
Evaluation of the Anti-Inflammatory and Antioxidant Potential of Cymbopogon citratus Essential Oil in Zebrafish

 , , , , , , and
, , , , , , and 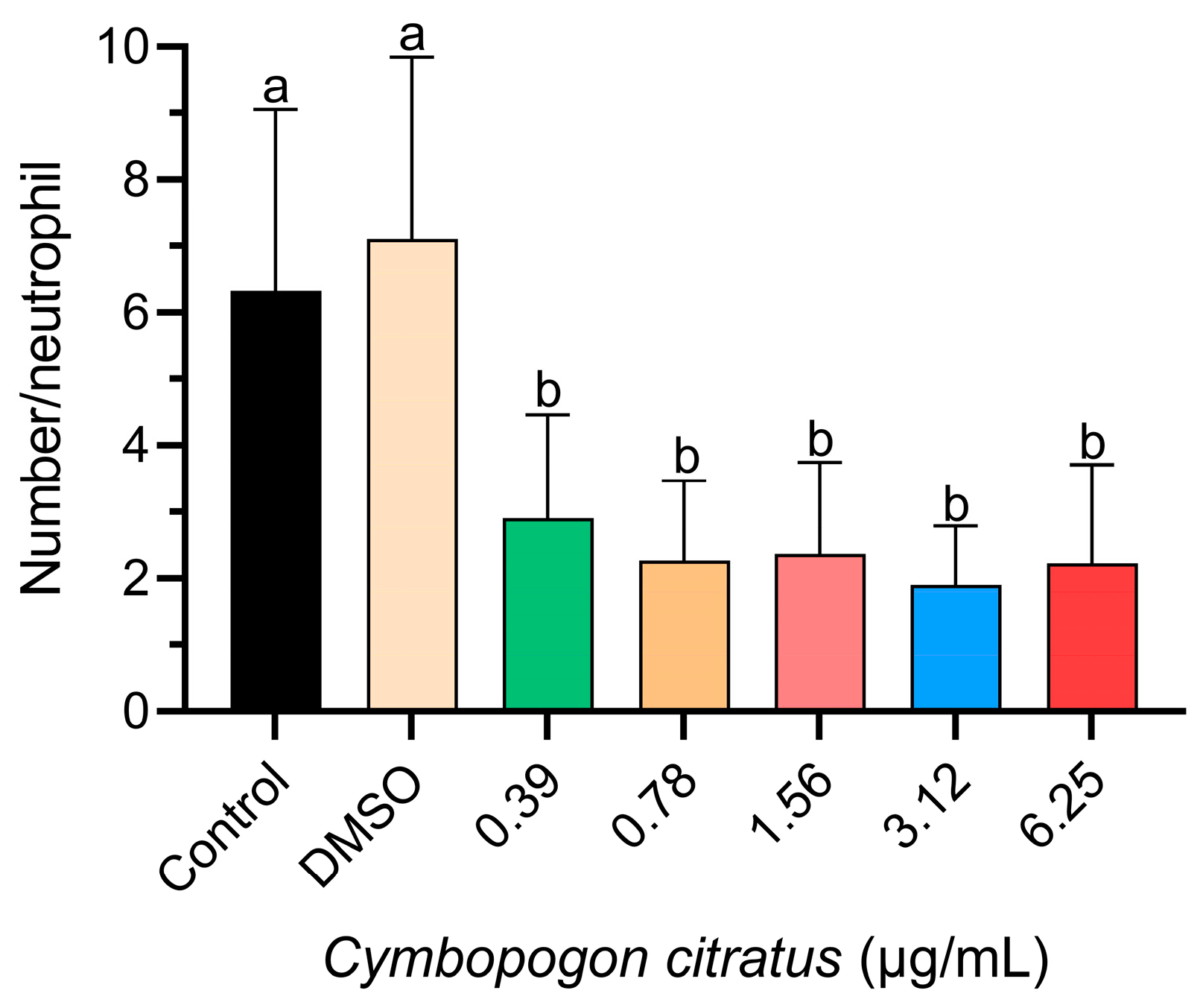{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material and Obtaining Essential Oil
2.3. Zebrafish Maintenance and Embryo Collection
2.4. Neutrophil Migration
2.5. Caudal Fin Regeneration
2.6. Antioxidant and Apoptotic Activity Induced by Hydrogen Peroxide (H2O2)
2.7. Statistical Analysis
3. Results
3.1. Neutrophil Migration
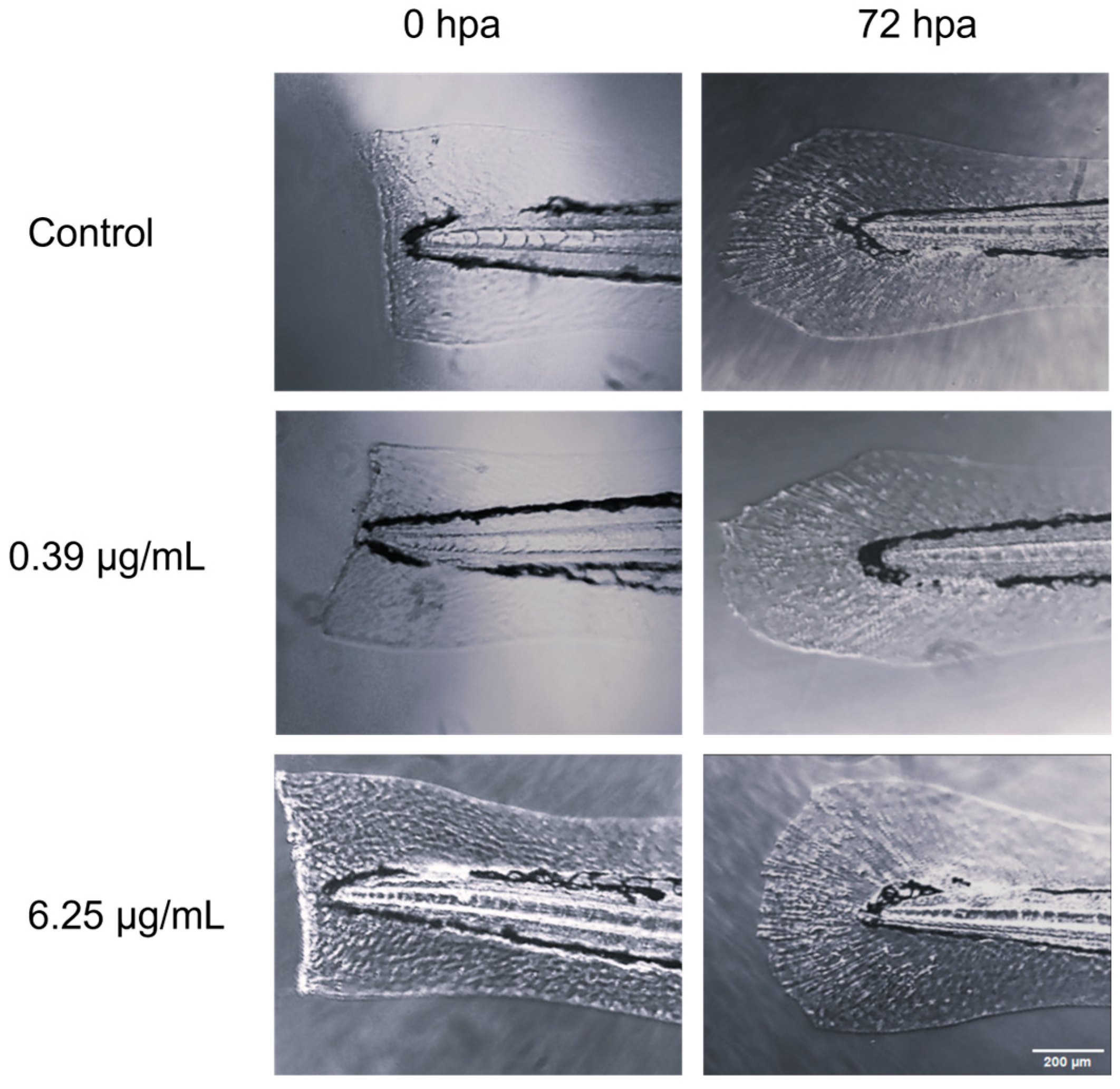
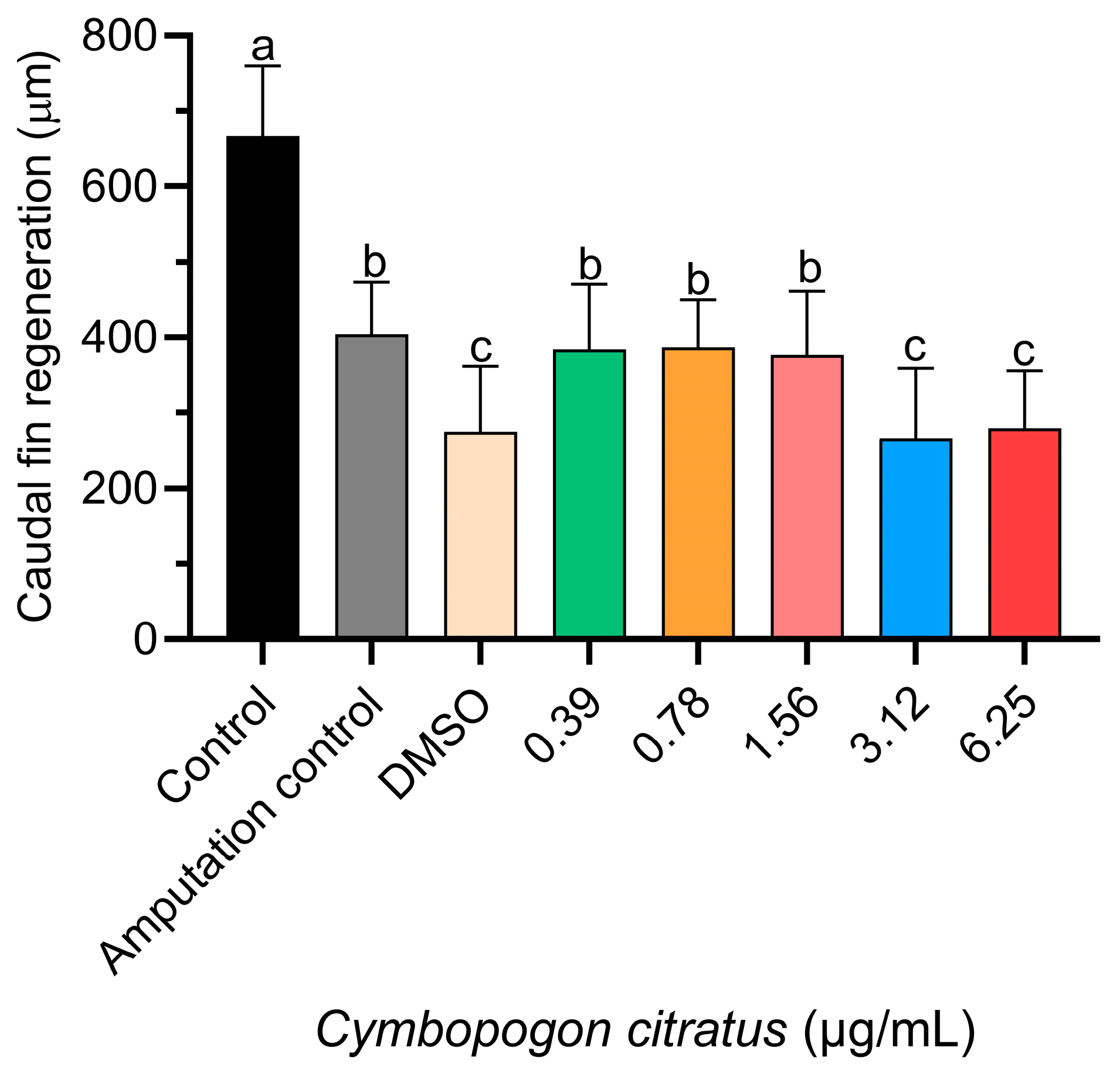
3.2. Tail Regeneration
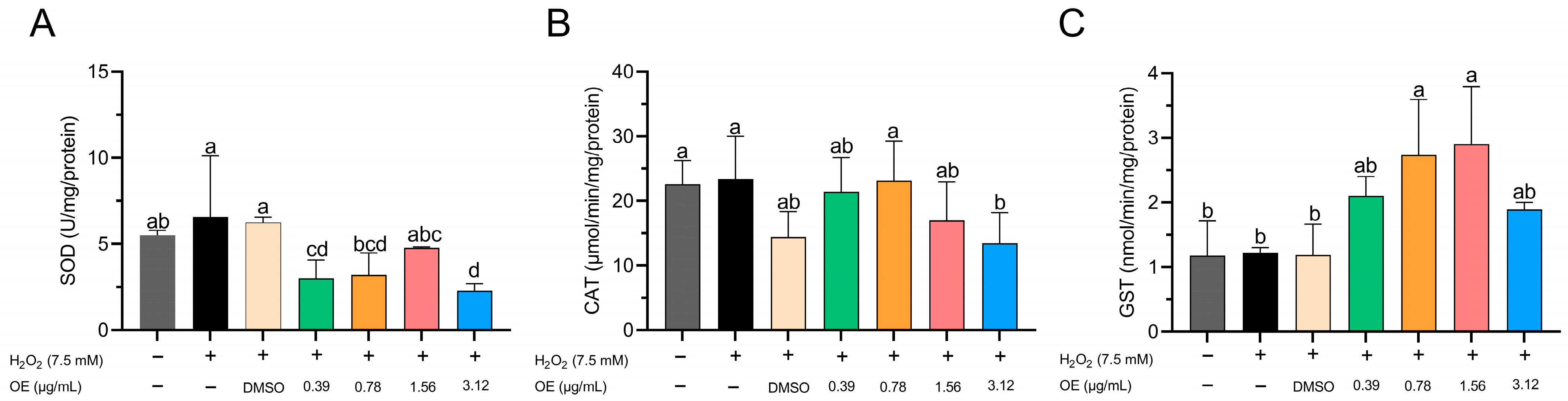
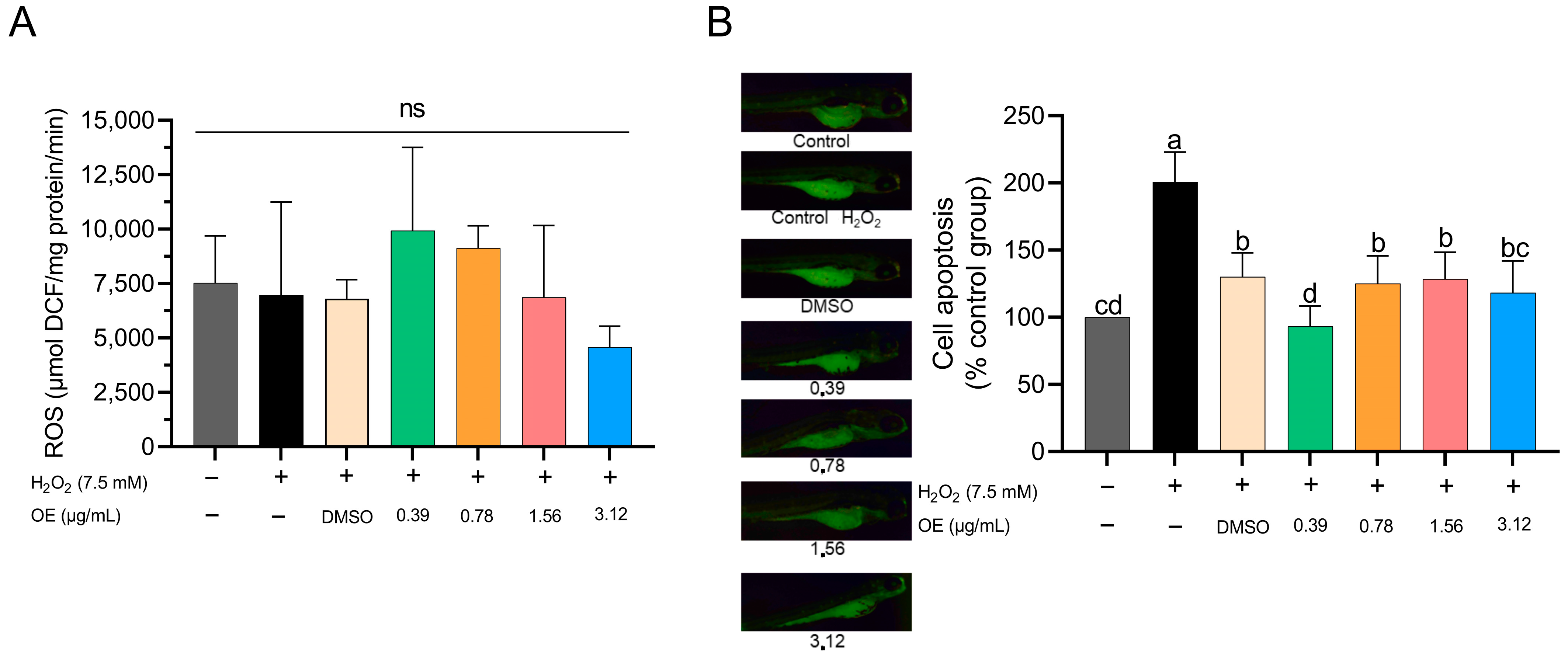
3.3. Antioxidant Activity Induced by H2O2
3.4. ROS and Apoptosis Induced by H2O2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rajčević, N.; Bukvički, D.; Dodoš, T.; Marin, P.D. Interactions between Natural Products—A Review. Metabolites 2022, 12, 1256. [Google Scholar] [CrossRef] [PubMed]
- Bruning, M.C.R.; Mosegui, G.B.G.; Vianna, C.M. A Utilização Da Fitoterapia e de Plantas Medicinais Em Unidades Básicas de Saúde Nos Municípios de Cascavel e Foz Do Iguaçu—Paraná: A Visão Dos Profissionais de Saúde. Ciência Saúde Coletiva 2012, 17, 2675–2685. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Paduan, R.H.; Gazin, Z.C.; Jacomassi, E.; D’Oliveira, P.S.; Cortez, D.A.G.; Cortez, L.E.R. Determinação Do Rendimento e Atividade Antimicrobiana Do Óleo Essencial de Cymbopogon citratus (DC.) Stapf Em Função de Sazonalidade e Consorciamento. Rev. Bras. Farmacogn. 2009, 19, 436–441. [Google Scholar] [CrossRef]
- Karami, S.; Yargholi, A.; Lamardi, S.N.S.; Soleymani, S.; Shirbeigi, L.; Rahimi, R. A Review of Ethnopharmacology, Phytochemistry and Pharmacology of Cymbopogon Species. Res. J. Pharmacogn. 2021, 8, 83–112. [Google Scholar]
- Nambiar, V.S.; Matela, H. Potential Functions of Lemon Grass (Cymbopogon citratus) in Health and Disease. Int. J. Pharm. Biol. Arch. 2012, 3, 1035–1043. [Google Scholar]
- Bolouri, P.; Salami, R.; Kouhi, S.; Kordi, M.; Asgari Lajayer, B.; Hadian, J.; Astatkie, T. Applications of Essential Oils and Plant Extracts in Different Industries. Molecules 2022, 27, 8999. [Google Scholar] [CrossRef] [PubMed]
- Lackmann, C.; Santos, M.M.; Rainieri, S.; Barranco, A.; Hollert, H.; Spirhanzlova, P.; Velki, M.; Seiler, T.-B. Novel Procedures for Whole Organism Detection and Quantification of Fluorescence as a Measurement for Oxidative Stress in Zebrafish (Danio rerio) Larvae. Chemosphere 2018, 197, 200–209. [Google Scholar] [CrossRef]
- Lintermans, L.L.; Stegeman, C.A.; Heeringa, P.; Abdulahad, W.H. T Cells in Vascular Inflammatory Diseases. Front. Immunol. 2014, 5, 504. [Google Scholar] [CrossRef]
