
Assessing the Distribution and Richness of Mammalian Species Using a Stacking Species Distribution Model in a Temperate Forest


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
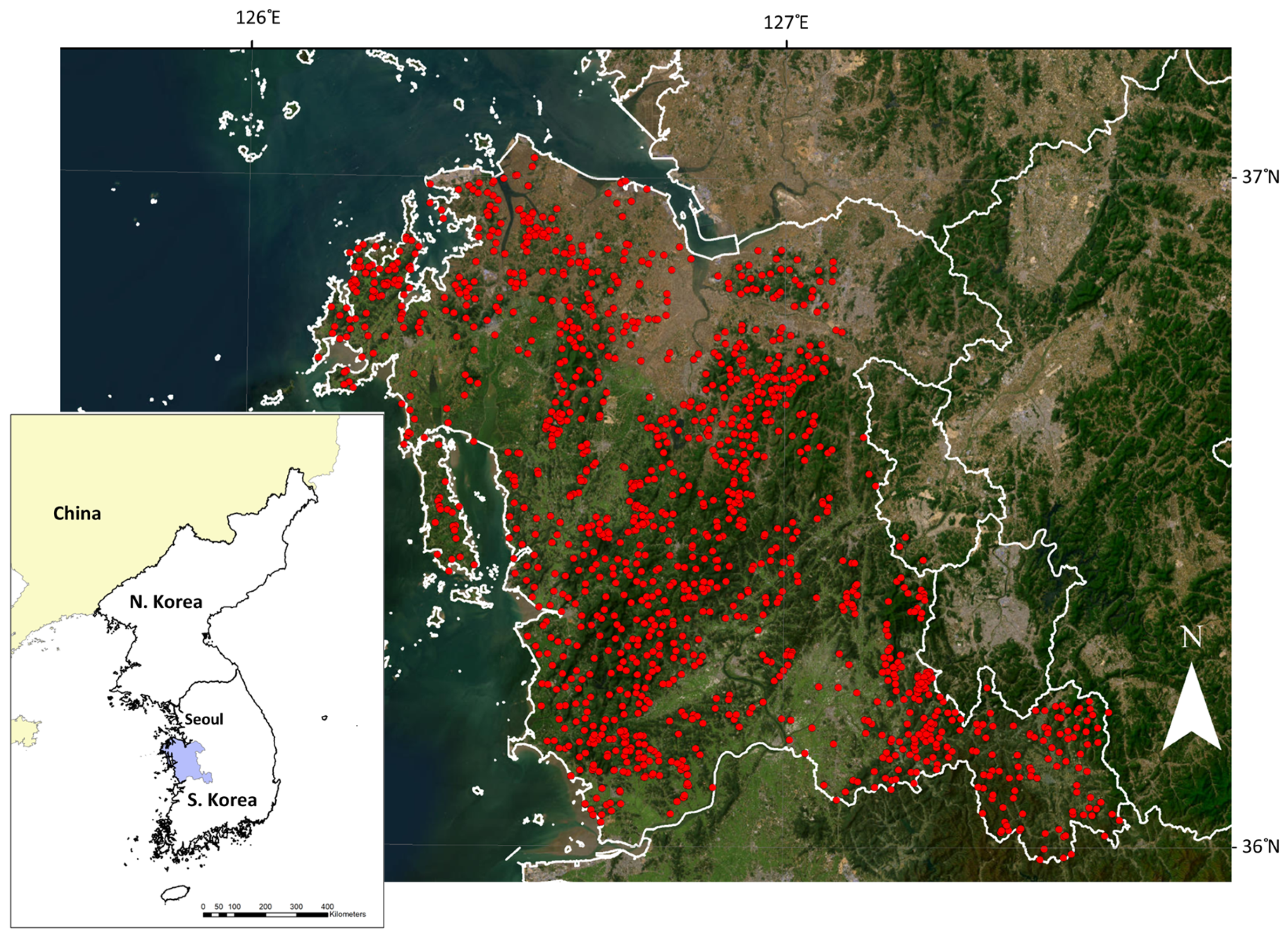
2.1. Study Area
2.2. Data and Analysis
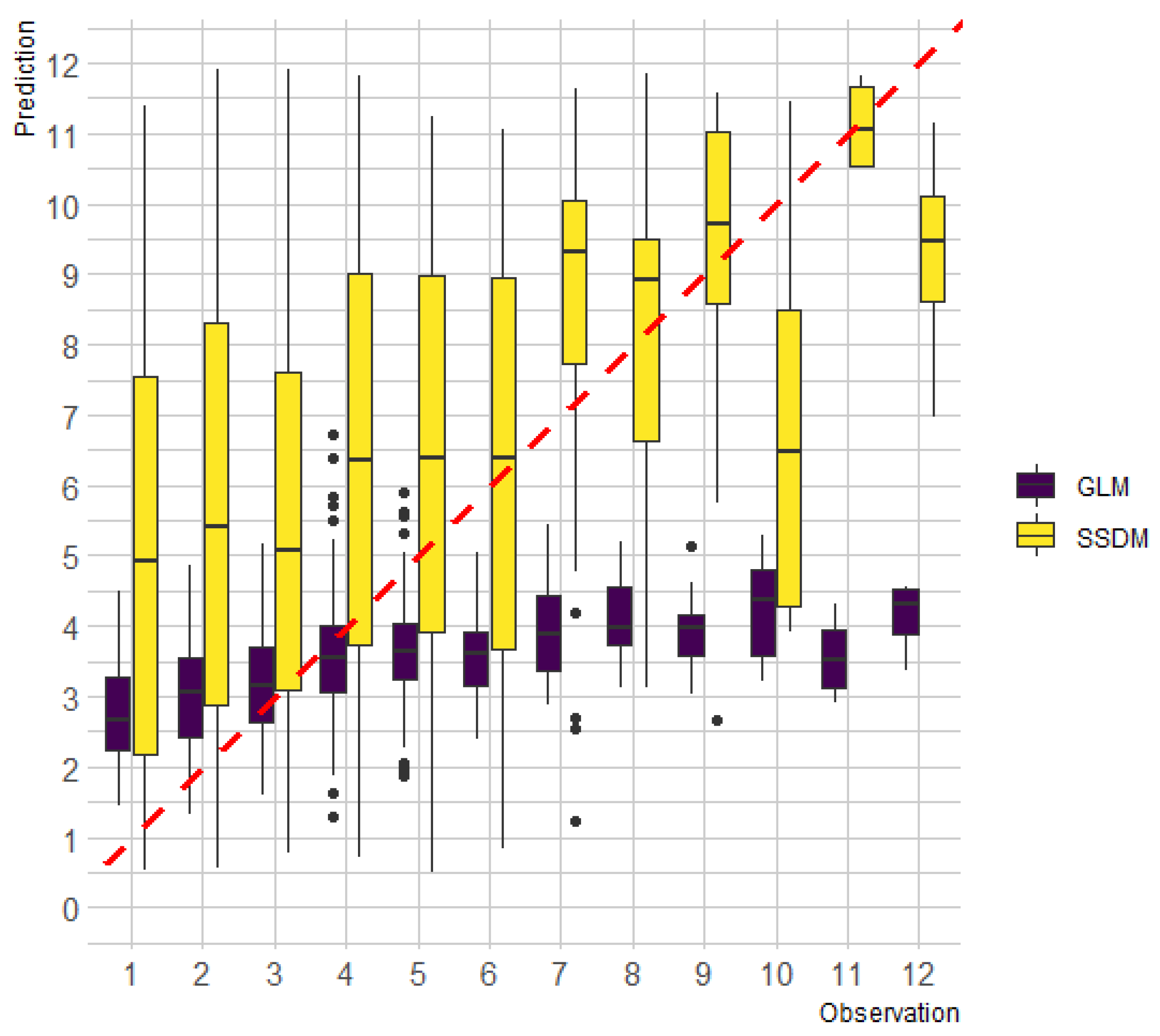
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buisson, L.; Thuiller, W.; Casajus, N.; Lek, S.; Grenouillet, G. Uncertainty in Ensemble Forecasting of Species Distribution. Glob. Chang. Biol. 2010, 16, 1145–1157. [Google Scholar] [CrossRef]
- Thuiller, W. BIOMOD–Optimizing Predictions of Species Distributions and Projecting Potential Future Shifts under Global Change. Glob. Chang. Biol. 2003, 9, 1353–1362. [Google Scholar] [CrossRef]
- Calabrese, J.M.; Certain, G.; Kraan, C.; Dormann, C.F. Stacking Species Distribution Models and Adjusting Bias by Linking Them to Macroecological Models. Glob. Ecol. Biogeogr. 2014, 23, 99–112. [Google Scholar] [CrossRef]
- Pecchi, M.; Marchi, M.; Burton, V.; Giannetti, F.; Moriondo, M.; Bernetti, I.; Bindi, M.; Chirici, G. Species Distribution Modelling to Support Forest Management. A Literature Review. Ecol. Model. 2019, 411, 108817. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Guillera-Arroita, G.; Lahoz-Monfort, J.J. A Review of Evidence about Use and Performance of Species Distribution Modelling Ensembles like BIOMOD. Divers. Distrib. 2019, 25, 839–852. [Google Scholar] [CrossRef]
