
Discrepancies between Genetic and Visual Coat Color Assignment in Sarcidano Horse

 , , , and
, , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Territory
2.2. Amplification and Mutations Dedection
Statistical Analysis
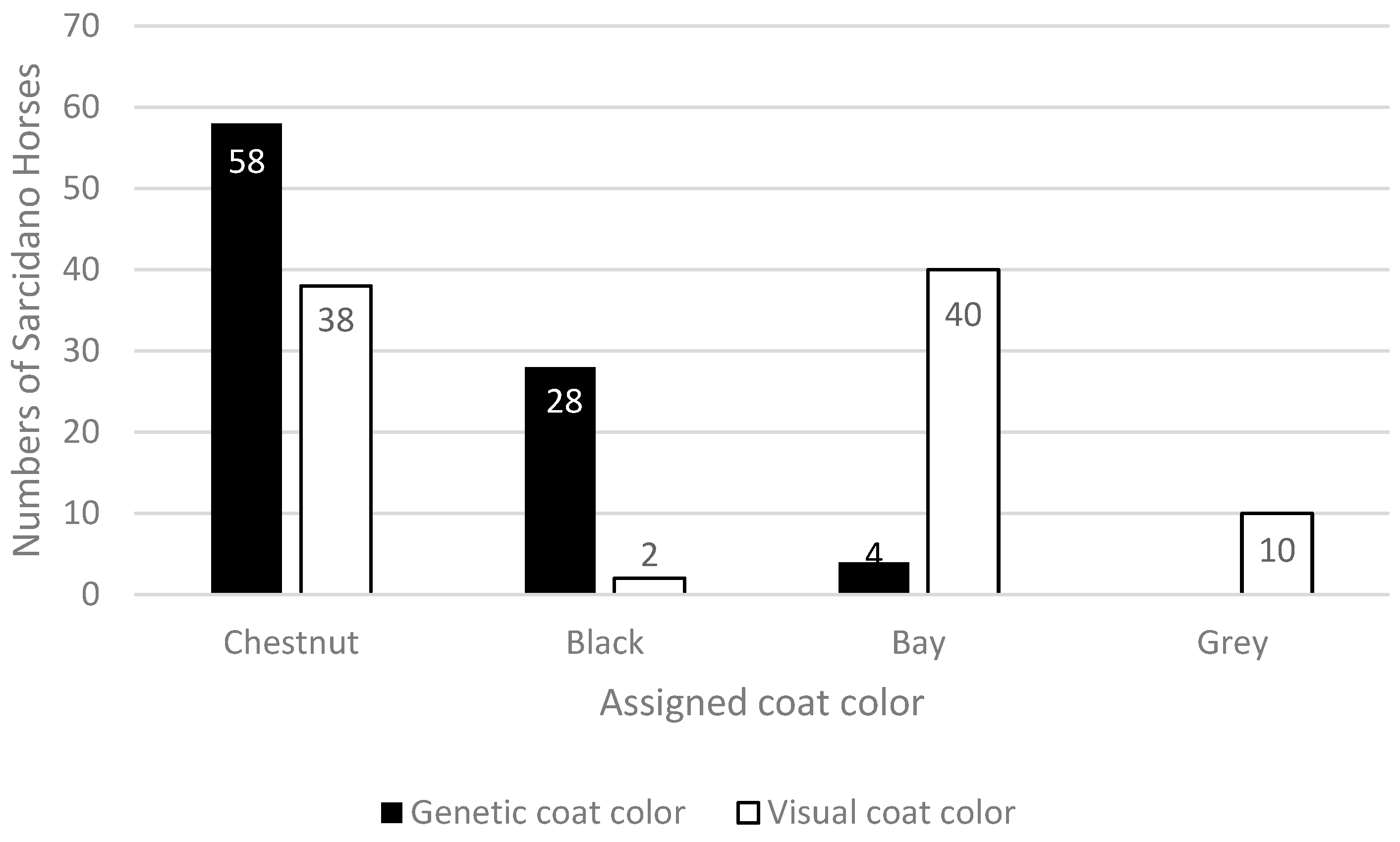
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ANAREAI. Available online: https://www.anareai.it/ (accessed on 15 September 2023).
