRef]
- Su, H.; Bista, M.; Li, M. Mapping habitat suitability for Asiatic black bear and red panda in Makalu Barun National Park of Nepal from Maxent and GARP models. Sci. Rep. 2021, 11, 14135. [Google Scholar] [CrossRef] [PubMed]
- Wanghe, K.; Guo, X.; Hu, F.; Ahmad, S.; Jin, X.; Khan, T.U.; Xiao, Y.; Luan, X. Spatial coincidence between mining activities and protected areas of giant panda habitat: The geographic overlaps and implications for conservation. Biol. Conserv. 2020, 247, 108600. [Google Scholar] [CrossRef]
- Chinese State Forestry Administration. The Third National Survey Report of the Giant Panda in China; Science Press: Beijing, China, 2006. [Google Scholar]
- Morelle, K.; Lejeune, P. Seasonal variations of wild boar Sus scrofa distribution in agricultural landscapes: A species distribution modelling approach. Eur. J. Wildl. Res. 2015, 61, 45–56. [Google Scholar] [CrossRef]
- Telemeco, R.S.; Warner, D.A.; Reida, M.K.; Janzen, F.J. Extreme developmental temperatures result in morphological abnormalities in painted turtles (Chrysemys picta): A climate change perspective. Integr. Zool. 2013, 8, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Blair, R.B.; Launer, A.E. Butterfly diversity and human land use: Species assemblages along an urban grandient. Biol. Conserv. 1997, 80, 113–125. [Google Scholar] [CrossRef]
- Sirami, C.; Caplat, P.; Popy, S.; Clamens, A.; Arlettaz, R.; Jiguet, F.; Brotons, L.; Martin, J.L. Impacts of global change on species distributions: Obstacles and solutions to integrate climate and land use. Glob. Ecol. Biogeogr. 2017, 26, 385–394. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA. 2017, 114, E6089–E6096. [Google Scholar] [CrossRef]
- Laliberte, A.S.; Ripple, W.J. Range contractions of North American carnivores and ungulates. BioScience 2004, 54, 123–138. [Google Scholar] [CrossRef]
- Huang, P.; Bian, K.; Huang, Z.; Li, Q.; Dunn, D.W.; Fang, G.; Liu, J.; Wang, M.; Yang, X.; Pan, R. Human activities and elevational constraints restrict ranging patterns of snub-nosed monkeys in a mountainous refuge. Integr. Zool. 2021, 16, 202–213. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Sunday, J.M.; Bates, A.E.; Dulvy, N.K. Thermal tolerance and the global redistribution of animals. Nat. Commun. 2012, 2, 686–690. [Google Scholar] [CrossRef]
- Abrahms, B.; Carter, N.H.; Clark-Wolf, T.; Gaynor, K.M.; Johansson, E.; McInturff, A.; Nisi, A.C.; Rafiq, K.; West, L. Climate change as a global amplifier of human–wildlife conflict. Nat. Commun. 2023, 13, 224–234. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.; De Siqueira, M.F.; Grainger, A.; Hannah, L. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Mainka, S.A.; Howard, G.W. Climate change and invasive species: Double jeopardy. Integr. Zool. 2010, 5, 102–111. [Google Scholar] [CrossRef]
- Li, X.; Hu, W.; Bleisch, W.V.; Li, Q.; Wang, H.; Lu, W.; Sun, J.; Zhang, F.; Ti, B.; Jiang, X. Functional diversity loss and change in nocturnal behavior of mammals under anthropogenic disturbance. Conserv. Biol. 2022, 36, e13839. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, M.; Li, J. The impact of ecological protection projects on giant panda habitats. Acta. Ecol. Sin. 2011, 31, 154–163. [Google Scholar]
- Wang, F.; Winkler, J.; Vina, A.; McShea, W.J.; Li, S.; Connor, T.; Zhao, Z.; Wang, D.; Yang, H.; Tang, Y. The hidden risk of using umbrella species as conservation surrogates: A spatio-temporal approach. Biol. Conserv. 2021, 253, 108913. [Google Scholar] [CrossRef]
- Gao, H.; Guan, T.; Zhu, D.; Li, W.; Zhou, F.; Zhao, D.; Li, C.; Zhang, L. Assessment of effective conservation of the Sichuan takin by giant panda reserves through functional zoning. Integr. Zool. 2020, 15, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Palik, B.J.; Goebel, P.C.; Kirkman, L.K.; West, L. Using landscape hierarchies to guide restoration of disturbed ecosystems. Ecol. Appl. 2000, 10, 189–202. [Google Scholar] [CrossRef]
- Bista, D.; Shrestha, S.; Sherpa, P.; Thapa, G.J.; Kokh, M.; Lama, S.T.; Khanal, K.; Thapa, A.; Jnawali, S.R. Distribution and habitat use of red panda in the Chitwan-Annapurna Landscape of Nepal. PLoS ONE 2017, 12, e0178797. [Google Scholar] [CrossRef]