- Biswas, S.K. Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxid. Med. Cell. Longev. 2016, 2016, 569831. [Google Scholar] [CrossRef]
- Li, F.S.; Weng, J.K. Demystifying Traditional Herbal Medicine with Modern Approach. Nat. Plants 2017, 3, 17109. [Google Scholar] [CrossRef]
- Stuart, K.A.; Welsh, K.; Walker, M.C.; Edrada-Ebel, R.A. Metabolomic Tools Used in Marine Natural Product Drug Discovery. Expert Opin. Drug Discov. 2020, 15, 499–522. [Google Scholar] [CrossRef] [PubMed]
- Aware, C.B.; Patil, D.N.; Suryawanshi, S.S.; Mali, P.R.; Rane, M.R.; Gurav, R.G.; Jadhav, J.P. Natural Bioactive Products as Promising Therapeutics: A Review of Natural Product-Based Drug Development. S. Afr. J. Bot. 2022, 151, 512–528. [Google Scholar] [CrossRef]
- Mizgirev, I.V.; Revskoy, S. A New Zebrafish Model for Experimental Leukemia Therapy. Cancer Biol. Ther. 2010, 9, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Chahardehi, A.M.; Arsad, H.; Lim, V. Zebrafish as a Successful Animal Model for Screening Toxicity of Medicinal Plants. Plants 2020, 9, 1345. [Google Scholar] [CrossRef] [PubMed]
- Teame, T.; Zhang, Z.; Ran, C.; Zhang, H.; Yang, Y.; Ding, Q.; Xie, M.; Gao, C.; Ye, Y.; Duan, M.; et al. The Use of Zebrafish (Danio rerio) as Biomedical Models. Anim. Front. 2019, 9, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Bambino, K.; Chu, J. Zebrafish in Toxicology and Environmental Health. Curr. Top. Dev. Biol. 2017, 124, 331. [Google Scholar] [CrossRef]
- Han, H.S.; Jang, G.H.; Jun, I.; Seo, H.; Park, J.; Glyn-Jones, S.; Seok, H.K.; Lee, K.H.; Mantovani, D.; Kim, Y.C.; et al. Transgenic Zebrafish Model for Quantification and Visualization of Tissue Toxicity Caused by Alloying Elements in Newly Developed Biodegradable Metal. Sci. Rep. 2018, 8, 13818. [Google Scholar] [CrossRef] [PubMed]
- Jayasinghe, C.D.; Jayawardena, U.A. Toxicity Assessment of Herbal Medicine Using Zebrafish Embryos: A Systematic Review. Evid. Based Complement. Altern. Med. 2019, 2019, 7272808. [Google Scholar] [CrossRef]
- Pham, D.H.; De Roo, B.; Nguyen, X.B.; Vervaele, M.; Kecskés, A.; Ny, A.; Copmans, D.; Vriens, H.; Locquet, J.P.; Hoet, P.; et al. Use of Zebrafish Larvae as a Multi-Endpoint Platform to Characterize the Toxicity Profile of Silica Nanoparticles. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- He, J.H.; Gao, J.M.; Huang, C.J.; Li, C.Q. Zebrafish Models for Assessing Developmental and Reproductive Toxicity. Neurotoxicol. Teratol. 2014, 42, 35–42. [Google Scholar] [CrossRef]
- Duarte da Silva, K.C.; do Carmo Rodrigues Virote, B.; de Fátima Santos, M.; Dias Castro, T.F.; de Azevedo Martins, M.S.; Carneiro, W.F.; Solis Murgas, L.D. Embriotoxic and Antioxidant Effects of Cymbopogon citratus Leaf Volatile Oil on Zebrafish. Rev. Bras. Farmacogn. 2023, 33, 778–789. [Google Scholar] [CrossRef]