- Morelli, L.; Useli, A.; Sanna, D.; Barbato, M.; Contu, D.; Pala, M.; Cancedda, M.; Francalacci, P. Mitochondrial DNA lineages of Italian Giara and Sarcidano horses. Genet. Mol. Res. 2014, 13, 8241–8257. [Google Scholar] [CrossRef]
- Cardinali, I.; Lancioni, H.; Giontella, A.; Capodiferro, M.R.; Capomaccio, S.; Buttazzoni, L.; Biggio, G.P.; Cherchi, R.; Albertini, E.; Olivieri, A.; et al. An overview of ten Italian horse breeds through mitochondrial DNA. PLoS ONE 2016, 7, e0153004. [Google Scholar] [CrossRef]
- Giontella, A.; Cardinali, I.; Pieramati, C.; Cherchi, R.; Biggio, G.P.; Achilli, A.; Silvestrelli, M.; Lancioni, H. A Genetic Window on Sardinian Native Horse Breeds through Uniparental Molecular Systems. Animals 2020, 10, 1544. [Google Scholar] [CrossRef]
- Teixeira, R.B.; Rendahl, A.K.; Anderson, S.M.; Mickelson, J.R.; Sigler, D.; Buchanan, B.R.; Coleman, R.J.; McCue, M.E. Coat color genotypes and risk and severity of melanoma in gray quarter horses. J. Vet. Intern. Med. 2013, 27, 1201–1208. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.X.; Chen, N.B.; Liu, W.J.; Li, R.; Lan, X.Y.; Chen, H.; Lei, C.Z.; Dang, R.H. Frequency of gray coat color in native Chinese horse breeds. Genet. Mol. Res. 2015, 30, 14144–14150. [Google Scholar] [CrossRef] [PubMed]
- Nowacka-Woszuk, J.; Mackowski, M.; Stefaniuk-Szmukier, M.; Cieslak, J. The equine graying with age mutation of the STX17 gene: A copy number study using droplet digital PCR reveals a new pattern. Anim. Genet. 2021, 52, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; He, X.; Zhao, Y.; Bai, D.; Shiraigo, W.; Zhao, Q.; Manglai, D. Regulatory pathway analysis of coat color genes in Mongolian horses. Hereditas 2017, 29, 13. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.P.; Schwengber, E.B.; Albrecht, F.F.; Isola, J.V.; van der Linden, L.D.S. Beyond fifty shades: The genetics of horse colors. In Trends and Advances in Veterinary Genetics; IntechOpen: London, UK, 2017. [Google Scholar]
- Sponenberg, D.P.; Bellone, R. Equine Color Genetics, 4th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2017; ISBN 978-1-119-13058-1. [Google Scholar]
- Beaumont, K.A.; Shekar, S.L.; Newton, R.A.; James, M.R.; Stow, J.L.; Duffy, D.L.; Sturm, R.A. Receptor function, dominant negative activity and phenotype correlations for MC1R variant alleles. Hum. Mol. Genet. 2007, 16, 2249–2260. [Google Scholar] [CrossRef] [PubMed]
- Shang, S.; Yu, Y.; Zhao, Y.; Dang, W.; Zhang, J.; Qin, X.; Irwin, D.M.; Qin Wang, Q.; Liu, F.; Wang, Z.; et al. Synergy between MC1R and ASIP for coat color in horses (Equus caballus). J. Anim. Sci. 2019, 97, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- Thiruvenkadan, A.K.; Kandasamy, N.; Panneerselvam, S. Coat colour inheritance in horses. Livest. Sci. 2008, 117, 109–129. [Google Scholar] [CrossRef]
- Cosso, G.; Carcangiu, V.; Luridiana, S.; Fiori, S.; Columbano, N.; Masala, G.; Careddu, G.M.; Sanna Passino, E.; Mura, M.C. Characterization of the Sarcidano Horse Coat Color Genes. Animals 2022, 12, 2677. [Google Scholar] [CrossRef] [PubMed]
