
Relationship between Dairy Cow Health and Intensity of Greenhouse Gas Emissions




Abstract
:Simple Summary
Abstract
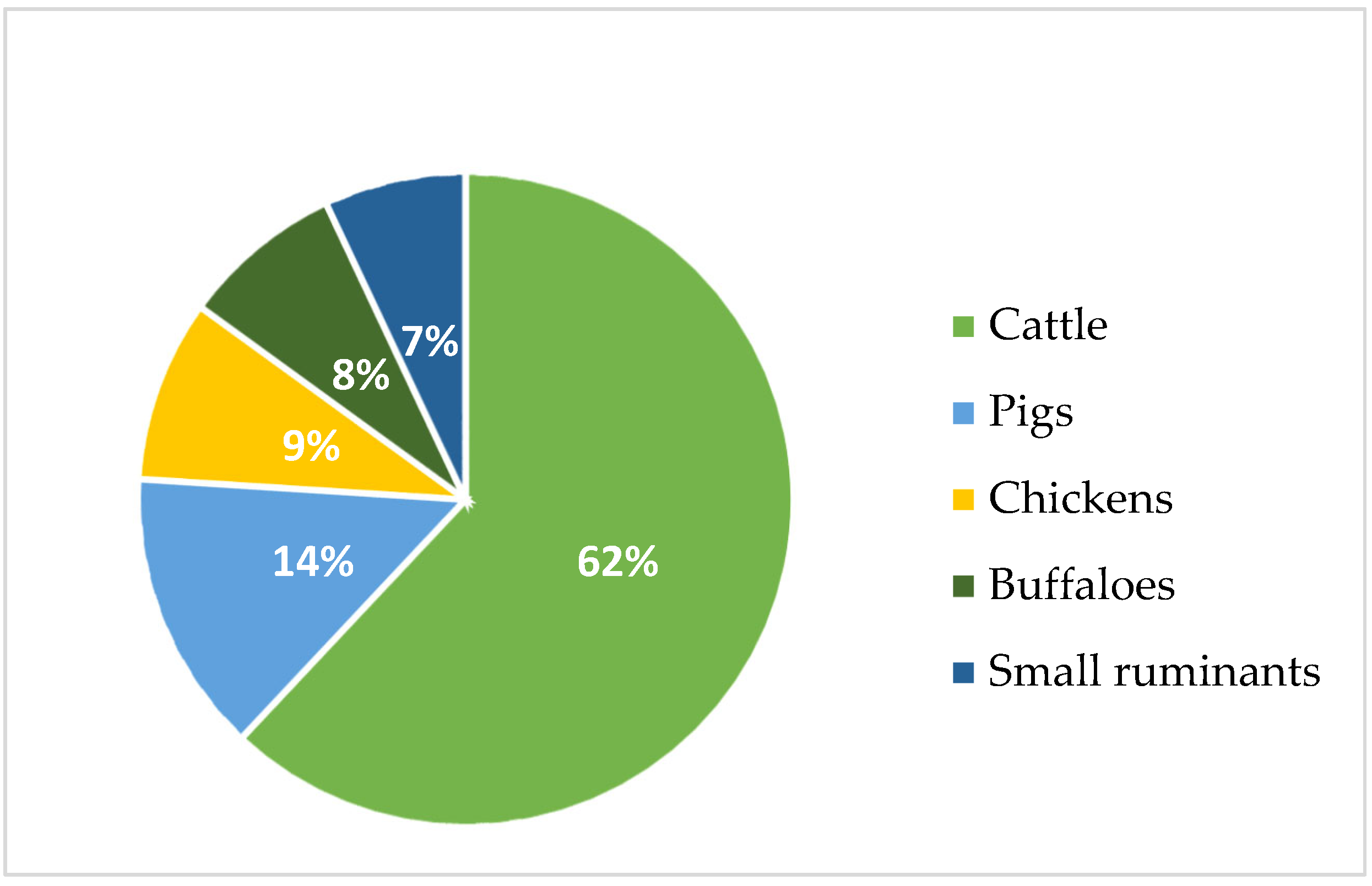
1. Introduction
2. Dairy Cow Health and Methane Emissions
2.1. Metabolic Diseases
2.2. Mastitis
2.3. Lameness
2.4. Parasites
2.5. Viral Infections
2.6. Methane Emissions and Cow Blood Parameters
3. Heritability
4. Methane Emissions and Heat Stress in Dairy Cow
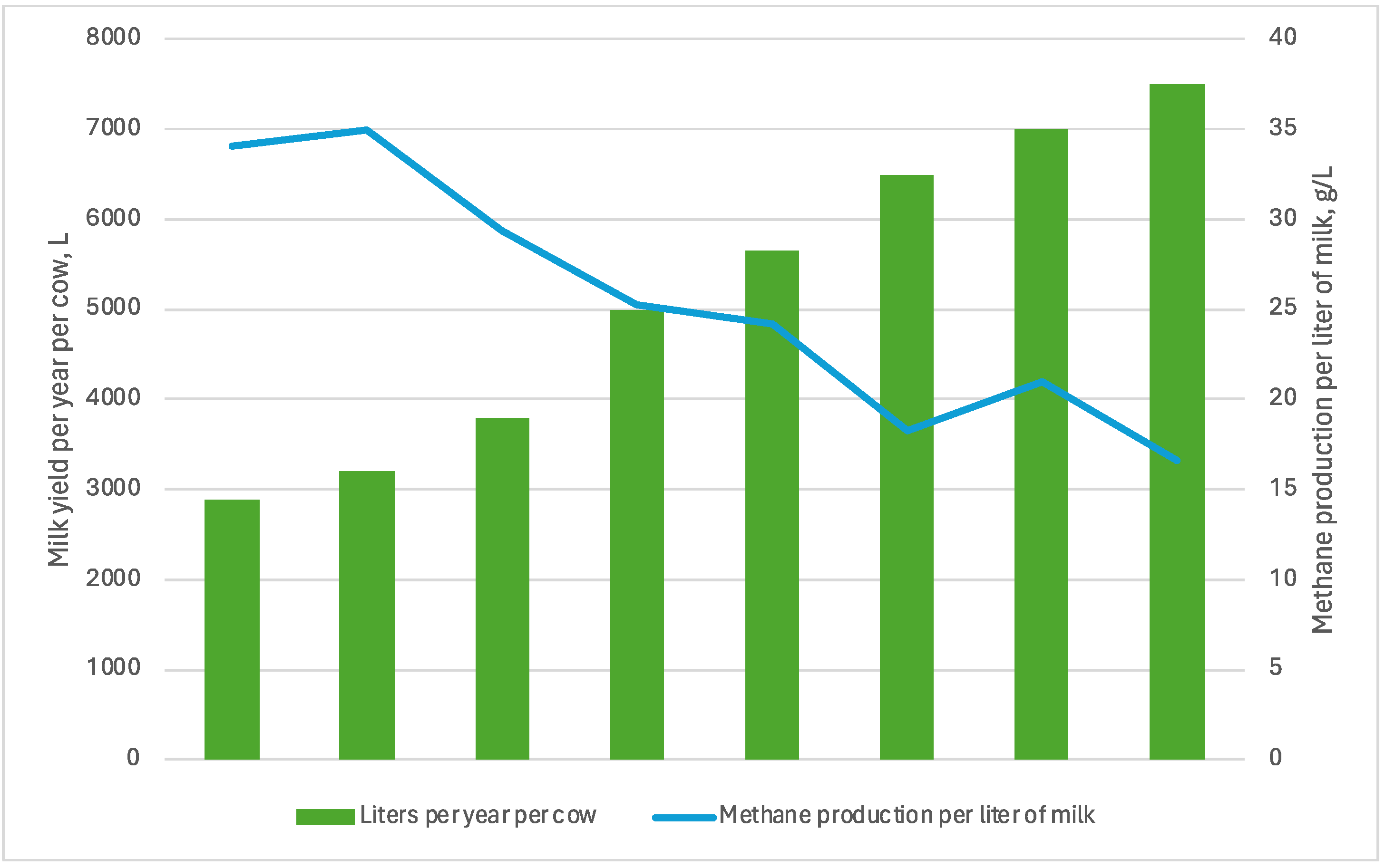
5. Milk Production and Composition
6. Innovative Technologies Registered Physiological Parameters—Smart Dairy
6.1. Body Weight
6.2. Rumination
6.3. Chewing Rates
6.4. Temperament
6.5. Infrared Thermography
7. Breeding Programmes
8. Conclusions
9. Future Implications and Progressive Pathways
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Woolery, S.; Osei, E.; Yu, M.; Guney, S.; Lovell, A.; Jafri, H. The Carbon Footprint of a 5000-Milking-Head Dairy Operation in Central Texas. Agriculture 2023, 13, 2109. [Google Scholar] [CrossRef]
- Blunden, J.; Boyer, T. State of the Climate in 2021. Bull. Am. Meteorol. Soc. 2022, 103, S1–S465. [Google Scholar] [CrossRef]
- Starsmore, K.; Lopez-Villalobos, N.; Shalloo, L.; Egan, M.; Burke, J.; Lahart, B. Animal Factors That Affect Enteric Methane Production Measured Using the GreenFeed Monitoring System in Grazing Dairy Cows. J. Dairy Sci. 2023, S0022-0302(23)00805-6. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.; Crosson, P.; O’Brien, D.; Schulte, R.P.O. The Carbon Navigator: A Decision Support Tool to Reduce Greenhouse Gas Emissions from Livestock Production Systems. Animal 2013, 7, 427–436. [Google Scholar] [CrossRef]
- Reyes, D.C.; Meredith, J.; Puro, L.; Berry, K.; Kersbergen, R.; Soder, K.J.; Quigley, C.; Donihue, M.; Cox, D.; Price, N.N.; et al. Maine Organic Dairy Producers’ Receptiveness to Seaweed Supplementation and Effect of Chondrus Crispus on Enteric Methane Emissions in Lactating Cows. Front. Vet. Sci. 2023, 10, 1153097. [Google Scholar] [CrossRef]
- Britt, J.H.; Cushman, R.A.; Dechow, C.D.; Dobson, H.; Humblot, P.; Hutjens, M.F.; Jones, G.A.; Mitloehner, F.M.; Ruegg, P.L.; Sheldon, I.M.; et al. Review: Perspective on High-Performing Dairy Cows and Herds. Animal 2021, 15, 100298. [Google Scholar] [CrossRef]