- Hasegawa, T.; Hall, C.J.; Crosier, P.S.; Abe, G.; Kawakami, K.; Kudo, A.; Kawakami, A. Transient Inflammatory Response Mediated by Interleukin-1β Is Required for Proper Regeneration in Zebrafish Fin Fold. eLife 2017, 6, e22716. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, A.; Fukazawa, T.; Takeda, H. Early Fin Primordia of Zebrafish Larvae Regenerate by a Similar Growth Control Mechanism with Adult Regeneration. Dev. Dyn. 2004, 231, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Polednik, K.M.; Koch, A.C.; Felzien, L.K. Effects of Essential Oil from Thymus Vulgaris on Viability and Inflammation in Zebrafish Embryos. Zebrafish 2018, 15, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Le Guyader, D.; Redd, M.J.; Colucci-Guyon, E.; Murayama, E.; Kissa, K.; Briolat, V.; Mordelet, E.; Zapata, A.; Shinomiya, H.; Herbomel, P. Origins and Unconventional Behavior of Neutrophils in Developing Zebrafish. Blood 2008, 111, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Walters, K.B.; Dodd, M.E.; Mathias, J.R.; Gallagher, A.J.; Bennin, D.A.; Rhodes, J.; Kanki, J.P.; Look, A.T.; Grinblat, Y.; Huttenlocher, A. Muscle Degeneration and Leukocyte Infiltration Caused by Mutation of Zebrafish Fad24. Dev. Dyn. 2009, 238, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Gu, L.; Tan, H.; Liu, P.; Gao, G.; Tian, L.; Chen, H.; Lu, T.; Qian, H.; Fu, Z.; et al. Effects of 17α-ethinylestradiol on Caudal Fin Regeneration in Zebrafish Larvae. Sci. Total Environ. 2019, 653, 10–22. [Google Scholar] [CrossRef]
- Kim, S.; Kim, M.; Kang, M.C.; Lee, H.H.L.; Cho, C.H.; Choi, I.; Park, Y.; Lee, S.H. Antioxidant Effects of Turmeric Leaf Extract against Hydrogen Peroxide-Induced Oxidative Stress In Vitro in Vero Cells and In Vivo in Zebrafish. Antioxidants 2021, 10, 112. [Google Scholar] [CrossRef]
- Jayawardena, T.U.; Wang, L.; Asanka Sanjeewa, K.K.; Kang, S.; Lee, J.S.; Jeon, Y.J. Antioxidant Potential of Sulfated Polysaccharides from Padina boryana; Protective Effect against Oxidative Stress in In Vitro and In Vivo Zebrafish Model. Mar. Drugs 2020, 18, 212. [Google Scholar] [CrossRef]
- Jin, S.J.; Piao, S.; Cha, Y.N.; Kim, C. Taurine Chloramine Activates Nrf2, Increases HO-1 Expression and Protects Cells from Death Caused by Hydrogen Peroxide. J. Clin. Biochem. Nutr. 2009, 45, 37–43. [Google Scholar] [CrossRef]
