Simple Summary
Bovine breeding is an economically important endeavor in China. Ovarian traits play a significant role in the reproductive characteristics. This study aimed to underscore the discovery of a novel indel locus within the HMGA2 gene in Holstein cows. Our study found that a variant in the bovine HMGA2 gene has a significant connection with ovarian traits. These findings contribute to a more promising outlook for the bovine industry.
Abstract
The high-mobility group AT-hook 2(HMGA2) gene has been widely studied in the context of cancer and animal growth. However, recently, several studies have uncovered its critical role in cell proliferation. A genome-wide association study (GWAS) further suggests that the HMGA2 gene is a candidate gene in fertility, indicating its connection not only to growth traits but also to reproduction, specifically ovarian traits. Thus, this study aimed to analyze the distribution of the HMGA2 gene in 54 bovine breeds worldwide, identify important short fragment variants (indels), and investigate the relationship between HMGA2 and ovarian development. The dataset included genotypic information from a bovine population of 634 individuals (n = 634). After genotyping and analyzing four selected loci, we found that one out of four loci, rs133750033 (P4-D22-bp), was polymorphic. Our results also reveal that this indel of HMGA2 is significantly associated with certain ovarian traits (p < 0.05). Specifically, it has connection with ovarian length (p = 0.004) and ovarian height (p = 0.026) during diestrus. Additionally, we discovered a higher expression of the HMGA2 gene in Asian cattle breeds. In summary, this study suggests that HMGA2 has the potential to serve as an animal fertility testing marker gene. Moreover, these findings contribute to a more promising outlook for the bovine industry.
1. Introduction
Boosting the reproductive capacity of female livestock has been an ongoing economic priority in animal breeding. Specifically in the cattle sector, the persistent selection of high-producing dairy cows has resulted in a decline in female fertility [1]. Consequently, in recent times, there has been a growing focus on research aimed at enhancing bovine fertility. Various metrics, such as the development of ovaries and the caliber of mature ovarian follicles, are employed to assess cattle’s reproductive capabilities. The optimal development of reproductive organs, including ovaries and testes, is associated with the production of superior quality gametes (sperm and ova). This is particularly vital in bovines, where the quality of mature follicles is crucial for the efficacy of artificial insemination techniques. Following the release of follicles by the ovaries, structures akin to ovarian glands, notably the corpus luteum, undergo rapid transformation from these follicles. The corpus luteum functions as a ‘temporary gland’, primarily engaged in the synthesis and release of progesterone. It may also undergo transformation or decay into the white body, playing a critical role in altering or sustaining the receptivity of the endometrium for pregnancy. Thus, identifying key genes influencing the functions of the ovaries, corpus luteum, and related characteristics is fundamental for employing molecular marker-assisted selection techniques to augment cattle fertility [2,3].
Fertility in cattle is a multifaceted characteristic influenced by numerous genes and mutations that are involved in reproductive processes, spanning a range of biological pathways. In the recent era, fueled by the swift advancements in sequencing technologies, there is an emerging trend to elucidate the underlying causes and mechanisms of complex traits in Holstein cows. This is achieved through comprehensive genome sequencing and bioinformatics analyses, encompassing the estimation of genetic substitution rates, exploring associations between milk composition and breeding values, and identifying potential candidate genes linked to longevity, fertility, and other intricate traits [4]. Bovine breeding is also an important economic issue in China. Therefore, the study of how to improve the breeding capacity of bovine remains a top priority. The ovary, a crucial organ in female reproduction, is responsible for producing ova and secreting estrogen. Numerous studies have highlighted that the morphology of the ovary, influenced by various elements such as hormonal levels and genetic factors, directly correlates with a female’s reproductive capabilities [5]. It has been consistently observed that young, non-pregnant cows with larger ovaries exhibit a greater quantity of follicles and oocytes. Thus, the structure and functionality of the ovary play pivotal roles in the regulation of reproductive processes [6].
The high-mobility group AT-hook 2 (HMGA2) gene is well known for its strong connection with animal growth [7]. First of all, it was found that the gene is significantly associated with human height [8]. Studies also indicate a strong association between the HMGA2 gene and cancer. For example, HMGA2 is dysregulated in a wide array of human malignancies, encompassing lung, breast, and ovarian cancers, where its heightened expression is linked to an increased likelihood of cancer advancement [9]. RNA sequencing comparisons between cancerous and normal tissues have revealed diverse expression patterns of HMGA2 across various cancer types [10].