- Elith, J.H.; Graham, C.P.; Anderson, R.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel Methods Improve Prediction of Species’ Distributions from Occurrence Data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Segurado, P.; Araújo, M.B. An Evaluation of Methods for Modelling Species Distributions. J. Biogeogr. 2004, 31, 1555–1568. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble Forecasting of Species Distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Ferrier, S.; Guisan, A. Spatial Modelling of Biodiversity at the Community Level. J. Appl. Ecol. 2006, 43, 393–404. [Google Scholar] [CrossRef]
- Mateo, R.G.; Felicísimo, Á.M.; Pottier, J.; Guisan, A.; Muñoz, J. Do Stacked Species Distribution Models Reflect Altitudinal Diversity Patterns? PLoS ONE 2012, 7, e32586. [Google Scholar] [CrossRef]
- Biber, M.F.; Voskamp, A.; Niamir, A.; Hickler, T.; Hof, C. A Comparison of Macroecological and Stacked Species Distribution Models to Predict Future Global Terrestrial Vertebrate Richness. J. Biogeogr. 2020, 47, 114–129. [Google Scholar] [CrossRef]
- Schmitt, S.; Pouteau, R.; Justeau, D.; de Boissieu, F.; Birnbaum, P. SSDM: An r Package to Predict Distribution of Species Richness and Composition Based on Stacked Species Distribution Models. Methods Ecol. Evol. 2017, 8, 1795–1803. [Google Scholar] [CrossRef]