- Kang, M.C.; Kim, S.Y.; Kim, E.A.; Lee, J.H.; Kim, Y.S.; Yu, S.K.; Chae, J.B.; Choe, I.H.; Cho, J.H.; Jeon, Y.J. Antioxidant Activity of Polysaccharide Purified from Acanthopanax koreanum Nakai Stems in Vitro and in Vivo Zebrafish Model. Carbohydr. Polym. 2015, 127, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Hydrogen Peroxide as a Central Redox Signaling Molecule in Physiological Oxidative Stress: Oxidative Eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.; Félix, L.; Luzio, A.; Parra, S.; Cabecinha, E.; Bellas, J.; Monteiro, S.M. Toxicological Effects Induced on Early Life Stages of Zebrafish (Danio rerio) after an Acute Exposure to Microplastics Alone or Co-Exposed with Copper. Chemosphere 2020, 261, 127748. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.; Deng, Y.; Cao, Z.; Liao, X.; Zhang, J.; Lu, H. The Hepatoprotective Effects of Salvia plebeia R. Br. Extract in Zebrafish (Danio rerio). Fish Shellfish Immunol. 2019, 95, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Claiborne, A. Catalase Activity. In Handbook of Methods for Oxygen Radical Research; Greenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 283–284. [Google Scholar]
- Song, Y.; Zhu, L.S.; Wang, J.; Wang, J.H.; Liu, W.; Xie, H. DNA Damage and Effects on Antioxidative Enzymes in Earthworm (Eisenia foetida) Induced by Atrazine. Soil Biol. Biochem. 2009, 41, 905–909. [Google Scholar] [CrossRef]
- Gu, J.; Zhang, J.; Chen, Y.; Wang, H.; Guo, M.; Wang, L.; Wang, Z.; Wu, S.; Shi, L.; Gu, A.; et al. Neurobehavioral Effects of Bisphenol S Exposure in Early Life Stages of Zebrafish Larvae (Danio rerio). Chemosphere 2019, 217, 629–635. [Google Scholar] [CrossRef]
- Habig, W.H.; Jakoby, W.B. Assays for Differentiation of Glutathione S-Transferases. Methods Enzym. 1981, 77, 398–405. [Google Scholar] [CrossRef]
- Velki, M.; Lackmann, C.; Barranco, A.; Ereño Artabe, A.; Rainieri, S.; Hollert, H.; Seiler, T.B. Pesticides Diazinon and Diuron Increase Glutathione Levels and Affect Multixenobiotic Resistance Activity and Biomarker Responses in Zebrafish (Danio rerio) Embryos and Larvae. Environ. Sci. Eur. 2019, 31, 1–18. [Google Scholar] [CrossRef]
- Xia, Q.; Luo, J.; Mei, X.; Wang, Y.; Huang, W.; Wang, J.; Yang, R.; Ma, Z.; Lin, R. A Developmental Toxicity Assay of Carpesii Fructus on Zebrafish Embryos/Larvae. Toxicol. Res. 2017, 6, 460–467. [Google Scholar] [CrossRef]
- Zhao, C.; Jia, Z.; Li, E.; Zhao, X.; Han, T.; Tian, J.; Li, F.; Zou, D.; Lin, R. Hepatotoxicity Evaluation of Euphorbia kansui on Zebrafish Larvae in Vivo. Phytomedicine 2019, 62, 152959. [Google Scholar] [CrossRef] [PubMed]
- Driver, A.S.; Kodavanti, P.R.S.; Mundy, W.R. Age-Related Changes in Reactive Oxygen Species Production in Rat Brain Homogenates. Neurotoxicol. Teratol. 2000, 22, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Cruvinel, W.D.; Mesquita Júnior, D.; Araújo, J.A.; Catelan, T.T.; Souza, A.W.; Silva, N.P.; Andrade, L.E. Immune System: Part I. Fundamentals of Innate Immunity with Emphasis on Molecular and Cellular Mechanisms of Inflammatory Response. Rev. Bras. Reum. 2010, 50, 434–447. [Google Scholar] [CrossRef]