Typically, HMGA2 shows greater expression in sarcomas, brain and central nervous system tumors, as well as in cancers of the esophagus, head and neck, lung, melanoma, ovary, and pancreas, relative to their normal tissue counterparts. Subsequently, researchers observed that the influence of this gene extended to animals, establishing notable associations, such as its association with the body sizes of horses and dogs [10,11,12]. The HMGA2 gene with respect to growth regulation is highly conserved among mammals [13]. Research indicates a potential association between HMGA2 and ovarian traits [14]. Recent investigations in oncology have illuminated the significant involvement of the HMGA2 in key processes of tumor formation, such as the epithelial–mesenchymal transition (EMT) and the development of new blood vessels (angiogenesis). Studies suggest that HMGA2 plays an essential role in controlling angiogenesis in both endothelial cells and their progenitor counterparts. Notably, a study focusing on oral squamous cell carcinoma revealed a noteworthy upregulation in HMGA2 accompanied by a heightened expression in angiogenic genes such as vascular endothelial growth factor (VEGF)-A, VEGF-C, and fibroblast growth factor-2 in these tumors. This upregulation contributes to a marked increase in blood vessel formation, emphasizing the pivotal role of the HMGA2 gene in angiogenesis regulation. This link between HMGA2 overexpression and elevated vascular density may potentially contribute to tumor formation [15]. The overexpression of HMGA2 is observed in a variety of mesenchymal tumors across multiple organ systems, including those in the head and neck, lungs, bones, breast, and female reproductive organs. Notably, this overexpression is a common feature in uterine smooth muscle tumors and other mesenchymal neoplasms of gynecological origin, where specific molecular and genetic alterations result in the increased expression of HMGA2 [16]. Most recently, as a candidate risk gene, HMGA2 was suggested to be related to polycystic ovary syndrome (PCOS) [17]. Along with this, a new GWAS study has uncovered the fact that the HMGA2 gene may also be a candidate gene for bovine fertility [18]. However, whether there is a connection between the HMGA2 gene and bovine fertility traits remains obscure.
Polycystic ovary syndrome (PCOS) represents the most prevalent endocrine disorder affecting the reproductive health of women in their childbearing years. It is characterized by hyperandrogenism and polycystic ovarian changes, often accompanied by insulin resistance (IR) and a series of metabolic abnormalities similar to the clinical manifestations of type 2 diabetes [19]. A genome wide association study (GWAS) revealed that high-mobility group A2 (HMGA2) is a common susceptibility gene for PCOS and type 2 diabetes [17]. So, we speculate that the HMGA2 gene may also be associated with PCOS in bovine. Moreover, HMGA2 is a non-histone chromosomal protein, with no transcriptional activity, and can regulate the transcription of other genes by altering chromosome structure.
In this study, we explored the potential role of HMGA2 in bovine fertility by analyzing data from 634 cattle at various estrus stages. Our goal was to enhance the understanding of HMGA2 and contribute to the ongoing development of cattle reproductive capabilities.
2. Materials and Methods
2.1. Bovine Ovary Collection
We assembled a comprehensive set of 634 ovary specimens from healthy Holstein cows, all within the uniform age range of 5–6 years and raised under consistent environmental conditions. The samples were collected from fixed dairy farms in Shaanxi Province, and the feeding management and nutritional conditions were the same. The dimensions of each ovary sample, including height, width, length, and weight, were accurately measured by the same individual using a consistent methodology. Furthermore, we carefully documented the number of formations such as of the corpus luteum and corpus albicans, as well as mature follicles, and measured their individual diameters. The measurement tools included a sanitized electronic scale for ovary weight evaluation, and either a vernier caliper or a two-sided ruler to measure the height, width, and length of each ovary. Furthermore, we determined the estrus stage indirectly by typing the corpus luteum: a type 1 and type 2 corpus luteum indicated the late stages of estrus; types 3 and 4 suggested the dioestrum; the absence of follicles and corpus luteum on the ovaries indicated pre-estrus; while no corpus luteum and large follicles indicated estrus (corpus luteum type 1: conical; corpus luteum type 2: volcanic crater; corpus luteum type 3: mushroom shaped; corpus luteum type 4: flattened; corpus luteum type 5: no corpus luteum) [20]. For association analysis, we collected these portions of ovarian samples. The experiments of this study were approved by the International Animal Care and Use Committee of the Northwest A&F University (protocol number: NWAFAC1008).
2.2. InDel Loci Selection and Primer Design
The extraction of bovine genomic DNA was performed using the salt-extraction technique as detailed by Aljanabi and Martinez [21]. For an efficient PCR analysis, the DNA isolated from ovarian tissues was diluted to a concentration of 10 ng/µL. The purity of these samples was ascertained using a NanoDrop 1000 (Thermo Scientific, Waltham, MA, USA).