- Hortal, J.; Lobo, J.M.; Jiménez-Valverde, A. Limitations of Biodiversity Databases: Case Study on Seed-Plant Diversity in Tenerife, Canary Islands. Conserv. Biol. 2007, 21, 853–863. [Google Scholar] [CrossRef]
- Hortal, J.; Rodríguez, J.; Nieto-Díaz, M.; Lobo, J.M. Regional and Environmental Effects on the Species Richness of Mammal Assemblages. J. Biogeogr. 2008, 35, 1202–1214. [Google Scholar] [CrossRef]
- Jang, W.; Kim, B.; Chung, O.-S.; Lee, J.K. Analysis of Water Deer Roadkills Using Point Process Modeling in Chungcheongnamdo, South Korea. Forests 2022, 13, 209. [Google Scholar] [CrossRef]
- MOLIT Statistics System Korean Statistical Information Service. Available online: https://kosis.kr/index/index.do (accessed on 22 August 2023).
- Chung, O.-S.; Lee, J.K. Association of Leopard Cat Occurrence with Environmental Factors in Chungnam Province, South Korea. Animals 2023, 13, 122. [Google Scholar] [CrossRef]
- Korea Forest Service. Statistical Yearbook of Forestry 2018; Korea Forest Service: Daejeon, Republic of Korea, 2018. [Google Scholar]
- Kim, E.S.; Choi, H.I. Estimation of the Relative Severity of Floods in Small Ungauged Catchments for Preliminary Observations on Flash Flood Preparedness: A Case Study in Korea. Int. J. Environ. Res. Public Health 2012, 9, 1507–1522. [Google Scholar] [CrossRef]
- Korea Meteorological Administration Korea Meteorological Administration Website. Available online: https://www.weather.go.kr/w/index.do (accessed on 14 May 2023).