- Abbas, A.; Lichtman, A.; Pillai, S. Cellular and Molecular Immunology E-Book; Elsevier Health Sciences: Philadelphia, PA, USA, 2014; ISBN 0323286453. [Google Scholar]
- Freire, M.O.; Van Dyke, T.E. Natural Resolution of Inflammation. Periodontology 2000 2013, 63, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, K.; Andón, F.T.; El-Sayed, R.; Fadeel, B. Mechanisms of Carbon Nanotube-Induced Toxicity: Focus on Pulmonary Inflammation. Adv. Drug Deliv. Rev. 2013, 65, 2087–2097. [Google Scholar] [CrossRef] [PubMed]
- Muralidharan, S.; Mandrekar, P. Cellular Stress Response and Innate Immune Signaling: Integrating Pathways in Host Defense and Inflammation. J. Leukoc. Biol. 2013, 94, 1167–1184. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Kubes, P. Neutrophil Recruitment and Function in Health and Inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Halima, M.; Xie, Y.; Schaaf, M.J.M.; Meijer, A.H.; Wang, M. Ginsenoside Rg1 Acts as a Selective Glucocorticoid Receptor Agonist with Anti-Inflammatory Action without Affecting Tissue Regeneration in Zebrafish Larvae. Cells 2020, 9, 1107. [Google Scholar] [CrossRef]
- Yang, Y.; Zheng, K.; Mei, W.; Wang, Y.; Yu, C.; Yu, B.; Deng, S.; Hu, J. Anti-Inflammatory and Proresolution Activities of Bergapten Isolated from the Roots of Ficus hirta in an in Vivo Zebrafish Model. Biochem. Biophys. Res. Commun. 2018, 496, 763–769. [Google Scholar] [CrossRef]
- Leite, B.L.S.; Bonfim, R.R.; Antoniolli, A.R.; Thomazzi, S.M.; Araújo, A.A.S.; Blank, A.F.; Estevam, C.S.; Cambui, E.V.F.; Bonjardim, L.R.; Albuquerque Júnior, R.L.C.; et al. Assessment of Antinociceptive, Anti-Inflammatory and Antioxidant Properties of Cymbopogon winterianus Leaf Essential Oil. Pharm. Biol. 2010, 48, 1164–1169. [Google Scholar] [CrossRef]
- Loram, L.C.; Fuller, A.; Fick, L.G.; Cartmell, T.; Poole, S.; Mitchell, D. Cytokine Profiles during Carrageenan-Induced Inflammatory Hyperalgesia in Rat Muscle and Hind Paw. J. Pain 2007, 8, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Pérez, G.S.; Zavala, S.M.; Arias, G.L.; Ramos, L.M. Anti-Inflammatory Activity of Some Essential Oils. J. Essent. Oil Res. 2011, 23, 38–44. [Google Scholar] [CrossRef]
- Emílio-Silva, M.T.; Rodrigues, V.P.; Bueno, G.; Ohara, R.; Martins, M.G.; Horta-Júnior, J.A.C.; Branco, L.G.S.; Rocha, L.R.M.; Hiruma-Lima, C.A. Hypothermic Effect of Acute Citral Treatment during LPS-Induced Systemic Inflammation in Obese Mice: Reduction of Serum TNF-α and Leptin Levels. Biomolecules 2020, 10, 1454. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.T.; Chen, C.J.; Lin, T.Y.; Tung, J.C.; Wang, S.Y. Anti-Inflammation Activity of Fruit Essential Oil from Cinnamomum insularimontanum Hayata. Bioresour. Technol. 2008, 99, 8783–8787. [Google Scholar] [CrossRef] [PubMed]