Utilizing the bovine HMGA2 sequence available on NCBI (https://www.ncbi.nlm.nih.gov, accessed on 3 February 2022), we conducted a search in the Ensemble database (https://asia.ensembl.org/index.html, accessed on 4 February 2022) to identify indels longer than 10 bp, which are typically easier to genotype. This search led to the discovery of 2 indel loci in the 5′ UTR region (rs516271779, rs797520182) and another 2 in an intron (rs797609300, rs133750033), all associated with the bovine HMGA2 gene (GeneID:100297175, NC_037332). A primer for each of these loci was designed using the NCBI’s primer design tool (https://www.ncbi.nlm.nih.gov/tools/primer-blast/, accessed on 4 February 2022) and subsequently synthesized by TSINGKE Biological Technology in Beijing, China, resulting in the development and preparation of 4 primers.
2.3. PCR Amplification and Genotyping
In our genotyping study, we employed 4 distinct primers to analyze polymorphisms across 4 indel loci (Table 1). A composite DNA sample, derived from 24 randomly chosen DNA extracts, exhibited polymorphic variations under a touch-down PCR method. The resulting PCR products, separated on 2.5% agarose gels, were individually amplified for further analysis. Following the Polymerase Chain Reaction (PCR) procedure conducted on a sample of 100 individuals, we analyzed the data for each locus and its association with ovarian features to explore the possible connections between locus polymorphisms and ovarian traits. The entire process and reaction setups adhered to the methodology outlined by Akhatayeva [22].

Table 1.
PCR primer sequences for bovine HMGA2 amplification.
2.4. Statistical Analysis
The assessments of the polymorphism information content (PIC) and Hardy–Weinberg equilibrium (HWE) were executed using the GDIcall Online Calculator, adhering to the methodologies proposed by Nei [23].
To elucidate the relationship between genotypes and ovarian characteristics, we utilized a general linear model, defined as Yij = μ + Gi + Eij. Here, Yij represents the specific ovarian trait of the ijth cow, μ is the aggregate mean, Gi denotes the genotype of the ijth cow, and Eij corresponds to the random error component [24].
To examine the link between indel loci and ovarian characteristics, we employed both one-way ANOVA and the independent sample t-test, utilizing SPSS software version 23.0 (IBM Corp., Armonk, NY, USA). Our methodology involved analyzing indels that displayed 3 genotypes (II, ID, and DD), each with over 3 occurrences, through one-way ANOVA. For scenarios where any of these 3 genotypes presented less than 3 instances, we conducted an independent sample t-test. This extensive analysis with SPSS yielded clear findings on the possible association between indel loci and ovarian features. We considered results with a p-value below 0.05 to be statistically significant.
2.5. Worldwide Bovine Breeds’ Sample Information
Our data were collected from the database of BGVD (http://animal.omics.pro/code/index.php/BosVar, accessed on 3 July 2022), which contains 54 bovine breeds and 432 samples. The set of samples contains the following numbers of individuals: 108 West European, 28 Northeast Asian, 9 Middle East, 9 Tibetan, 47 North-Central Chinese, 21 Northwest Chinese, 33 South Chinese, 24 Indo-Pakistan, 83 Central-South European, and 70 African cattle.
This database also provides the signatures of a selection for 8 groups, 6 of which were the “core” cattle groups and were identified as ancestral groups (Indian indicine (IN), Chinese indicine (CN), East Asian taurine (EA), Eurasian taurine (EUA), European taurine (EUR), African taurine (AFR)). In this research, data processing and integration into the BGVD followed a methodology consistent with established protocols. Initially, Illumina short, 250 bp paired-end reads were mapped to the Btau 5.0.1 genome version (GCF_000003205.7) using the Burrows–Wheeler Aligner (BWA) algorithm, achieving an approximate 13× coverage across the bovine genome in various cattle breeds. Redundant reads were eliminated employing Picard tools (accessible at http://broadinstitute.github.io/picard/, accessed on 3 January 2019). Subsequently, the Genome Analysis Toolkit (GATK) facilitated the identification of SNPs and indels, uncovering roughly 60.4 million autosomal SNPs and about 6.8 million autosomal indels. The phasing of these SNPs was conducted using Beagle software (Version: 5.1). SnpEff was employed for the annotation of SNPs and indels. In utilizing PLINK, the minor allele frequency (MAF) was calculated for the entire cattle population, alongside allele frequencies for individual breeds and a defined “core” cattle group. CNVcaller software (Version: 0.1.1) was instrumental in detecting CNVs, resulting in the identification of 76,634 CNV regions (CNVR) across 432 cattle genomes, which were then annotated using Annovar. Considering the prevalent use of 3 bovine genome versions—Btau 5.0.1, UMD3.1.1, and the more recent ARS-UCD1.2 (project accession: NKLS00000000)—the conversion of variation coordinates to the formats of the latter 2 genomes was achieved through the creation of liftOver chain files (Btau5.0.1ToUMD3.1.1.chain.gz and Btau5.0.1ToARS-UCD1.2.chain.gz) and their subsequent application via liftOver [25]. The geographic information and other detailed information for each sample is provided on the homepage (http://animal.omics.pro/code/index.php/BosVar, accessed on 5 January 2019) and the corresponding “Sample Table” page.