- Jang, W.; Seol, A.; Chung, O.-S.; Sagong, J.; Lee, J.K. Avian Reporting Rates in Chugcheongnam Province, South Korea Depend on Distance from Forest Edge, Size of Trees, and Size of Forest Fragments. Forests 2019, 10, 364. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Greenwell, B.; Boehmke, B.; Cunningham, J. GBM Developers. Gbm: Generalized Boosted Regression Models. Available online: https://cran.r-project.org/web/packages/gbm/gbm.pdf (accessed on 13 August 2022).
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. Available online: https://journal.r-project.org/articles/RN-2002-022/RN-2002-022.pdf (accessed on 19 December 2023).
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. Dismo: Species Distribution Modeling. Available online: https://cran.r-project.org/web/packages/dismo/dismo.pdf (accessed on 4 December 2022).
- Therneau, T.; Atkinson, B. Rpart: Recursive Partitioning and Regression Trees. Available online: https://cran.r-project.org/web/packages/rpart/rpart.pdf (accessed on 18 May 2021).
- Meyer, D.; Dimitriadou, E.; Hornik, K.; Weingessel, A.; Leisch, F. E1071: Misc Functions of the Department of Statistics, Probability Theory Group (Formerly: E1071), TU Wien. Available online: https://cran.r-project.org/web/packages/e1071/e1071.pdf (accessed on 28 June 2021).
- Mateo, R.G.; de la Estrella, M.; Felicísimo, Á.M.; Muñoz, J.; Guisan, A. A New Spin on a Compositionalist Predictive Modelling Framework for Conservation Planning: A Tropical Case Study in Ecuador. Biol. Conserv. 2013, 160, 150–161. [Google Scholar] [CrossRef]
- Guisan, A.; Rahbek, C. SESAM–A New Framework Integrating Macroecological and Species Distribution Models for Predicting Spatio-Temporal Patterns of Species Assemblages. J. Biogeogr. 2011, 38, 1433–1444. [Google Scholar] [CrossRef]
- Crisp, M.D.; Laffan, S.; Linder, H.P.; Monro, A. Endemism in the Australian Flora. J. Biogeogr. 2001, 28, 183–198. [Google Scholar] [CrossRef]
- Pottier, J.; Dubuis, A.; Pellissier, L.; Maiorano, L.; Rossier, L.; Randin, C.F.; Vittoz, P.; Guisan, A. The Accuracy of Plant Assemblage Prediction from Species Distribution Models Varies along Environmental Gradients. Glob. Ecol. Biogeogr. 2013, 22, 52–63. [Google Scholar] [CrossRef]
- Rovero, F.; Martin, E.; Rosa, M.; Ahumada, J.A.; Spitale, D. Estimating Species Richness and Modelling Habitat Preferences of Tropical Forest Mammals from Camera Trap Data. PLoS ONE 2014, 9, e103300. [Google Scholar] [CrossRef]
- de la Sancha, N.U. Patterns of Small Mammal Diversity in Fragments of Subtropical Interior Atlantic Forest in Eastern Paraguay. Mammalia 2014, 78, 437–449. [Google Scholar] [CrossRef]
- Laurance, W.F. Rainforest Fragmentation and the Structure of Small Mammal Communities in Tropical Queensland. Biol. Conserv. 1994, 69, 23–32. [Google Scholar] [CrossRef]
- Rickart, E.A. Elevational Diversity Gradients, Biogeography and the Structure of Montane Mammal Communities in the Intermountain Region of North America. Glob. Ecol. Biogeogr. 2001, 10, 77–100. [Google Scholar] [CrossRef]
- Li, J.S.; Song, Y.L.; Zeng, Z.G. Elevational Gradients of Small Mammal Diversity on the Northern Slopes of Mt. Qilian, China. Glob. Ecol. Biogeogr. 2003, 12, 449–460. [Google Scholar] [CrossRef]