- Soliman, T.; Barnes, A.; Helgesen, I.S. The Hidden Carbon Impact of Animal Disease. PLoS ONE 2023, 18, e0292659. [Google Scholar] [CrossRef]
- Herzog, A.; Winckler, C.; Zollitsch, W. In Pursuit of Sustainability in Dairy Farming: A Review of Interdependent Effects of Animal Welfare Improvement and Environmental Impact Mitigation. Agric. Ecosyst. Environ. 2018, 267, 174–187. [Google Scholar] [CrossRef]
- Singaravadivelan, A.; Sachin, P.B.; Harikumar, S.; Vijayakumar, P.; Vindhya, M.V.; Farhana, F.M.B.; Rameesa, K.K.; Mathew, J. Life Cycle Assessment of Greenhouse Gas Emission from the Dairy Production System—Review. Trop. Anim. Health Prod. 2023, 55, 320. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Bovenhuis, H. Validation Strategy Can Result in an Overoptimistic View of the Ability of Milk Infrared Spectra to Predict Methane Emission of Dairy Cattle. J. Dairy Sci. 2019, 102, 6288–6295. [Google Scholar] [CrossRef]
- Aryee, G.; Luecke, S.M.; Dahlen, C.R.; Swanson, K.C.; Amat, S. Holistic View and Novel Perspective on Ruminal and Extra-Gastrointestinal Methanogens in Cattle. Microorganisms 2023, 11, 2746. [Google Scholar] [CrossRef]
- Lassen, J.; Løvendahl, P. Heritability Estimates for Enteric Methane Emissions from Holstein Cattle Measured Using Noninvasive Methods. J. Dairy Sci. 2016, 99, 1959–1967. [Google Scholar] [CrossRef]
- McGuffey, R.K. A 100-Year Review: Metabolic Modifiers in Dairy Cattle Nutrition. J. Dairy Sci. 2017, 100, 10113–10142. [Google Scholar] [CrossRef]
- Bačėninaitė, D.; Džermeikaitė, K.; Antanaitis, R. Global Warming and Dairy Cattle: How to Control and Reduce Methane Emission. Animals 2022, 12, 2687. [Google Scholar] [CrossRef]
- Ghassemi Nejad, J.; Ju, M.-S.; Jo, J.-H.; Oh, K.-H.; Lee, Y.-S.; Lee, S.-D.; Kim, E.-J.; Roh, S.; Lee, H.-G. Advances in Methane Emission Estimation in Livestock: A Review of Data Collection Methods, Model Development and the Role of AI Technologies. Animals 2024, 14, 435. [Google Scholar] [CrossRef]
- New FAO Report Maps Pathways towards Lower Livestock Emissions. Available online: https://www.fao.org/newsroom/detail/new-fao-report-maps-pathways-towards-lower-livestock-emissions/en (accessed on 2 February 2024).