- Mackowski, M.; Wodas, L.; Brooks, S.A.; Cieslak, J. TBX3 and ASIP genotypes reveal discrepancies in officially recorded coat colors of Hucul horses. Animal 2019, 13, 1811–1816. [Google Scholar] [CrossRef] [PubMed]
- Silva, I.L.S.; Junqueira, G.S.B.; Oliveira, C.A.A.; Costa, R.B.; De Camargo, G.M.F. Inconsistencies in horse coat color registration: A case study. J. Equine Sci. 2020, 31, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Cieslak, J.; Brooks, S.A.; Wodas, L.; Mantaj, W.; Borowska, A.; Sliwowska, J.H.; Ziarniak, K.; Mackowski, M. Genetic Background of the Polish Primitive Horse (Konik) Coat Color Variation-New Insight into Dun Dilution Phenotypic Effect. J. Hered. 2021, 112, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Cieslak, M.; Reissmann, M.; Hofreiter, M.; Lydwig, A. Colours of domestication. Biol. Rev. 2011, 86, 885–899. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, H.I.; Ahmad, M.J.; Jabbir, F.; Ahmar, S.; Ahmad, N.; Elokil, A.A.; Chen, J. The Domestication Makeup: Evolution, Survival, and Challenges. Front. Ecol. Evol. 2020, 8, 103. [Google Scholar] [CrossRef]
- Ludwig, A.; Pruvost, M.; Reissmann, M.; Benecke, N.; Brockmann, G.A.; Castaños, P.; Cieslak, M.; Lippold, S.; Llorente, L.; Malaspinas, A.S.; et al. Coat color variation at the beginning of horse domestication. Science 2009, 324, 485. [Google Scholar] [CrossRef]
- Reissmann, M.; Musa, L.; Zakizadeh, S.; Ludwig, A. Distribution of coat-color-associated alleles in the domestic horse population and Przewalski’s horse. J. Appl. Genet. 2016, 57, 519–525. [Google Scholar] [CrossRef]
- R Core Team 2023-Version 4.3.2 R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 30 October 2023).
- Marklund, L.; Moller, M.J.; Sandberg, K.; Andersson, L. A missense mutation in the gene for melanocyte-stimulating hormone receptor (MC1R) is associated with the chestnut coat color in horses. Mamm. Genome 1996, 7, 895–899. [Google Scholar] [CrossRef]
- Rieder, S.; Taourit, S.; Mariat, D.; Langlois, B.; Guerin, G. Mutations in the agouti (ASIP), the extension (MC1R), and the brown (TYRP1) loci and their association to coat color phenotypes in horses (Equus caballus). Mamm. Genome 2001, 12, 450–455. [Google Scholar] [CrossRef]
- Druml, T.; Grilz-Seger, G.; Horna, M.; Brem, G. Discriminant Analysis of Colour Measurements Reveals Allele Dosage Effect of ASIP/MC1R in Bay Horses. Czech J. Anim. Sci. 2018, 63, 347–355. [Google Scholar] [CrossRef]
- Corbin, L.J.; Pope, J.; Sanson, J.; Antczak, D.F.; Miller, D.; Sadeghi, R.; Brooks, S.A. An Independent Locus Upstream of ASIP Controls Variation in the Shade of the Bay Coat Colour in Horses. Genes 2020, 11, 606. [Google Scholar] [CrossRef]
- Avila, F.; Hughes, S.S.; Magdesian, K.G.; Penedo, M.C.T.; Bellone, R.R. Breed Distribution and Allele Frequencies of Base Coat Color, Dilution, and White Patterning Variants across 28 Horse Breeds. Genes 2022, 13, 1641. [Google Scholar] [CrossRef]