Here, we analyzed the distribution of P4- rs133750033 in the HMGA2 gene in 54 bovine breeds and 6 core bovine breeds.
3. Results
3.1. Identification of the Indels in Bovine HMGA2 Gene
In the process of identifying genotypes within four indel loci using a pooled DNA approach, it was determined that two loci, P1-rs516271779 and P4-rs133750033, exhibited polymorphism, each displaying three genotype forms: homozygous insertion (II), homozygous deletion (DD), and heterozygous (ID) types (Figure S1, Table 1 and Table S1). The P4 locus was specifically located in the fourth intron of the HMGA2 gene (Figure 1). The primer diagram of the bovine HMGA2 gene is shown in Figure 2. The deletion sequence of P4, as revealed through direct sequencing, matched the sequence registered in the NCBI database (P4-22bp, CTGGAAGCAGTCCACTAGCATC) (Figure 3). Furthermore, the PCR products of P4, following a separation through 2.5% agarose gel electrophoresis, are displayed in Figure 4.

Figure 1.
Gene structure of bovine HMGA2 gene.

Figure 2.
Primer diagram of bovine HMGA2 gene.

Figure 3.
Sequence diagram of P4-D22bp of bovine HMGA2 gene.

Figure 4.
Electrophoresis pattern of P4-D22bp of bovine HMGA2 gene.
3.2. Estimation of Polymorphism Parameters of Indel (P4-D22-bp) of HMGA2
Utilizing the genetic variations found in the HMGA2-P4 sites, we computed the frequencies of genotypes and alleles, along with other population genetics parameters. Within the HMGA2-P4 locus, the occurrence of the DD genotype (0.48) surpassed that of the ID genotype (0.34) (Table 2).
Table 2.
Estimation of polymorphism parameters of P4-D22-bp of bovine HMGA2 gene.
To gain a more thorough understanding of the genotype distribution in the P1 and P4 regions, we estimated a range of polymorphism parameters. These encompassed the frequencies of genotypes and alleles, the Hardy–Weinberg equilibrium (HWE), and metrics such as homozygosity (Ho), heterozygosity (He), effective number of alleles (Ne), and polymorphism information content (PIC). For P1, the estimates were as follows: Ho: 0.52, He: 0.48, Ne: 1.84, and PIC: 0.35. Correspondingly, in P4, the estimates were Ho: 0.75, He: 0.25, Ne: 1.33, and PIC: 0.22.
3.3. Association Analysis of HMGA2 with Ovarian Traits
To determine the association of P1 and P4 with ovarian characteristics, we conducted an analysis of their relevance using SPSS 23. As previously mentioned, following the completion of PCR procedures in 100 individuals, we analyzed the relationship between each polymorphic locus and ovarian traits. Upon finalizing the PCR process for groups of 100, 200, 300, and 400 individuals, our findings consistently suggested a linkage between P4 and ovarian traits during diestrus. However, no significant association was observed between P1 and ovarian traits.
P4 was correlated with ovarian length (p = 0.004) and ovarian height (p = 0.026) during diestrus. Individuals with the II and ID genotypes possessed ovaries with greater length and height than those with the DD genotype (Table 3).
Table 3.
Relationships between P4-D22-bp polymorphisms of HMGA2 and ovarian morphological traits during diestrus (Mean ± SE).
3.4. Analysis of HMGA2 Gene Distribution in 54 Bovine Breeds and 6 Core Bovine Breeds Worldwide
Based on the BGVD database (http://animal.omics.pro/code/index.php/BosVar, accessed on 3 July 2022), the distribution of the HMGA2 gene in 54 bovine breeds worldwide is illustrated in Figure 5. This gene exhibits a high frequency in the Mishima (frequency = 0.571), Bashan (frequency = 0.400), and Hanwoo (frequency = 0.389) breeds of cattle. Subsequently, we focused on six core bovine breeds from the database, revealing a strong regional specificity. In comparison to Europe (frequency = 0.013), the gene demonstrates higher frequency in East Asian breeds (frequency = 0.347) (Figure 6).
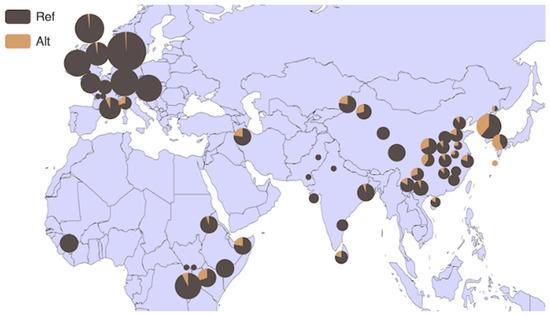
Figure 5.