- Rahbek, C. The Elevational Gradient of Species Richness: A Uniform Pattern? Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Lomolino, M.V. Elevation Gradients of Species-Density: Historical and Prospective Views. Glob. Ecol. Biogeogr. 2001, 10, 3–13. [Google Scholar] [CrossRef]
- Colwell, R.K.; Lees, D.C. The Mid-Domain Effect: Geometric Constraints on the Geography of Species Richness. Trends Ecol. Evol. 2000, 15, 70–76. [Google Scholar] [CrossRef]
- Shuai, L.-Y.; Ren, C.-L.; Yan, W.-B.; Song, Y.-L.; Zeng, Z.-G. Different Elevational Patterns of Rodent Species Richness between the Southern and Northern Slopes of a Mountain. Sci. Rep. 2017, 7, 8743. [Google Scholar] [CrossRef]
- Jeong, E.; Cho, M.; Cho, H.; Cho, B.; Han, S. Characteristics of Forest Road Cut Slopes Affecting the Movement of Mammals in South Korea. For. Sci. Technol. 2021, 17, 155–161. [Google Scholar] [CrossRef]
- Park, Y.-S.; Lee, W.-S. Characteristics of Habitat-using of Siberian Roe Deer in Seoraksan(Mt.) National Park. J. Korean Soc. Environ. Restor. Technol. 2014, 17, 91–109. [Google Scholar] [CrossRef]
- Jeong, S.; Kim, H.G.; Thorne, J.H.; Lee, H.; Cho, Y.-H.; Lee, D.K.; Park, C.H.; Seo, C. Evaluating Connectivity for Two Mid-Sized Mammals across Modified Riparian Corridors with Wildlife Crossing Monitoring and Species Distribution Modeling. Glob. Ecol. Conserv. 2018, 16, e00485. [Google Scholar] [CrossRef]
- Kim, Y.; Cho, S.; Choung, Y. Habitat Preference of Wild Boar (Sus scrofa) for Feeding in Cool-Temperate Forests. J. Ecol. Environ. 2019, 43, 30. [Google Scholar] [CrossRef]
- Wu, J.; Wang, J.; Zhu, Y.; Bu, X.; Xiang, R.; Lu, Q.; Cui, S.; Hao, Y.; Sheng, Y.; Meng, X. Summer Habitat Selection and Impacts of Human Disturbance on Leopard Cats (Prionailurus bengalensis). Ecosyst. Health Sustain. 2020, 6, 1856630. [Google Scholar] [CrossRef]
- Cincotta, R.P. The Biological Diversity That Is Humanly Possible: Three Models Relevant to Human Population’s Relationship with Native Species. In Human Population: Its Influences on Biological Diversity; Cincotta, R.P., Gorenflo, L.J., Eds.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2011; pp. 61–73. ISBN 978-3-642-16707-2. [Google Scholar]
- McKee, J.; Chambers, E.; Guseman, J. Human Population Density and Growth Validated as Extinction Threats to Mammal and Bird Species. Hum. Ecol. 2013, 41, 773–778. [Google Scholar] [CrossRef]
- Luck, G.W. A Review of the Relationships between Human Population Density and Biodiversity. Biol. Rev. 2007, 82, 607–645. [Google Scholar] [CrossRef]
- Megaze, A.; Balakrishnan, M.; Belay, G. Current Population Estimate and Distribution of the African Buffalo in Chebera Churchura National Park, Ethiopia. Afr. J. Ecol. 2018, 56, 12–19. [Google Scholar] [CrossRef]
- Gunda, D.M.; Chambi, D.; Eustace, A. Do Vegetation, Disturbances, and Water Influence Large Mammal Distribution? Geol. Ecol. Landsc. 2022, 6, 150–158. [Google Scholar] [CrossRef]
- Rich, L.N.; Miller, D.A.W.; Robinson, H.S.; McNutt, J.W.; Kelly, M.J. Using Camera Trapping and Hierarchical Occupancy Modelling to Evaluate the Spatial Ecology of an African Mammal Community. J. Appl. Ecol. 2016, 53, 1225–1235. [Google Scholar] [CrossRef]