- Abe, S.; Maruyama, N.; Hayama, K.; Ishibashi, H.; Inoue, S.; Oshima, H.; Yamaguchi, H. Suppression of Tumor Necrosis Factor-Alpha-Induced Neutrophil Adherence Responses by Essential Oils. Mediat. Inflamm. 2003, 12, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Boukhatem, M.N.; Ferhat, M.A.; Kameli, A.; Saidi, F.; Kebir, H.T. Lemon Grass (Cymbopogon citratus) Essential Oil as a Potent Anti-Inflammatory and Antifungal Drugs. Libyan J. Med. 2014, 9, 25431. [Google Scholar] [CrossRef] [PubMed]
- Gemberling, M.; Bailey, T.J.; Hyde, D.R.; Poss, K.D. The Zebrafish as a Model for Complex Tissue Regeneration. Trends Genet. 2013, 29, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Singulani, J.L.; Pedroso, R.S.; Ribeiro, A.B.; Nicolella, H.D.; Freitas, K.S.; Damasceno, J.L.; Vieira, T.M.; Crotti, A.E.M.; Tavares, D.C.; Martins, C.H.G.; et al. Geraniol and Linalool Anticandidal Activity, Genotoxic Potential and Embryotoxic Effect on Zebrafish. Future Microbiol. 2018, 13, 1637–1646. [Google Scholar] [CrossRef]
- Souza, A.C.S.; Silva, L.K.; Queiroz, T.B.; Marques, E.S.; Hiruma-Lima, C.A.; Gaivão, I.O.M.; Maistro, E.L. Citral Presents Cytotoxic and Genotoxic Effects in Human Cultured Cells. Drug Chem. Toxicol. 2020, 43, 435–440. [Google Scholar] [CrossRef]
- Plata-Rueda, A.; Rolim, G.D.S.; Wilcken, C.F.; Zanuncio, J.C.; Serrão, J.E.; Martínez, L.C. Acute Toxicity and Sublethal Effects of Lemongrass Essential Oil and Their Components against the Granary Weevil, Sitophilus granarius. Insects 2020, 11, 379. [Google Scholar] [CrossRef]
- Gudas, L.J. Emerging Roles for Retinoids in Regeneration and Differentiation in Normal and Disease States. Biochim. Biophys. Acta 2012, 1821, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Mathew, L.K.; Sengupta, S.; Franzosa, J.A.; Perry, J.; La Du, J.; Andreasen, E.A.; Tanguay, R.L. Comparative Expression Profiling Reveals an Essential Role for Raldh2 in Epimorphic Regeneration. J. Biol. Chem. 2009, 284, 33642–33653. [Google Scholar] [CrossRef] [PubMed]
- Viera-Vera, J.; García-Arrarás, J.E. Retinoic Acid Signaling Is Associated with Cell Proliferation, Muscle Cell Dedifferentiation, and Overall Rudiment Size during Intestinal Regeneration in the Sea Cucumber, Holothuria glaberrima. Biomolecules 2019, 9, 873. [Google Scholar] [CrossRef] [PubMed]
- Scadding, S.R. Citral, an Inhibitor of Retinoic Acid Synthesis, Modifies Pattern Formation during Limb Regeneration in the Axolotl Ambystoma mexicanum. Can. J. Zool. 2011, 77, 1835–1837. [Google Scholar] [CrossRef]