Allele frequency distribution of P4-D22bp of HMGA2 in 54 cattle breeds worldwide. Note: REF: reference genotype; ALT: alternate genotype.

Figure 6.
Allele frequency distribution of P4-D22bp of HMGA2 in six core cattle breeds worldwide. Note: REF: reference genotype; ALT: alternate genotype.
4. Discussion
Ovaries play a pivotal role in the production of oocytes, a critical component in the reproductive cycle. Additionally, they contain corpus luteum and various components intricately linked to female reproductive endocrine regulation [26,27]. The primary roles of the ovaries involve generating developmentally competent oocytes capable of fertilization, leading to successful conception, and producing sex steroid hormones crucial for initiating and maintaining the estrous cycle as well as supporting pregnancy. The concept of an ovarian reserve refers to the inherent capability of these functions and is recognized as a critical determinant of fertility in mammalian females, including both humans and cattle. This reserve is a key factor in assessing reproductive potential and health in these species [28]. More importantly, the morphological characteristics of the ovary are closely related to the quantity of follicles, exerting an indirect influence on female animal fertility. Enhancing our knowledge of ovarian characteristics is crucial for advancing animal fertility, which is a pivotal factor in increasing the profitability of the animal breeding industry [29]. Scientists have explored the association between ovarian traits and polymorphic mutations. For example, studies have shown that the SEPT7 gene is closely related to ovarian length [30], and other studies have demonstrated a connection between the HSD17B3 gene and ovary morphological traits as well as mature follicle number [31]. The HMGA2 gene is regarded as a candidate gene for improving female fertility, whereas its function still needs further verification.
Up to now, most studies into the association between HMGA2 and animal reproduction have predominantly focused on diseases, including PCOS and ovarian serous papillary carcinoma, as previously mentioned. At the same time, through the database of RGD (http://animal.omics.pro/code/index.php/RGD, accessed on 4 July 2022),we observed that the HMGA2 gene is expressed in the embryonic gonadal ridge, which is related to ovarian traits (Figure S2). The results of our current study strongly suggest a significant relationship between mutations in the HMGA2 gene and ovarian traits, signaling a potential association between this gene and ovarian characteristics. Furthermore, our results align with previous GWAS results that identified the HMGA2 gene as a candidate gene for fertility. Specifically, as previous GWAS studies have shown, microRNA-33b-5p (MIR-33b-5p) is related to the pathogenesis in PCOS where HMGA2 is also involved. MIR-33b-5p can inhibit glucose transporter 4 (GLUT4) production by targeting HMGA2 [32]. In addition, the HMGA2 gene is significantly involved in modulating the GLUT4 expression [32]. The HMGA2 gene is significantly associated with GLUT4, a transporter responsible for glucose absorption in skeletal muscle and adipose tissue in insulin responsive tissues [32]. GLUT4 plays a key role in glucose metabolism in organisms. The connection between HMGA2 and GLUT4, along with their influence on the ovary through microRNA-33b-5p, suggests that HMGA2 may ultimately impact ovarian traits by affecting GLUT4.
Additionally, research has identified HMGA2, a protein typically present during fetal development, as being excessively expressed in ovarian cancer cases [33]. Furthermore, the prevalence of heightened HMGA2 levels is observed in both initial and advanced stages of high-grade ovarian serous papillary carcinoma [34]. Moreover, HMGA2 plays a significant role in regulating cell growth and differentiation, especially during the embryonic stage [35]. This may be another explanation for the close association between the HMGA2 gene and fertility. An analysis of the frequency of the HMGA2 gene in different breeds revealed regional specificity: it is more widely expressed in bovine breeds in Asia compared to European breeds. This phenomenon may be due to the different uses of bovine and the selection for desired traits. Simultaneously, through amino acid sequence alignment, we found that the HMGA2 gene is highly conserved in cattle and humans (Figure S3). Previously, we discussed the possible relationship between the HMGA2 gene and human reproductive PCOS, which suggests that HMGA2 may affect ovarian traits in cattle. To ensure the accuracy and relevance of the analysis, we conducted a phased analysis of ovarian traits according to the estrus cycle, and our results indicate that this mutation site (P4-D22bp) in the HMGA2 gene is related to ovarian traits during diestrus. However, previous studies have shown that HMGA2 may be associated with PCOS, which can lead to the disruption of the estrous cycle. Meanwhile, due to the fact that the trait is controlled by multiple microgenes, our exploration was limited to examining the association between this locus and bovine ovarian traits. The specific nature of their association requires further dedicated study.