- Rho, P.-H. Using a Landscape Ecological Approach to Develop a Wildlife Habitat Model in Korea; Korea Environment Institute: Seoul, Republic of Korea, 2004; p. 22. [Google Scholar]
- de Boer, W.F.; Vis, M.J.P.; de Knegt, H.J.; Rowles, C.; Kohi, E.M.; van Langevelde, F.; Peel, M.; Pretorius, Y.; Skidmore, A.K.; Slotow, R.; et al. Spatial Distribution of Lion Kills Determined by the Water Dependency of Prey Species. J. Mammal. 2010, 91, 1280–1286. [Google Scholar] [CrossRef]
- Barry, S.; Elith, J. Error and Uncertainty in Habitat Models. J. Appl. Ecol. 2006, 43, 413–423. [Google Scholar] [CrossRef]
- Lyons, D.A.; Lowen, J.B.; Therriault, T.W.; Brickman, D.; Guo, L.; Moore, A.M.; Peña, A.; Wang, Z.; DiBacco, C. Identifying Marine Invasion Hotspots Using Stacked Species Distribution Models. Biol. Invasions 2020, 22, 3406–3423. [Google Scholar] [CrossRef]
- Hortal, J.; De Marco, P., Jr.; Santos, A.M.C.; Diniz-Filho, J.A.F. Integrating Biogeographical Processes and Local Community Assembly. J. Biogeogr. 2012, 39, 627–628. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A Misleading Measure of the Performance of Predictive Distribution Models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Leroy, B.; Delsol, R.; Hugueny, B.; Meynard, C.N.; Barhoumi, C.; Barbet-Massin, M.; Bellard, C. Without Quality Presence–Absence Data, Discrimination Metrics Such as TSS Can Be Misleading Measures of Model Performance. J. Biogeogr. 2018, 45, 1994–2002. [Google Scholar] [CrossRef]
- Grenié, M.; Violle, C.; Munoz, F. Is Prediction of Species Richness from Stacked Species Distribution Models Biased by Habitat Saturation? Ecol. Indic. 2020, 111, 105970. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Variables | Notation | Unit | Note | Source † |
|---|---|---|---|---|
| Mammal Occurrence | - | - | Binomial | CI |
| Forest Area | F_AREA | m2 | KFS | |
| Diameter Class | DIA_CL | - | Categorical data | KFS |
| Distance to Forest Edge | DIST_FOR | m | Log-transformed | KFS |
| Distance to Water Channel | DIST_WAT | m | Log-transformed | KNGII |
| Elevation | ELEV | m | KNGII | |
| Slope | SLOPE | % | KNGII | |
| Population Density | POP_DEN | people km−2 | Log-transformed | KNGII |
| Road Density | RD_DEN | roads km−2 | KNGII | |
| Annual Mean Temperature | BIO1 | °C | WorldClim | |
| Mean Diurnal Range | BIO2 | °C | WorldClim | |
| Isothermality | BIO3 | % | BIO2/BIO7 × 100 | WorldClim |
| Temperature Seasonality | BIO4 | °C | Standard deviation × 100 | WorldClim |
| Max Temperature of Warmest Month | BIO5 | °C | WorldClim | |
| Min Temperature of Coldest Month | BIO6 | °C | WorldClim | |
| Temperature Annual Range | BIO7 | °C | BIO5-BIO6 | WorldClim |
| Mean Temperature of Wettest Quarter | BIO8 | °C | WorldClim | |
| Mean Temperature of Driest Quarter | BIO9 | °C | WorldClim | |
| Mean Temperature of Warmest Quarter | BIO10 | °C | WorldClim | |
| Mean Temperature of Coldest Quarter | BIO11 | °C | WorldClim | |
| Annual Precipitation | BIO12 | mm | WorldClim | |
| Precipitation of Wettest Month | BIO13 | mm | WorldClim | |