- Kavar, T.; Čeh, E.; Dovč, P. A simplified PCR-based method for detection of gray coat color allele in horse. Mol. Cell Probes. 2012, 26, 256–258. [Google Scholar] [CrossRef] [PubMed]
- Pielberg, R.G.; Golovko, A.; Sundström, E.; Curik, I.; Lennartsson, J.; Seltenhammer, M.H.; Druml, T.; Binns, M.; Fitzsimmons, C.; Lindgren, G.; et al. A cis-acting regulatory mutation causes premature hair graying and susceptibility to melanoma in the horse. Nat. Genet. 2008, 40, 1004–1009. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.Y.; Zhao, Y.P.; Li, B.; Bou, G.; Zhang, X.Z.; Mongke, T.; Bao, T.; Gereliin, S.; Gereltuuin, T.; Li, C.; et al. Overview of the genetic control of horse coat color patterns. Yi Chuan 2018, 40, 357–368. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| MC1R | ASIP | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotypes | Alleles | Genotypes | Alleles | ||||||||
| C/C | C/T | T/T | C | T | 102/102 | 102/91 | 91/91 | 102 | 91 | ||
| Coat color | Black | 5 | 23 | 0 | 33 | 23 | 0 | 0 | 28 | 0 | 56 |
| Bay | 0 | 4 | 0 | 4 | 4 | 0 | 4 | 0 | 4 | 4 | |
| Chestnut | 0 | 0 | 58 | 0 | 116 | 2 | 22 | 34 | 26 | 90 | |
| Genotypes | Visual Observation | ||
|---|---|---|---|
| MC1R | ASIP | Correct (%) | Incorrect (%) |
| C/C | 91/91 | 60.0 | 40.0 |
| T/C | 91/102 | 75.0 | 25.0 |
| T/C | 91/91 | 0.0 | 100.0 |
| T/T | 91/91 | 61.8 | 38.2 |
| T/T | 91/102 | 59.1 | 40.9 |
| T/T | 102/102 | 100.0 | 0.0 |
| Total | 46.6 | 53.4 | |
| MC1R Genotype | ||||
|---|---|---|---|---|
| C/C | C/T | T/T | ||
| ASIP genotype | 102/102 | 0 Bay | 0 Bay | 2 Chestnut |
| 102/91 | 0 Bay | 4 Bay | 22 Chestnut * | |
| 91/91 | 5 Black | 23 Black ** | 34 Chestnut *** | |
| Visual Coat Color (VCC) | ||||||
|---|---|---|---|---|---|---|
| Bay | Gray | Black | Chestnut | Total GCC | ||
| Genetic Coat Color (GCC) | Bay | 0.03 | 0.00 | 0.00 | 0.01 | 0.04 |
| Black | 0.28 | 0.00 | 0.02 | 0.01 | 0.31 | |
| Chestnut | 0.14 | 0.11 | 0.00 | 0.40 | 0.65 | |
| Total VCC | 0.45 | 0.11 | 0.02 | 0.42 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mura, M.C.; Carcangiu, V.; Cosso, G.; Columbano, N.; Sanna Passino, E.; Luridiana, S. Discrepancies between Genetic and Visual Coat Color Assignment in Sarcidano Horse. Animals 2024, 14, 543. https://doi.org/10.3390/ani14040543
Mura MC, Carcangiu V, Cosso G, Columbano N, Sanna Passino E, Luridiana S. Discrepancies between Genetic and Visual Coat Color Assignment in Sarcidano Horse. Animals. 2024; 14(4):543. https://doi.org/10.3390/ani14040543
Chicago/Turabian StyleMura, Maria Consuelo, Vincenzo Carcangiu, Giovanni Cosso, Nicolò Columbano, Eraldo Sanna Passino, and Sebastiano Luridiana. 2024. "Discrepancies between Genetic and Visual Coat Color Assignment in Sarcidano Horse" Animals 14, no. 4: 543. https://doi.org/10.3390/ani14040543
APA StyleMura, M. C., Carcangiu, V., Cosso, G., Columbano, N., Sanna Passino, E., & Luridiana, S. (2024). Discrepancies between Genetic and Visual Coat Color Assignment in Sarcidano Horse. Animals, 14(4), 543. https://doi.org/10.3390/ani14040543