- Lyu, Z.; Shao, N.; Akinyemi, T.; Whitman, W.B. Methanogenesis. Curr. Biol. 2018, 28, R727–R732. [Google Scholar] [CrossRef]
- Conrad, R. Importance of Hydrogenotrophic, Aceticlastic and Methylotrophic Methanogenesis for Methane Production in Terrestrial, Aquatic and Other Anoxic Environments: A Mini Review. Pedosphere 2020, 30, 25–39. [Google Scholar] [CrossRef]
- van Gastelen, S.; Dijkstra, J. Prediction of Methane Emission from Lactating Dairy Cows Using Milk Fatty Acids and Mid-Infrared Spectroscopy. J. Sci. Food Agric. 2016, 96, 3963–3968. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A.; McGinn, S.M. Reducing Methane in Dairy and Beef Cattle Operations: What Is Feasible? Prairie Soil Crop 2008, 1, 17–21. [Google Scholar]
- Min, B.-R.; Lee, S.; Jung, H.; Miller, D.N.; Chen, R. Enteric Methane Emissions and Animal Performance in Dairy and Beef Cattle Production: Strategies, Opportunities, and Impact of Reducing Emissions. Animals 2022, 12, 948. [Google Scholar] [CrossRef] [PubMed]
- de Haas, Y.; Veerkamp, R.F.; de Jong, G.; Aldridge, M.N. Selective Breeding as a Mitigation Tool for Methane Emissions from Dairy Cattle. Animal 2021, 15, 100294. [Google Scholar] [CrossRef]
- Grešáková, Ľ.; Holodová, M.; Szumacher-Strabel, M.; Huang, H.; Ślósarz, P.; Wojtczak, J.; Sowińska, N.; Cieślak, A. Mineral Status and Enteric Methane Production in Dairy Cows during Different Stages of Lactation. BMC Vet. Res. 2021, 17, 287. [Google Scholar] [CrossRef]
- Özkan, Ş.; Vitali, A.; Lacetera, N.; Amon, B.; Bannink, A.; Bartley, D.J.; Blanco-Penedo, I.; De Haas, Y.; Dufrasne, I.; Elliott, J.; et al. Challenges and Priorities for Modelling Livestock Health and Pathogens in the Context of Climate Change. Environ. Res. 2016, 151, 130–144. [Google Scholar] [CrossRef]
- Zetouni, L.; Kargo, M.; Norberg, E.; Lassen, J. Genetic Correlations between Methane Production and Fertility, Health, and Body Type Traits in Danish Holstein Cows. J. Dairy Sci. 2018, 101, 2273–2280. [Google Scholar] [CrossRef]
- Caprarulo, V.; Ventura, V.; Amatucci, A.; Ferronato, G.; Gilioli, G. Innovations for Reducing Methane Emissions in Livestock toward a Sustainable System: Analysis of Feed Additive Patents in Ruminants. Animals 2022, 12, 2760. [Google Scholar] [CrossRef]
- Arnouts, S.; Brown, S.; de Arriba, M.L.; Donabedian, M.; Charlier, J. Technology Readiness Levels for Vaccine and Drug Development in Animal Health: From Discovery to Life Cycle Management. Front. Vet. Sci. 2022, 9, 1016959. [Google Scholar] [CrossRef]
- Dittmann, M.T.; Hammond, K.J.; Kirton, P.; Humphries, D.J.; Crompton, L.A.; Ortmann, S.; Misselbrook, T.H.; Südekum, K.-H.; Schwarm, A.; Kreuzer, M.; et al. Influence of ruminal methane on digesta retention and digestive physiology in non-lactating dairy cattle. Br. J. Nutr. 2016, 116, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Nan, X.; Yang, L.; Zheng, S.; Jiang, L.; Xiong, B. A Review of Enteric Methane Emission Measurement Techniques in Ruminants. Animals 2020, 10, 1004. [Google Scholar] [CrossRef] [PubMed]
- Cameron, L.; Chagunda, M.G.G.; Roberts, D.J.; Lee, M.A. A Comparison of Milk Yields and Methane Production from Three Contrasting High-Yielding Dairy Cattle Feeding Regimes: Cut-and-Carry, Partial Grazing and Total Mixed Ration. Grass Forage Sci. 2018, 73, 789–797. [Google Scholar] [CrossRef]
- Watt, L.J.; Clark, C.E.F.; Krebs, G.L.; Petzel, C.E.; Nielsen, S.; Utsumi, S.A. Differential Rumination, Intake, and Enteric Methane Production of Dairy Cows in a Pasture-Based Automatic Milking System. J. Dairy Sci. 2015, 98, 7248–7263. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Fagodiya, R.K.; Hiloidhari, M.; Dahiya, R.P.; Kumar, A. Methane Production and Estimation from Livestock Husbandry: A Mechanistic Understanding and Emerging Mitigation Options. Sci. Total Environ. 2020, 709, 136135. [Google Scholar] [CrossRef]
- Fu, Y.; Yao, S.; Wang, T.; Lu, Y.; Han, H.; Liu, X.; Lv, D.; Ma, X.; Guan, S.; Yao, Y.; et al. Effects of Melatonin on Rumen Microorganisms and Methane Production in Dairy Cow: Results from in Vitro and in Vivo Studies. Microbiome 2023, 11, 196. [Google Scholar] [CrossRef]
- Nguyen, B.T.; Briggs, K.R.; Eicker, S.; Overton, M.; Nydam, D.V. Herd Turnover Rate Reexamined: A Tool for Improving Profitability, Welfare, and Sustainability. Am. J. Vet. Res. 2023, 84, 1. [Google Scholar] [CrossRef] [PubMed]
- Ezenwa, V.O.; Civitello, D.J.; Classen, A.T.; Barton, B.T.; Becker, D.J.; Brenn-White, M.; Deem, S.L.; Kutz, S.; Malishev, M.; Penczykowski, R.M.; et al. Response to Charlier et al.: Climate–Disease Feedbacks Mediated by Livestock Methane Emissions Are Plausible. Trends Ecol. Evol. 2021, 36, 578–579. [Google Scholar] [CrossRef] [PubMed]