Moreover, although the mutation is situated in the fourth intron of the gene, numerous prior studies have highlighted that intronic mutations may be associated with traits. For instance, studies have shown that the introns of the IGF2 gene may affect muscle growth in pigs [36]. Similarly, the mutation in the intron of the DENDD1A gene has been linked to bovine fertility [37], showcasing parallels with our study. In our investigation, this 22 bp short fragment deletion is located in the intron region, lacking a direct influence on protein sequence. Nevertheless, it may still be connected to specific transcription factors, influencing gene expression. In summary, our research establishes a relationship between the newly discovered indel in the HMGA2 gene and various ovarian characteristics in cattle. This indel holds promise for effective utilization in marker-assisted selection (MAS) for bovine breeding purposes. However, to gain a deeper insight into how this indel influences bovine fertility, additional functional studies are required. Fertility constitutes a multifaceted characteristic influenced by a multitude of genes and genetic variations. Through the examination of specific indel loci and their associations with ovarian attributes, our objective is to offer insights into the utilization of molecular marker-assisted selection (MAS) for enhancing bovine female fertility. This advancement holds the potential to expedite progress within the cattle sector.
5. Conclusions
In conclusion, our research underscores the discovery of a novel indel locus within the HMGA2 gene in Holstein cows. Notably, we elucidated its association with ovarian length and ovarian height during diestrus. These findings represent a distinctive contribution to the field of cattle breeding, offering unique insights and emphasizing the diverse applications of genetic methodologies in advancing the industry.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ani14050796/s1, Figure S1: Electrophoresis pattern of bovine HMGA2 gene of P1-I8-bp. Table S1: Polymorphism parameters of P1-I8-bp of bovine HMGA2 gene. Figure S2: Expression of HMGA2 gene in bovine. Note: FPKM: Fragment Per Kilobase of transcript, per Million mapped reads. Figure S3: Phylogenetic tree based on the amino acid sequences of HMGA2 among different species. Note: Bootstrap method: 1000.
Author Contributions
S.S. and L.Z.: draft writing and data analysis; Y.Y., Y.W. and J.L.: data collection; Y.B., C.P. and J.L.: resources and writing—review and editing; X.L. and S.W.: supervision, project administration, and writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key R&D Program of China (2022YFF1000100), the Shaanxi Provincial Innovation Leadership Program in Sciences and Technologies for Young and Middle-aged Scientists (No.2023SR205), the Agricultural Improved Seed Project of Shandong Province under Grant No. 2020LZGC014, and the Project of Undergraduate Science and Technology Innovation Items of Northwest A&F University (X202310712707).
Institutional Review Board Statement
All experimental procedures used in this study followed the principle of the International Animal Care and Use Committee of the Northwest A&F University (protocol number: NWAFAC1008).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We greatly thank Wang’s team for providing access to dairy farms in Shaanxi for us to collect samples for this experiment.
Conflicts of Interest
The authors confirm that they have no conflicts of interest.
References
- Berglund, B. Genetic improvement of dairy cow reproductive performance. Reprod. Domest. Anim. 2008, 43 (Suppl. 2), 89–95. [Google Scholar] [CrossRef]
- Devoto, L.; Henríquez, S.; Kohen, P.; Strauss, J.F. The significance of estradiolmetabolites in human corpus luteum physiology. Steroids 2017, 123, 50–54. [Google Scholar] [CrossRef]
- Lande, R.; Thompson, R. Efficiency of marker-assisted selection in the improvement of quantitative traits. Genetics 1990, 124, 743–756. [Google Scholar] [CrossRef]