- Yonzon, P.B.; Hunter, M.L., Jr. Conservation of the red panda Ailurus fulgens. Biol. Conserv. 1991, 57, 1–11. [Google Scholar] [CrossRef]
- Li, Y.; Rao, T.; Gai, L.; Price, M.L.; Yuxin, L.; Jianghong, R. Giant pandas are losing their edge: Population trend and distribution dynamic drivers of the giant panda. Glob. Change Biol. 2023, 29, 4480–4495. [Google Scholar] [CrossRef]
- Nielsen, E.S.; Henriques, R.; Beger, M.; von der Heyden, S. Distinct interspecific and intraspecific vulnerability of coastal species to global change. Glob. Change Biol. 2021, 27, 3415–3431. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Sheppard, J.K.; Swaisgood, R.R.; Wang, G.; Nie, Y.; Wei, W.; Zhao, N.; Wei, F. Ecological scale and seasonal heterogeneity in the spatial behaviors of giant pandas. Integr. Zool. 2014, 9, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Pigot, A.L.; Merow, C.; Wilson, A.; Trisos, C.H. Abrupt expansion of climate change risks for species globally. Nat. Ecol. Evol. 2023, 7, 1060–1071. [Google Scholar] [CrossRef]
- Schultz, E.L.; Hülsmann, L.; Pillet, M.D.; Hartig, F.; Breshears, D.D.; Record, S.; Shaw, J.D.; DeRose, R.J.; Zuidema, P.A.; Evans, M.E. Climate-driven, but dynamic and complex? A reconciliation of competing hypotheses for species’ distributions. Ecol. Lett. 2022, 25, 38–51. [Google Scholar] [CrossRef]
- Lenoir, J.; Bertrand, R.; Comte, L.; Bourgeaud, L.; Hattab, T.; Murienne, J.; Grenouillet, G. Species better track climate warming in the oceans than on land. Nat. Ecol. Evol. 2020, 4, 1044–1059. [Google Scholar] [CrossRef]
- De Frenne, P.; Lenoir, J.; Luoto, M.; Scheffers, B.R.; Zellweger, F.; Aalto, J.; Ashcroft, M.B.; Christiansen, D.M.; Decocq, G.; De Pauw, K. Forest microclimates and climate change: Importance, drivers and future research agenda. Glob. Change Biol. 2021, 27, 2279–2297. [Google Scholar] [CrossRef]
- Yang, L.; Shi, K.; Ma, C.; Ren, G.; Fan, P. Mechanisms underlying altitudinal and horizontal range contraction: The western black crested gibbon. J. Biogeogr. 2021, 48, 321–331. [Google Scholar] [CrossRef]
- Dong, F.; Zhang, Q.; Chen, Y.L.; Lei, F.M.; Li, S.H.; Wu, F.; Yang, X.J. Potential millennial-scale avian declines by humans in southern China. Glob. Change Biol. 2022, 28, 5505–5513. [Google Scholar] [CrossRef] [PubMed]
- Fourcade, Y.; WallisDeVries, M.F.; Kuussaari, M.; van Swaay, C.A.; Heliölä, J.; Öckinger, E. Habitat amount and distribution modify community dynamics under climate change. Ecol. Lett. 2021, 24, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Xu, W.; Xiao, Y.; Pimm, S.L.; Shi, H.; Ouyang, Z. Spatial models of giant pandas under current and future conditions reveal extinction risks. Nat. Ecol. Evol. 2021, 5, 1309–1316. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yang, B.; Dai, Q.; Pan, H.; Zhong, X.; Ran, J.; Yang, X.; Gu, X.; Yang, Z.; Qi, D. Landscape-scale giant panda conservation based on metapopulations within China’s national park system. Sci. Adv. 2022, 8, eabl8637. [Google Scholar] [CrossRef] [PubMed]
- Lei, W.; Wei, W.; Pu, D.; Qubi, S.; Zhou, H.; Hong, M.; Tang, J.; Han, H. Comparative analysis of trophic niche using stable isotopes provides insight into resource use of giant pandas. Integr. Zool. 2023, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Wu, Q.; Wang, L.; Zhang, L.; Wei, F. Lineage-specific evolution of bitter taste receptor genes in the giant and red pandas implies dietary adaptation. Integr. Zool. 2018, 13, 152–159. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, D.I.; Nichols, J.D.; Hines, J.E.; Knutson, M.G.; Franklin, A.B. Estimating site occupancy, colonization, and local extinction when a species is detected imperfectly. Ecol. Evol. 2003, 84, 2200–2207. [Google Scholar] [CrossRef]
- Von Seth, J.; Dussex, N.; Díez-del-Molino, D.; Van Der Valk, T.; Kutschera, V.E.; Kierczak, M.; Steiner, C.C.; Liu, S.; Gilbert, M.T.P.; Sinding, M.-H.S. Genomic insights into the conservation status of the world’s last remaining Sumatran rhinoceros populations. Nat. Commun. 2021, 12, 2393. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Nadeau, C.P.; Urban, M.C.; Bridle, J.R. Climates past, present, and yet-to-come shape climate change vulnerabilities. Trends Ecol. Evol. 2017, 32, 786–800. [Google Scholar] [CrossRef]