- Hristov, A.N.; Ott, T.; Tricarico, J.; Rotz, A.; Waghorn, G.; Adesogan, A.; Dijkstra, J.; Montes, F.; Oh, J.; Kebreab, E.; et al. SPECIAL TOPICS—Mitigation of Methane and Nitrous Oxide Emissions from Animal Operations: III. A Review of Animal Management Mitigation Options1. J. Anim. Sci. 2013, 91, 5095–5113. [Google Scholar] [CrossRef] [PubMed]
- von Soosten, D.; Meyer, U.; Flachowsky, G.; Dänicke, S. Dairy Cow Health and Greenhouse Gas Emission Intensity. Dairy 2020, 1, 20–29. [Google Scholar] [CrossRef]
- Franzluebbers, A.J. Chapter 2—Cattle Grazing Effects on the Environment: Greenhouse Gas Emissions and Carbon Footprint. In Management Strategies for Sustainable Cattle Production in Southern Pastures; Rouquette, M., Aiken, G.E., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 11–34. ISBN 978-0-12-814474-9. [Google Scholar]
- Lehmann, J.O.; Mogensen, L.; Kristensen, T. Extended Lactations May Improve Cow Health, Productivity and Reduce Greenhouse Gas Emissions from Organic Dairy Production. Org. Agric. 2014, 4, 295–299. [Google Scholar] [CrossRef]
- Kabir, M.; Hasan, M.M.; Tanni, N.S.; Parvin, M.S.; Asaduzzaman, M.; Ehsan, M.A.; Islam, M.T. Metabolic Profiling in Periparturient Dairy Cows and Its Relation with Metabolic Diseases. BMC Res. Notes 2022, 15, 231. [Google Scholar] [CrossRef] [PubMed]
- Lacasse, P.; Vanacker, N.; Ollier, S.; Ster, C. Innovative Dairy Cow Management to Improve Resistance to Metabolic and Infectious Diseases during the Transition Period. Res. Vet. Sci. 2018, 116, 40–46. [Google Scholar] [CrossRef]
- Kappes, A.; Tozooneyi, T.; Shakil, G.; Railey, A.F.; McIntyre, K.M.; Mayberry, D.E.; Rushton, J.; Pendell, D.L.; Marsh, T.L. Livestock Health and Disease Economics: A Scoping Review of Selected Literature. Front. Vet. Sci. 2023, 10, 1168649. [Google Scholar] [CrossRef]
- Capper, J.L.; Williams, P. Investing in Health to Improve the Sustainability of Cattle Production in the United Kingdom: A Narrative Review. Vet. J. 2023, 296–297, 105988. [Google Scholar] [CrossRef] [PubMed]
- Preview: Economic Effects of Bovine Respiratory Disease. J. Anim. Sci. 2020, 98, skaa042. [CrossRef] [PubMed]
- van Soest, F.J.S.; Santman-Berends, I.M.G.A.; Lam, T.J.G.M.; Hogeveen, H. Failure and Preventive Costs of Mastitis on Dutch Dairy Farms. J. Dairy Sci. 2016, 99, 8365–8374. [Google Scholar] [CrossRef] [PubMed]
- Bonestroo, J.; Fall, N.; Hogeveen, H.; Emanuelson, U.; Klaas, I.C.; van der Voort, M. The Costs of Chronic Mastitis: A Simulation Study of an Automatic Milking System Farm. Prev. Vet. Med. 2023, 210, 105799. [Google Scholar] [CrossRef]
- Dolecheck, K.; Bewley, J. Animal board invited review: Dairy cow lameness expenditures, losses and total cost. Animal 2018, 12, 1462–1474. [Google Scholar] [CrossRef] [PubMed]
- Robcis, R.; Ferchiou, A.; Berrada, M.; Ndiaye, Y.; Herman, N.; Lhermie, G.; Raboisson, D. Cost of Lameness in Dairy Herds: An Integrated Bioeconomic Modeling Approach. J. Dairy Sci. 2023, 106, 2519–2534. [Google Scholar] [CrossRef] [PubMed]
- Ettema, J.F.; Krogh, M.A.; Østergaard, S. Economic Value of Information from an Alert System on Physiological Imbalance in Fresh Cows. Prev. Vet. Med. 2020, 181, 105039. [Google Scholar] [CrossRef]
- Cascone, G.; Licitra, F.; Stamilla, A.; Amore, S.; Dipasquale, M.; Salonia, R.; Antoci, F.; Zecconi, A. Subclinical Ketosis in Dairy Herds: Impact of Early Diagnosis and Treatment. Front. Vet. Sci. 2022, 9, 895468. [Google Scholar] [CrossRef]
- Mostert, P.F.; van Middelaar, C.E.; Bokkers, E.A.M.; de Boer, I.J.M. The Impact of Subclinical Ketosis in Dairy Cows on Greenhouse Gas Emissions of Milk Production. J. Clean. Prod. 2018, 171, 773–782. [Google Scholar] [CrossRef]
- Mostert, P.F.; Bokkers, E.A.M.; van Middelaar, C.E.; Hogeveen, H.; de Boer, I.J.M. Estimating the Economic Impact of Subclinical Ketosis in Dairy Cattle Using a Dynamic Stochastic Simulation Model. Animal 2018, 12, 145–154. [Google Scholar] [CrossRef]
- Steeneveld, W.; Amuta, P.; van Soest, F.J.S.; Jorritsma, R.; Hogeveen, H. Estimating the Combined Costs of Clinical and Subclinical Ketosis in Dairy Cows. PLoS ONE 2020, 15, e0230448. [Google Scholar] [CrossRef]
- Antanaitis, R.; Žilaitis, V.; Kucinskas, A.; Juozaitienė, V.; Leonauskaite, K. Changes in Cow Activity, Milk Yield, and Milk Conductivity before Clinical Diagnosis of Ketosis, and Acidosis. Vet. Med. Zootech. 2015, 70, 3–9. [Google Scholar]
- Abdela, N. Sub-Acute Ruminal Acidosis (SARA) and Its Consequence in Dairy Cattle: A Review of Past and Recent Research at Global Prospective. Achiev. Life Sci. 2016, 10, 187–196. [Google Scholar] [CrossRef]