- Lin, S.; Zhang, H.; Hou, Y.; Liu, L.; Li, W.; Jiang, J.; Han, B.; Zhang, S.; Sun, D. SNV discovery and functional candidate gene identification for milk composition based on whole genome resequencing of Holstein bulls with extremely high and low breeding values. PLoS ONE 2019, 14, e0220629. [Google Scholar] [CrossRef]
- Gershon, E.; Dekel, N. Newly Identified Regulators of Ovarian Folliculogenesis and Ovulation. Int. J. Mol. Sci. 2020, 21, 4565. [Google Scholar] [CrossRef]
- Bai, Y.; Zhang, F.; Zhang, H.; Xu, C.; Wu, L.; Xia, C. Follicular Fluid Metabolite Changes in Dairy Cows with Inactive Ovary Identified Using Untargeted Metabolomics. BioMed Res. Int. 2020, 2020, 9837543. [Google Scholar] [CrossRef]
- Aguiar, T.S.; Torrecilha, R.B.P.; Milanesi, M.; Utsunomiya, A.T.H.; Trigo, B.B.; Tijjani, A.; Musa, H.H.; Lopes, F.L.; Ajmone-Marsan, P.; Carvalheiro, R.; et al. Association of Copy Number Variation at Intron 3 of HMGA2 With Navel Length in Bos indicus. Front. Genet. 2018, 9, 627. [Google Scholar] [CrossRef] [PubMed]
- Lango Allen, H.; Estrada, K.; Lettre, G.; Berndt, S.I.; Weedon, M.N.; Rivadeneira, F.; Willer, C.J.; Jackson, A.U.; Vedantam, S.; Raychaudhuri, S.; et al. Hundreds of variants clustered in genomic loci and biological pathways affect human height. Nature 2010, 467, 832–838. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; Zhao, J.; Wang, H.; Chen, J.; Wu, J. HMGA2 facilitates colorectal cancer progression via STAT3-mediated tumor-associated macrophage recruitment. Theranostics 2022, 12, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Mansoori, B.; Mohammadi, A.; Ditzel, H.J.; Duijf, P.H.G.; Khaze, V.; Gjerstorff, M.F.; Baradaran, B. HMGA2 as a Critical Regulator in Cancer Development. Genes 2021, 12, 269. [Google Scholar] [CrossRef] [PubMed]
- Boyko, A.R.; Quignon, P.; Li, L.; Schoenebeck, J.J.; Degenhardt, J.D.; Lohmueller, K.E.; Zhao, K.; Brisbin, A.; Parker, H.G.; von Holdt, B.M.; et al. A simple genetic architecture underlies morphological variation in dogs. PLoS Biol. 2010, 8, e1000451. [Google Scholar] [CrossRef]
- Makvandi-Nejad, S.; Hoffman, G.E.; Allen, J.J.; Chu, E.; Gu, E.; Chandler, A.M.; Loredo, A.I.; Bellone, R.R.; Mezey, J.G.; Brooks, S.A.; et al. Four loci explain 83% of size variation in the horse. PLoS ONE 2012, 7, e39929. [Google Scholar] [CrossRef]
- Chung, J.; Zhang, X.; Collins, B.; Sper, R.B.; Gleason, K.; Simpson, S.; Koh, S.; Sommer, J.; Flowers, W.L.; Petters, R.M.; et al. High mobility group A2 (HMGA2) deficiency in pigs leads to dwarfism, abnormal fetal resource allocation, and cryptorchidism. Proc. Natl. Acad. Sci. USA 2018, 115, 5420–5425. [Google Scholar] [CrossRef]
- Liu, M.; Hummitzsch, K.; Hartanti, M.D.; Rosario, R.; Bastian, N.A.; Hatzirodos, N.; Bonner, W.M.; Irving-Rodgers, H.F.; Laven, J.S.E.; Anderson, R.A.; et al. Analysis of expression of candidate genes for polycystic ovary syndrome in adult and fetal human and fetal bovine ovaries. Biol Reprod. 2020, 103, 840–853. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, H. HMGA2 involvement in uterine leiomyomas development through angiogenesis activation. Fertil. Steril. 2020, 114, 974–975. [Google Scholar] [CrossRef]
- Wei, J.J. HMGA2: A Biomarker in Gynecologic Neoplasia. J. Clin. Transl. Pathol. 2022, 2, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhao, H.; Zhao, S.G.; Wei, D.M.; Zhao, Y.R.; Huang, T.; Muhammad, T.; Yan, L.; Gao, F.; Li, L.; et al. The HMGA2-IMP2 Pathway Promotes Granulosa Cell Proliferation in Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2019, 104, 1049–1059. [Google Scholar] [CrossRef]
- Neupane, M.; Geary, T.W.; Kiser, J.N.; Burns, G.W.; Hansen, P.J.; Spencer, T.E.; Neibergs, H.L. Loci and pathways associated with uterine capacity for pregnancy and fertility in beef cattle. PLoS ONE 2017, 12, e0188997. [Google Scholar] [CrossRef]
- Bachelot, A. Polycystic ovarian syndrome: Clinical and biological diagnosis. Ann. Biol. Clin. 2016, 74, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.P.; Redmer, D.A. Growth and development of the corpus luteum. J. Reprod. Fertil. Suppl. 1999, 54, 181–191. [Google Scholar] [CrossRef][Green Version]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Akhatayeva, Z.; Mao, C.; Jiang, F.G.; Pan, C.Y.; Lin, C.J.; Hao, K.J.; Lan, T.X.; Chen, H.; Zhang, Q.F.; Lan, X.Y. Indel variants within the PRL and GHR genes associated with sheep litter size. Reprod. Domest. Anim. 2020, 55, 1470–1478. [Google Scholar] [CrossRef]