- Kaneko, N.; Katsuyama, Y.; Kawamura, K.; Fujiwara, S. Regeneration of the Gut Requires Retinoic Acid in the Budding Ascidian Polyandrocarpa misakiensis. Dev. Growth Differ. 2010, 52, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Gulcin, İ. Antioxidants and Antioxidant Methods: An Updated Overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, I. Antioxidant Activity of Food Constituents: An Overview. Arch. Toxicol. 2012, 86, 345–391. [Google Scholar] [CrossRef]
- Neha, K.; Haider, M.R.; Pathak, A.; Yar, M.S. Medicinal Prospects of Antioxidants: A Review. Eur. J. Med. Chem. 2019, 178, 687–704. [Google Scholar] [CrossRef]
- Işlekel, S.; Işlekel, H.; Güner, G.; Özdamar, N. Alterations in Superoxide Dismutase, Glutathione Peroxidase and Catalase Activities in Experimental Cerebral Ischemia-Reperfusion. Res. Exp. Med. 1999, 199, 167–176. [Google Scholar] [CrossRef]
- Nita, D.A.I.; Nita, V.; Spulber, S.; Moldovan, M.; Popa, D.P.; Zagrean, A.M.; Zagrean, L. Oxidative Damage Following Cerebral Ischemia Depends on Reperfusion—A Biochemical Study in Rat. J. Cell. Mol. Med. 2001, 5, 163–170. [Google Scholar] [CrossRef]
- Taheri Mirghaed, A.; Paknejad, H.; Mirzargar, S.S. Hepatoprotective Effects of Dietary Artemisia (Artemisia annua) Leaf Extract on Common Carp (Cyprinus carpio) Exposed to Ambient Ammonia. Aquaculture 2020, 527, 735443. [Google Scholar] [CrossRef]
- Wu, M.; Xu, H.; Shen, Y.; Qiu, W.; Yang, M. Oxidative Stress in Zebrafish Embryos Induced by Short-Term Exposure to Bisphenol A, Nonylphenol, and Their Mixture. Environ. Toxicol. Chem. 2011, 30, 2335–2341. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Gupta, S. The Multifaceted Role of Glutathione S-Transferases in Cancer. Cancer Lett. 2018, 433, 33–42. [Google Scholar] [CrossRef]
- Dringen, R.; Gutterer, J.M.; Hirrlinger, J. Glutathione Metabolism in Brain Metabolic Interaction between Astrocytes and Neurons in the Defense against Reactive Oxygen Species. Eur. J. Biochem. 2000, 267, 4912–4916. [Google Scholar] [CrossRef] [PubMed]
- Glisic, B.; Mihaljevic, I.; Popovic, M.; Zaja, R.; Loncar, J.; Fent, K.; Kovacevic, R.; Smital, T. Characterization of Glutathione-S-Transferases in Zebrafish (Danio rerio). Aquat. Toxicol. 2015, 158, 50–62. [Google Scholar] [CrossRef]
- Andrade, B.F.M.T.; Braga, C.P.; Dos Santos, K.C.; Barbosa, L.N.; Rall, V.L.M.; Sforcin, J.M.; Fernandes, A.A.H.; Fernandes Júnior, A. Effect of Inhaling Cymbopogon martinii Essential Oil and Geraniol on Serum Biochemistry Parameters and Oxidative Stress in Rats. Biochem. Res. Int. 2014, 2014, 493183. [Google Scholar] [CrossRef] [PubMed]
- Bicas, J.L.; Neri-Numa, I.A.; Ruiz, A.L.T.G.; De Carvalho, J.E.; Pastore, G.M. Evaluation of the Antioxidant and Antiproliferative Potential of Bioflavors. Food Chem. Toxicol. 2011, 49, 1610–1615. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.G. Antioxidant Activity of Medicinal and Aromatic Plants. A Review. Flavour. Fragr. J. 2010, 25, 291–312. [Google Scholar] [CrossRef]