- Paiano, R.B.; Birgel, D.B.; Bonilla, J.; Birgel Junior, E.H. Evaluation of Biochemical Profile of Dairy Cows with Metabolic Diseases in Tropical Conditions. Reprod. Domest. Anim. 2020, 55, 1219–1228. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Masaki, T.; Ikuta, K.; Iwamoto, E.; Nishihara, K.; Hirai, M.; Uemoto, Y.; Terada, F.; Roh, S. Physiological Responses and Adaptations to High Methane Production in Japanese Black Cattle. Sci. Rep. 2022, 12, 11154. [Google Scholar] [CrossRef] [PubMed]
- McFadden, J.W. Review: Lipid Biology in the Periparturient Dairy Cow: Contemporary Perspectives. Animal 2020, 14, s165–s175. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.A.C.; Simões, J. Invited Review: Ketosis Diagnosis and Monitoring in High-Producing Dairy Cows. Dairy 2021, 2, 303–325. [Google Scholar] [CrossRef]
- Simanungkalit, G.; Bhuiyan, M.; Bell, R.; Sweeting, A.; Morton, C.L.; Cowley, F.; Hegarty, R. The Effects of Antibiotic-Free Supplementation on the Ruminal pH Variability and Methane Emissions of Beef Cattle under the Challenge of Subacute Ruminal Acidosis (SARA). Res. Vet. Sci. 2023, 160, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Kaur, U.; Malacco, V.M.R.; Bai, H.; Price, T.P.; Datta, A.; Xin, L.; Sen, S.; Nawrocki, R.A.; Chiu, G.; Sundaram, S.; et al. Invited Review: Integration of Technologies and Systems for Precision Animal Agriculture—A Case Study on Precision Dairy Farming. J. Anim. Sci. 2023, 101, skad206. [Google Scholar] [CrossRef]
- Antanaitis, R.; Anskienė, L.; Rapaliutė, E.; Bilskis, R.; Džermeikaitė, K.; Bačėninaitė, D.; Juškienė, V.; Juška, R.; Meškinytė, E. Relationship between Reticulorumen Parameters Measured in Real Time and Methane Emission and Heat Stress Risk in Dairy Cows. Animals 2022, 12, 3257. [Google Scholar] [CrossRef]
- Mickdam, E.; Khiaosa-ard, R.; Metzler-Zebeli, B.U.; Klevenhusen, F.; Chizzola, R.; Zebeli, Q. Rumen Microbial Abundance and Fermentation Profile during Severe Subacute Ruminal Acidosis and Its Modulation by Plant Derived Alkaloids In Vitro. Anaerobe 2016, 39, 4–13. [Google Scholar] [CrossRef]
- Wang, K.; Xiong, B.; Zhao, X. Could Propionate Formation Be Used to Reduce Enteric Methane Emission in Ruminants? Sci. Total Environ. 2023, 855, 158867. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A.; Ungerfeld, E.M.; Abdalla, A.L.; Alvarez, C.; Arndt, C.; Becquet, P.; Benchaar, C.; Berndt, A.; Mauricio, R.M.; McAllister, T.A.; et al. Invited Review: Current Enteric Methane Mitigation Options. J. Dairy Sci. 2022, 105, 9297–9326. [Google Scholar] [CrossRef] [PubMed]
- Børsting, C.F.; Brask, M.; Hellwing, A.L.F.; Weisbjerg, M.R.; Lund, P. Enteric Methane Emission and Digestion in Dairy Cows Fed Wheat or Molasses. J. Dairy Sci. 2020, 103, 1448–1462. [Google Scholar] [CrossRef] [PubMed]
- Eger, M.; Riede, S.; Breves, G. Induction of a Transient Acidosis in the Rumen Simulation Technique. J. Anim. Physiol. Anim. Nutr. 2018, 102, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Elmhadi, M.E.; Ali, D.K.; Khogali, M.K.; Wang, H. Subacute Ruminal Acidosis in Dairy Herds: Microbiological and Nutritional Causes, Consequences, and Prevention Strategies. Anim. Nutr. 2022, 10, 148–155. [Google Scholar] [CrossRef]
- Kaseke, T.B.; Chikwambi, Z.; Gomo, C.; Mashingaidze, A.B.; Murungweni, C. Antibacterial Activity of Medicinal Plants on the Management of Mastitis in Dairy Cows: A Systematic Review. Vet. Med. Sci. 2023, 9, 2800–2819. [Google Scholar] [CrossRef] [PubMed]
- Mostert, P.F.; Bokkers, E.A.; De Boer, I.J.; Van Middelaar, C.E. Estimating the impact of clinical mastitis in dairy cows on greenhouse gas emissions using a dynamic stochastic simulation model: A case study. Animal 2019, 13, 2913–2921. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Dong, Q.; Feng, Y. Risk Prediction Model of Clinical Mastitis in Lactating Dairy Cows Based on Machine Learning Algorithms. Prev. Vet. Med. 2023, 221, 106059. [Google Scholar] [CrossRef]
- Özkan Gülzari, Ş.; Vosough Ahmadi, B.; Stott, A.W. Impact of Subclinical Mastitis on Greenhouse Gas Emissions Intensity and Profitability of Dairy Cows in Norway. Prev. Vet. Med. 2018, 150, 19–29. [Google Scholar] [CrossRef]
- Ezenwa, V.O.; Civitello, D.J.; Barton, B.T.; Becker, D.J.; Brenn-White, M.; Classen, A.T.; Deem, S.L.; Johnson, Z.E.; Kutz, S.; Malishev, M.; et al. Infectious Diseases, Livestock, and Climate: A Vicious Cycle? Trends Ecol. Evol. 2020, 35, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Potter, T.L.; Arndt, C.; Hristov, A.N. Short Communication: Increased Somatic Cell Count Is Associated with Milk Loss and Reduced Feed Efficiency in Lactating Dairy Cows. J. Dairy Sci. 2018, 101, 9510–9515. [Google Scholar] [CrossRef]
- Hockstad, L.; Hanel, L. Inventory of U.S. Greenhouse Gas Emissions and Sinks; Environmental System Science Data Infrastructure for a Virtual Ecosystem (ESS-DIVE): Washington, DC, USA, 2018. [Google Scholar]