- Hui, Y.; Zhang, Y.; Wang, K.; Pan, C.; Chen, H.; Qu, L.; Song, X.; Lan, X. Goat DNMT3B: An indel mutation detection, association analysis with litter size and mRNA expression in gonads. Theriogenology 2020, 147, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, Z.; He, Z.; Tang, W.; Li, T.; Zeng, Z.; He, L.; Shi, Y.Y. A partition-ligation-combination-subdivision EM algorithm for haplotype inference with multiallelic markers: Update of the SHEsis. Cell Res. 2009, 19, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Fu, W.; Zhao, J.; Shen, J.; Chen, Q.; Zheng, Z.; Chen, H.; Sonstegard, T.S.; Lei, C.; Jiang, Y. BGVD: An Integrated Database for Bovine Sequencing Variations and Selective Signatures. Genom. Proteom. Bioinform. 2020, 18, 186–193. [Google Scholar] [CrossRef]
- Cao, C.; Zhou, Q.; Kang, Y.; Zhanerke, A.; Liu, P.; Bai, Y.; Li, R.; Jiang, Y.; Zhang, Q.; Lan, X.; et al. A repertoire of single nucleotide polymorphisms (SNPs) of major fecundity BMPR1B gene among 75 sheep breeds worldwide. Theriogenology 2024, 219, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.K.; Singh, P.; Setia, A.; Sharma, A.K. Insecticides and ovarian functions. Environ. Mol. Mutagen. 2020, 61, 369–392. [Google Scholar] [CrossRef] [PubMed]
- Nagai, K.; Yanagawa, Y.; Katagiri, S.; Nagano, M. The relationship between antral follicle count in a bovine ovary and developmental competence of in vitro-grown oocytes derived from early antral follicles. Biomed. Res. 2016, 37, 63–71. [Google Scholar] [CrossRef]
- Franchi, F.F.; Hernandes, M.P.; Coalho Ferreira, A.L.; Vieira de Lima, V.A.; de Oliveira Mendes, L.; Musa de Aquino, A.; Scarano, W.R.; César de Souza Castilho, A. Fractal analysis and histomolecular phenotyping provides insights into extracellular matrix remodeling in the developing bovine fetal ovary. Biochem. Biophys. Res. Commun. 2020, 523, 823–828. [Google Scholar] [CrossRef]
- Lan, K.; Shen, C.; Li, J.; Zhang, S.; Lan, X.; Pan, C.; Wang, Y. A novel indel within the bovine SEPT7 gene is associated with ovary length. Anim. Biotechnol. 2023, 34, 8–14. [Google Scholar] [CrossRef]
- Li, J.; Zhang, S.; Shen, C.; Niu, Z.; Yang, H.; Zhang, K.; Liu, Z.; Wang, Y.; Lan, X. Indel mutations within the bovine HSD17B3 gene are significantly associated with ovary morphological traits and mature follicle number. J. Steroid Biochem. Mol. Biol. 2021, 209, 105833. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jiang, H.; Xiao, L.; Yang, X. MicroRNA-33b-5p is overexpressed and inhibits GLUT4 by targeting HMGA2 in polycystic ovarian syndrome: An in vivo and in vitro study. Oncol. Rep. 2018, 39, 3073–3085. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, A.; Liu, Z.; Gellert, L.; Zou, X.; Yang, G.; Lee, P.; Yang, X.; Wei, J.J. HMGA2: A biomarker significantly overexpressed in high-grade ovarian serous carcinoma. Mod. Pathol. 2010, 23, 673–681. [Google Scholar] [CrossRef]
- Wu, J.; Liu, Z.; Shao, C.; Gong, Y.; Hernando, E.; Lee, P.; Narita, M.; Muller, W.; Liu, J.; Wei, J.J. HMGA2 overexpression-induced ovarian surface epithelial transformation is mediated through regulation of EMT genes. Cancer Res. 2011, 71, 349–359. [Google Scholar] [CrossRef]
- Hock, R.; Witte, F.; Brocher, J.; Schütz, M.; Scheer, U. Expression of HMGA2 variants during oogenesis and early embryogenesis of Xenopus laevis. Eur. J. Cell Biol. 2006, 85, 519–528. [Google Scholar] [CrossRef]
- Van Laere, A.S.; Nguyen, M.; Braunschweig, M.; Nezer, C.; Collette, C.; Moreau, L.; Archibald, A.L.; Haley, C.S.; Buys, N.; Tally, M.; et al. A regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig. Nature 2003, 425, 832–836. [Google Scholar] [CrossRef]
- Zheng, J.; Deng, T.; Jiang, E.; Li, J.; Wijayanti, D.; Wang, Y.; Ding, X.; Lan, X. Genetic variations of bovine PCOS-related DENND1A gene identified in GWAS significantly affect female reproductive traits. Gene 2021, 802, 145867. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).