- Fadel, H.; Marx, F.; El-Sawy, A.; El-Ghorab, A. Effect of Extraction Techniques on the Chemical Composition and Antioxidant Activity of Eucalyptus Camaldulensis Var. Brevirostris Leaf Oils. Eur. Food Res. Technol. 1999, 208, 212–216. [Google Scholar] [CrossRef]
- Graßmann, J. Terpenoids as Plant Antioxidants. Vitam. Horm. 2005, 72, 505–535. [Google Scholar] [CrossRef]
- Buch, P.; Patel, V.; Ranpariya, V.; Sheth, N.; Parmar, S. Neuroprotective Activityof Cymbopogon martinii against Cerebral Ischemia/Reperfusion-Induced Oxidative Stress in Rats. J. Ethnopharmacol. 2012, 142, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Tayeboon, G.S.; Tavakoli, F.; Hassani, S.; Khanavi, M.; Sabzevari, O.; Ostad, S.N. Effects of Cymbopogon citratus and Ferula assa-foetida Extracts on Glutamate-Induced Neurotoxicity. Vitr. Cell. Dev. Biol. Anim. 2013, 49, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.M.; Hua, K.F.; Lin, Y.C.; Chen, A.; Chang, J.M.; Kuoping Chao, L.; Ho, C.L.; Ka, S.M. Citral Is Renoprotective for Focal Segmental Glomerulosclerosis by Inhibiting Oxidative Stress and Apoptosis and Activating Nrf2 Pathway in Mice. PLoS ONE 2013, 8, e74871. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duarte da Silva, K.C.; Carneiro, W.F.; Virote, B.d.C.R.; Santos, M.d.F.; de Oliveira, J.P.L.; Castro, T.F.D.; Bertolucci, S.K.V.; Murgas, L.D.S. Evaluation of the Anti-Inflammatory and Antioxidant Potential of Cymbopogon citratus Essential Oil in Zebrafish. Animals 2024, 14, 581. https://doi.org/10.3390/ani14040581
Duarte da Silva KC, Carneiro WF, Virote BdCR, Santos MdF, de Oliveira JPL, Castro TFD, Bertolucci SKV, Murgas LDS. Evaluation of the Anti-Inflammatory and Antioxidant Potential of Cymbopogon citratus Essential Oil in Zebrafish. Animals. 2024; 14(4):581. https://doi.org/10.3390/ani14040581
Chicago/Turabian StyleDuarte da Silva, Kiara Cândido, William Franco Carneiro, Bárbara do Carmo Rodrigues Virote, Maria de Fátima Santos, João Paulo Lima de Oliveira, Tássia Flávia Dias Castro, Suzan Kelly Vilela Bertolucci, and Luis David Solis Murgas. 2024. "Evaluation of the Anti-Inflammatory and Antioxidant Potential of Cymbopogon citratus Essential Oil in Zebrafish" Animals 14, no. 4: 581. https://doi.org/10.3390/ani14040581
APA StyleDuarte da Silva, K. C., Carneiro, W. F., Virote, B. d. C. R., Santos, M. d. F., de Oliveira, J. P. L., Castro, T. F. D., Bertolucci, S. K. V., & Murgas, L. D. S. (2024). Evaluation of the Anti-Inflammatory and Antioxidant Potential of Cymbopogon citratus Essential Oil in Zebrafish. Animals, 14(4), 581. https://doi.org/10.3390/ani14040581