- MacFadden, D.R.; McGough, S.F.; Fisman, D.; Santillana, M.; Brownstein, J.S. Antibiotic Resistance Increases with Local Temperature. Nat. Clim. Chang. 2018, 8, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Weigele, H.C.; Gygax, L.; Steiner, A.; Wechsler, B.; Burla, J.-B. Moderate Lameness Leads to Marked Behavioral Changes in Dairy Cows. J. Dairy Sci. 2018, 101, 2370–2382. [Google Scholar] [CrossRef] [PubMed]
- Herzog, A.; Hörtenhuber, S.; Winckler, C.; Kral, I.; Zollitsch, W. Welfare Intervention and Environmental Impacts of Milk Production—Cradle-to-Farm-Gate Effects of Implementing Rubber Mats in Austrian Dairy Farms. J. Clean. Prod. 2020, 277, 123953. [Google Scholar] [CrossRef]
- Chen, W.; White, E.; Holden, N.M. The Effect of Lameness on the Environmental Performance of Milk Production by Rotational Grazing. J. Environ. Manag. 2016, 172, 143–150. [Google Scholar] [CrossRef]
- Somers, J.; O’Grady, L. Foot Lesions in Lame Cows on 10 Dairy Farms in Ireland. Ir. Vet. J. 2015, 68, 10. [Google Scholar] [CrossRef]
- Mostert, P.F.; van Middelaar, C.E.; de Boer, I.J.M.; Bokkers, E.A.M. The Impact of Foot Lesions in Dairy Cows on Greenhouse Gas Emissions of Milk Production. Agric. Syst. 2018, 167, 206–212. [Google Scholar] [CrossRef]
- Podpečan, O.; Hajdinjak, M.; Posedi, J. Helminth Control as a Part of Animal Welfare Measure Protocol in Grazing Cattle in Slovenia. Agriculture 2023, 13, 1038. [Google Scholar] [CrossRef]
- Fox, N.J.; Smith, L.A.; Houdijk, J.G.M.; Athanasiadou, S.; Hutchings, M.R. Ubiquitous Parasites Drive a 33% Increase in Methane Yield from Livestock. Int. J. Parasitol. 2018, 48, 1017–1021. [Google Scholar] [CrossRef]
- Fernandes, M.A.; de Mello Tavares Lima, P.; do Amarante, A.F.T.; Abdalla, A.L.; Louvandini, H. Hematological, Biochemical Alterations and Methane Production in Sheep Submitted to Mixed Infection of Haemonchus Contortus and Trichostrongylus Colubriformis. Small Rumin. Res. 2022, 216, 106798. [Google Scholar] [CrossRef]
- Houdijk, J.G.M.; Tolkamp, B.J.; Rooke, J.A.; Hutchings, M.R. Animal Health and Greenhouse Gas Intensity: The Paradox of Periparturient Parasitism. Int. J. Parasitol. 2017, 47, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, N.N.; MacLeod, M.; Hayward, A.; McNeilly, T.; Ferguson, K.D.; Skuce, P.J. Liver Fluke in Beef Cattle—Impact on Production Efficiency and Associated Greenhouse Gas Emissions Estimated Using Causal Inference Methods. Prev. Vet. Med. 2022, 200, 105579. [Google Scholar] [CrossRef] [PubMed]
- Wathes, D.C.; Oguejiofor, C.F.; Thomas, C.; Cheng, Z. Importance of Viral Disease in Dairy Cow Fertility. Engineering 2020, 6, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Brito, B.; Hick, P. Milk as a Diagnostic Fluid to Monitor Viral Diseases in Dairy Cattle. Aust. Vet. J. 2024, 102, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Capper, J.L. The Impact of Controlling Diseases of Significant Global Importance on Greenhouse Gas Emissions from Livestock Production. One Health Outlook 2023, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.; Chatterton, J.; Hateley, G.; Curwen, A.; Elliott, J. A Systems-Life Cycle Assessment Approach to Modelling the Impact of Improvements in Cattle Health on Greenhouse Gas Emissions. Adv. Anim. Biosci. 2015, 6, 29–31. [Google Scholar] [CrossRef]
- McAloon, C.G.; Whyte, P.; More, S.J.; Green, M.J.; O’Grady, L.; Garcia, A.; Doherty, M.L. The Effect of Paratuberculosis on Milk Yield—A Systematic Review and Meta-Analysis. J. Dairy Sci. 2016, 99, 1449–1460. [Google Scholar] [CrossRef]
- Reintke, J.; Brügemann, K.; Yin, T.; Wagner, H.; Wehrend, A.; Müller, A.; König, S. Associations between Minerals and Metabolic Indicators in Maternal Blood Pre- and Postpartum with Ewe Body Condition, Methane Emissions, and Lamb Body Weight Development. Anim. Int. J. Anim. Biosci. 2021, 15, 100034. [Google Scholar] [CrossRef]
- Gul, F.; Amin, H.; Naz, S.; Khan, M.T.; Alhidary, I.A.; Khan, R.U.; Pugliese, G.; Tufarelli, V. Evaluation of Blood Minerals and Oxidative Stress Changing Pattern in Prepartum and Postpartum Achai and Holstein Friesian Dairy Cows. Reprod. Domest. Anim. 2024, 59, e14525. [Google Scholar] [CrossRef]
- Ornelas, L.T.C.; Silva, D.C.; Tomich, T.R.; Campos, M.M.; Machado, F.S.; Ferreira, A.L.; Maurício, R.M.; Pereira, L.G.R. Differences in Methane Production, Yield and Intensity and Its Effects on Metabolism of Dairy Heifers. Sci. Total Environ. 2019, 689, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Kamalanathan, S.; Houlahan, K.; Miglior, F.; Chud, T.C.S.; Seymour, D.J.; Hailemariam, D.; Plastow, G.; de Oliveira, H.R.; Baes, C.F.; Schenkel, F.S. Genetic Analysis of Methane Emission Traits in Holstein Dairy Cattle. Animals 2023, 13, 1308. [Google Scholar] [CrossRef] [PubMed]