| Precipitation of Driest Month | BIO14 | mm | WorldClim | |
| Precipitation Seasonality | BIO15 | - | Coefficient of variation | WorldClim |
| Precipitation of Wettest Quarter | BIO16 | mm | WorldClim | |
| Precipitation of Driest Quarter | BIO17 | mm | WorldClim | |
| Precipitation of Warmest Quarter | BIO18 | mm | WorldClim | |
| Precipitation of Coldest Quarter | BIO19 | mm | WorldClim |
| Species Name | Scientific Name | Notation | Occurrences |
|---|---|---|---|
| Korean water deer | Hydropotes inermis | HYIN | 1209 |
| Large mole | Mogera robusta | MORO | 858 |
| Eurasian red squirrel | Sciurus vulgaris | SCVU | 675 |
| Common raccoon dog | Nyctereutes procyonoides | NYPR | 545 |
| Leopard cat | Prionailurus bengalensis | PRBE | 370 |
| Siberian chipmunks | Eutamias sibiricus | EUSI | 177 |
| Yellow weasel | Mustela sibirica | MUSI | 167 |
| Korean hare | Lepus coreanus | LECO | 136 |
| Wild boar | Sus scrofa | SUSC | 90 |
| Eurasian river otter | Lutra lutra | LULU | 76 |
| Asian Badger | Meles leucurus | MELE | 34 |
| Amur hedgehog | Erinaceus amurensis | ERAM | 24 |
| Siberian roe deer | Capreolus pygargus | CAPY | 23 |
| Yellow-throated marten | Martes flavigula | MAFL | 10 |
| Eurasian harvest mouse | Micromys minutus | MIMI | 10 |
| Siberian flying squirrel | Pteromys volans | PTVO | 5 |
| Variable | Relative Importance | Impacted Species |
|---|---|---|
| DIST_FOR | 28.4 | EUSI (16.8), HYIN (74.6), LECO (19.4), LULU (17.5), MORO (78.6), MUSI (43.7), NYPR (51.6), PRBE (54.2), SCVU (62.5), SUSC (7.9) |
| ELEV | 10.1 | CAPY (13.8), ERAM (12.2), EUSI (12.8), HYIN (5.5), MAFL (9.5), MELE (13.3), MUSI (9.3), NYPR (5.4), PRBE (5.6), SUSC (50.5) |
| SLOPE | 5.8 | CAPY (9.7), ERAM (16.0), LECO (12.4), LULU (10.9), MAFL (7.8), MORO (2.9) |
| POP_DEN | 4.9 | MAFL (15.3), MELE (11.6) |
| DIST_WAT | 4.1 | LECO (11.7), MELE (13.4) |
| DIA_CL | 3.7 | CAPY (16.9), LULU (7.5), MORO (1.6) |
| BIO10 | 3.3 | ERAM (22.1) |
| BIO13 | 2.9 | - |
| F_AREA | 2.7 | SUSC (4.6) |
| BIO4 | 2.7 | EUSI (6.6), SCVU (4.3) |
| Species Richness Error | Prediction Success | Cohen’s Kappa | Specificity | Sensitivity | Jaccard Index |
|---|---|---|---|---|---|
| 3.89 | 0.72 | 1.00 | 0.67 | 0.93 | 0.41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, O.-S.; Lee, J.K. Assessing the Distribution and Richness of Mammalian Species Using a Stacking Species Distribution Model in a Temperate Forest. Animals 2024, 14, 759. https://doi.org/10.3390/ani14050759
Chung O-S, Lee JK. Assessing the Distribution and Richness of Mammalian Species Using a Stacking Species Distribution Model in a Temperate Forest. Animals. 2024; 14(5):759. https://doi.org/10.3390/ani14050759
Chicago/Turabian StyleChung, Ok-Sik, and Jong Koo Lee. 2024. "Assessing the Distribution and Richness of Mammalian Species Using a Stacking Species Distribution Model in a Temperate Forest" Animals 14, no. 5: 759. https://doi.org/10.3390/ani14050759
APA StyleChung, O.-S., & Lee, J. K. (2024). Assessing the Distribution and Richness of Mammalian Species Using a Stacking Species Distribution Model in a Temperate Forest. Animals, 14(5), 759. https://doi.org/10.3390/ani14050759