- Li, Y.; Cohen, J.M.; Rohr, J.R. Review and synthesis of the effects of climate change on amphibians. Integr. Zool. 2013, 8, 145–161. [Google Scholar] [CrossRef]
- Meehl, G.A.; Tebaldi, C. More intense, more frequent, and longer lasting heat waves in the 21st century. Science 2004, 305, 994–997. [Google Scholar] [CrossRef]
- Zhang, J.; Pan, S.; Che, Q.; Wei, W.; Zhao, X.; Tang, J. Impacts of climate change on the distributions and diversity of the giant panda with its sympatric mammalian species. Ecol. Indic. 2022, 144, 109452. [Google Scholar] [CrossRef]
- Nicholson, J.M.; Van Manen, F.T. Using occupancy models to determine mammalian responses to landscape changes. Integr. Zool. 2009, 4, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Thapa, A.; Wu, R.; Hu, Y.; Nie, Y.; Singh, P.B.; Khatiwada, J.R.; Yan, L.; Gu, X.; Wei, F. Predicting the potential distribution of the endangered red panda across its entire range using MaxEnt modeling. Ecol. Evol. 2018, 8, 10542–10554. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Dong, L.; Zhang, S.; Ran, J.; Yue, B. Landscape pattern of giant panda habitat in the Liangshan Mountains, Sichuan, China. Chines. J. Appl. Environ. Biol. 2010, 16, 179–184. [Google Scholar] [CrossRef]
- Li, C.; Connor, T.; Bai, W.; Yang, H.; Zhang, J.; Qi, D.; Zhou, C. Dynamics of the giant panda habitat suitability in response to changing anthropogenic disturbance in the Liangshan Mountains. Biol. Conserv. 2019, 237, 445–455. [Google Scholar] [CrossRef]
- Rong, Z.; Zhou, H.; Wei, W.; Zhang, J.; Sheng, L.; Zhang, Z. Giant panda habitat suitability assessment in Tangjiahe Nature Reserve based on MAXENT model. J. Lanzhou. Univ. 2017, 53, 269–273. [Google Scholar]
- Sun, Y.; Wang, S.; Feng, J.; Ge, J.; Wang, T. Free-ranging livestock changes the acoustic properties of summer soundscapes in a Northeast Asian temperate forest. Biol. Conserv. 2023, 283, 110–123. [Google Scholar] [CrossRef]
- Du Preez, P.J.; Brown, L.R. Impact of domestic animals on ecosystem integrity of Lesotho high altitude peatlands. Ecosyst. Bodivers. 2011, 12, 249–270. [Google Scholar]
- McChesney, G.J.; Tershy, B.R. History and status of introduced mammals and impacts to breeding seabirds on the California Channel and northwestern Baja California Islands. Col. Waterbirds 1998, 21, 335–347. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor Type | Environmental Factors | Description | Unit |
|---|---|---|---|
| Climate | BIO1 | Annual Mean Temperature | °C |
| BIO4 | Temperature Seasonality | - | |
| BIO14 | Precipitation of Driest Month | mm | |
| Terrain | ELEVATION | Mean Elevation | m |
| SLOPE | Mean Slope | ° | |
| Land Use | FARMLAND | Proportion of Cropland Area | % |
| FOREST | Proportion of Forest Area | % | |
| ROAD | Distance to Main Road | m |
| Mountains | 3rd Survey Area (km2) | 4th Survey Area (km2) | ΔArea (km2) | Relative Change (%) |
|---|---|---|---|---|
| Minshan | 0.00 | 0.00 | 0.00 | 0.00 |
| Qionglai | 4094.97 | 5717.70 | 1622.73 | 39.63 |
| Daxiangling | 828.92 | 1059.22 | 230.30 | 27.78 |
| Xiangxiangling | 989.47 | 1538.94 | 549.47 | 55.53 |
| Liangshan | 2898.68 | 2949.07 | 50.39 | 1.74 |
| Total | 8812.04 | 11,264.93 | 2452.89 | 27.84 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruan, T.; Wei, W.; Zhang, Z.; Zhou, H. Research on the Changes in Distribution and Habitat Suitability of the Chinese Red Panda Population. Animals 2024, 14, 424. https://doi.org/10.3390/ani14030424
Ruan T, Wei W, Zhang Z, Zhou H. Research on the Changes in Distribution and Habitat Suitability of the Chinese Red Panda Population. Animals. 2024; 14(3):424. https://doi.org/10.3390/ani14030424
Chicago/Turabian StyleRuan, Tao, Wei Wei, Zejun Zhang, and Hong Zhou. 2024. "Research on the Changes in Distribution and Habitat Suitability of the Chinese Red Panda Population" Animals 14, no. 3: 424. https://doi.org/10.3390/ani14030424
APA StyleRuan, T., Wei, W., Zhang, Z., & Zhou, H. (2024). Research on the Changes in Distribution and Habitat Suitability of the Chinese Red Panda Population. Animals, 14(3), 424. https://doi.org/10.3390/ani14030424