- Manzanilla-Pech, C.I.V.; Løvendahl, P.; Gordo, D.M.; Difford, G.F.; Pryce, J.E.; Schenkel, F.; Wegmann, S.; Miglior, F.; Chud, T.C.; Moate, P.J.; et al. Breeding for Reduced Methane Emission and Feed-Efficient Holstein Cows: An International Response. J. Dairy Sci. 2021, 104, 8983–9001. [Google Scholar] [CrossRef] [PubMed]
- Stepanchenko, N.; Stefenoni, H.; Hennessy, M.; Nagaraju, I.; Wasson, D.E.; Cueva, S.F.; Räisänen, S.E.; Dechow, C.D.; Pitta, D.W.; Hristov, A.N. Microbial Composition, Rumen Fermentation Parameters, Enteric Methane Emissions, and Lactational Performance of Phenotypically High and Low Methane-Emitting Dairy Cows. J. Dairy Sci. 2023, 106, 6146–6170. [Google Scholar] [CrossRef] [PubMed]
- Pszczola, M.; Rzewuska, K.; Mucha, S.; Strabel, T. Heritability of Methane Emissions from Dairy Cows over a Lactation Measured on Commercial Farms1. J. Anim. Sci. 2017, 95, 4813–4819. [Google Scholar] [CrossRef] [PubMed]
- Neethirajan, S. Innovative Strategies for Sustainable Dairy Farming in Canada amidst Climate Change. Sustainability 2024, 16, 265. [Google Scholar] [CrossRef]
- Pulina, G.; Tondo, A.; Danieli, P.P.; Primi, R.; Matteo Crovetto, G.; Fantini, A.; Macciotta, N.P.P.; Atzori, A.S. How to Manage Cows Yielding 20,000 Kg of Milk: Technical Challenges and Environmental Implications. Ital. J. Anim. Sci. 2020, 19, 865–879. [Google Scholar] [CrossRef]
- Gauly, M.; Ammer, S. Review: Challenges for Dairy Cow Production Systems Arising from Climate Changes. Animal 2020, 14, s196–s203. [Google Scholar] [CrossRef] [PubMed]
- Wankar, A.K.; Singh, G.; Yadav, B. Effect of Temperature x THI on Acclimatization in Buffaloes Subjected to Simulated Heat Stress: Physio-Metabolic Profile, Methane Emission and Nutrient Digestibility. Biol. Rhythm. Res. 2021, 52, 1589–1603. [Google Scholar] [CrossRef]
- Son, A.-R.; Islam, M.; Kim, S.-H.; Lee, S.-S.; Lee, S.-S. Influence of Dietary Organic Trace Minerals on Enteric Methane Emissions and Rumen Microbiota of Heat-Stressed Dairy Steers. J. Anim. Sci. Technol. 2023, 65, 132–148. [Google Scholar] [CrossRef]
- Lanzoni, L.; Chagunda, M.G.G.; Fusaro, I.; Chincarini, M.; Giammarco, M.; Atzori, A.S.; Podaliri, M.; Vignola, G. Assessment of Seasonal Variation in Methane Emissions of Mediterranean Buffaloes Using a Laser Methane Detector. Animals 2022, 12, 3487. [Google Scholar] [CrossRef] [PubMed]
- Souza, V.C.; Moraes, L.E.; Baumgard, L.H.; Santos, J.E.P.; Mueller, N.D.; Rhoads, R.P.; Kebreab, E. Modeling the Effects of Heat Stress in Animal Performance and Enteric Methane Emissions in Lactating Dairy Cows. J. Dairy Sci. 2023, 106, 4725–4737. [Google Scholar] [CrossRef] [PubMed]
- Meneses, J.A.M.; de Sá, O.A.A.L.; Coelho, C.F.; Pereira, R.N.; Batista, E.D.; Ladeira, M.M.; Casagrande, D.R.; Gionbelli, M.P. Effect of Heat Stress on Ingestive, Digestive, Ruminal and Physiological Parameters of Nellore Cattle Feeding Low- or High-Energy Diets. Livest. Sci. 2021, 252, 104676. [Google Scholar] [CrossRef]
- Zehetmeier, M.; Baudracco, J.; Hoffmann, H.; Heißenhuber, A. Does Increasing Milk Yield per Cow Reduce Greenhouse Gas Emissions? A System Approach. Animal 2012, 6, 154–166. [Google Scholar] [CrossRef]
- Lean, I.; Westwood, C.; Playford, M. Livestock Disease Threats Associated with Intensification of Pastoral Dairy Farming. N. Z. Vet. J. 2008, 56, 261–269. [Google Scholar] [CrossRef]
- Moate, P.; Williams, S.R.O.; Deighton, M.; Pryce, J.; Hayes, B.; Jacobs, J.; Eckard, R.; Hannah, M.; Wales, B. Mitigation of Enteric Methane Emissions from the Australian Dairy Industry. In Proceedings of the 5th Australasian Dairy Science Symposium, Hamilton, New Zealand, 19–21 November 2014. [Google Scholar]
- Breider, I.S.; Wall, E.; Garnsworthy, P.C. Short Communication: Heritability of Methane Production and Genetic Correlations with Milk Yield and Body Weight in Holstein-Friesian Dairy Cows. J. Dairy Sci. 2019, 102, 7277–7281. [Google Scholar] [CrossRef]
- Bell, M.J.; Potterton, S.L.; Craigon, J.; Saunders, N.; Wilcox, R.H.; Hunter, M.; Goodman, J.R.; Garnsworthy, P.C. Variation in Enteric Methane Emissions among Cows on Commercial Dairy Farms. Animal 2014, 8, 1540–1546. [Google Scholar] [CrossRef] [PubMed]
- Hardan, A.; Garnsworthy, P.C.; Bell, M.J. Variability in Enteric Methane Emissions among Dairy Cows during Lactation. Animals 2023, 13, 157. [Google Scholar] [CrossRef] [PubMed]
- Yanibada, B.; Hohenester, U.; Pétéra, M.; Canlet, C.; Durand, S.; Jourdan, F.; Ferlay, A.; Morgavi, D.P.; Boudra, H. Milk Metabolome Reveals Variations on Enteric Methane Emissions from Dairy Cows Fed a Specific Inhibitor of the Methanogenesis Pathway. J. Dairy Sci. 2021, 104, 12553–12566. [Google Scholar] [CrossRef]
- Negussie, E.; de Haas, Y.; Dehareng, F.; Dewhurst, R.J.; Dijkstra, J.; Gengler, N.; Morgavi, D.P.; Soyeurt, H.; van Gastelen, S.; Yan, T.; et al. Invited Review: Large-Scale Indirect Measurements for Enteric Methane Emissions in Dairy Cattle: A Review of Proxies and Their Potential for Use in Management and Breeding Decisions. J. Dairy Sci. 2017, 100, 2433–2453. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.R.O.; Marett, L.; Moate, P.; Garner, J.; Pryce, J.; Hayes, B.; Wales, B. Selecting Cows for Low Enteric Methane Emissions May Affect Milk Composition. In Proceedings of the Australasian Dairy Science Symposium, Hamilton, New Zealand, 19–21 November 2014. [Google Scholar]
- Martinsson, E.; Hansson, H.; Mittenzwei, K.; Storm, H. Evaluating Environmental Effects of Adopting Automatic Milking Systems on Norwegian Dairy Farms. Eur. Rev. Agric. Econ. 2024, 51, 128–156. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty Years of Research on Rumen Methanogenesis: Lessons Learned and Future Challenges for Mitigation. Animal 2020, 14, s2–s16. [Google Scholar] [CrossRef] [PubMed]
- Marçal-Pedroza, M.G.; Campos, M.M.; Sacramento, J.P.; Pereira, L.G.R.; Machado, F.S.; Tomich, T.R.; Paranhos da Costa, M.J.R.; Sant’Anna, A.C. Are Dairy Cows with a More Reactive Temperament Less Efficient in Energetic Metabolism and Do They Produce More Enteric Methane? Animal 2021, 15, 100224. [Google Scholar] [CrossRef]
- Lovarelli, D.; Bacenetti, J.; Guarino, M. A Review on Dairy Cattle Farming: Is Precision Livestock Farming the Compromise for an Environmental, Economic and Social Sustainable Production? J. Clean. Prod. 2020, 262, 121409. [Google Scholar] [CrossRef]
- Bokde, N.D.; Milkevych, V.; Nielsen, R.K.; Villumsen, T.M.; Sahana, G. A Novel Approach for Anomaly Detection in Dairy Cow Gas Emission Records. Comput. Electron. Agric. 2023, 214, 108286. [Google Scholar] [CrossRef]
- Niloofar, P.; Francis, D.P.; Lazarova-Molnar, S.; Vulpe, A.; Vochin, M.-C.; Suciu, G.; Balanescu, M.; Anestis, V.; Bartzanas, T. Data-Driven Decision Support in Livestock Farming for Improved Animal Health, Welfare and Greenhouse Gas Emissions: Overview and Challenges. Comput. Electron. Agric. 2021, 190, 106406. [Google Scholar] [CrossRef]
- Neethirajan, S. SOLARIA-SensOr-Driven resiLient and Adaptive monitoRIng of Farm Animals. Agriculture 2023, 13, 436. [Google Scholar] [CrossRef]
- Neethirajan, S. Net Zero Dairy Farming—Advancing Climate Goals with Big Data and Artificial Intelligence. Climate 2024, 12, 15. [Google Scholar] [CrossRef]
- Puppel, K.; Kuczyńska, B. Metabolic Profiles of Cow’s Blood; a Review. J. Sci. Food Agric. 2016, 96, 4321–4328. [Google Scholar] [CrossRef]
- Shibata, M.; Terada, F. Factors Affecting Methane Production and Mitigation in Ruminants. Anim. Sci. J. 2010, 81, 2–10. [Google Scholar] [CrossRef]
- Paudyal, S. Using Rumination Time to Manage Health and Reproduction in Dairy Cattle: A Review. Vet. Q. 2021, 41, 292–300. [Google Scholar] [CrossRef]
- Lopes, L.S.F.; Schenkel, F.S.; Houlahan, K.; Rochus, C.M.; Oliveira, G.A.; Oliveira, H.R.; Miglior, F.; Alcantara, L.M.; Tulpan, D.; Baes, C.F. Estimates of Genetic Parameters for Rumination Time, Feed Efficiency, and Methane Production Traits in First Lactation Holstein Cows. J. Dairy Sci. 2024, in press. [Google Scholar] [CrossRef]
- Zetouni, L.; Difford, G.F.; Lassen, J.; Byskov, M.V.; Norberg, E.; Løvendahl, P. Is Rumination Time an Indicator of Methane Production in Dairy Cows? J. Dairy Sci. 2018, 101, 11074–11085. [Google Scholar] [CrossRef] [PubMed]
- Lassen, J.; Difford, G.F. Review: Genetic and Genomic Selection as a Methane Mitigation Strategy in Dairy Cattle. Animal 2020, 14, s473–s483. [Google Scholar] [CrossRef] [PubMed]
- Smith, O.A.; Rochus, C.M.; Baes, C.F.; van Staaveren, N. A Note on Dairy Cow Behavior When Measuring Enteric Methane Emissions with the GreenFeed Emission Monitoring System in Tie-Stalls. JDS Commun. 2023, in press. [Google Scholar] [CrossRef]
- López-Paredes, J.; Goiri, I.; Atxaerandio, R.; García-Rodríguez, A.; Ugarte, E.; Jiménez-Montero, J.A.; Alenda, R.; González-Recio, O. Mitigation of Greenhouse Gases in Dairy Cattle via Genetic Selection: 1. Genetic Parameters of Direct Methane Using Noninvasive Methods and Proxies of Methane. J. Dairy Sci. 2020, 103, 7199–7209. [Google Scholar] [CrossRef] [PubMed]
- Mikuła, R.; Pszczola, M.; Rzewuska, K.; Mucha, S.; Nowak, W.; Strabel, T. The Effect of Rumination Time on Milk Performance and Methane Emission of Dairy Cows Fed Partial Mixed Ration Based on Maize Silage. Animals 2022, 12, 50. [Google Scholar] [CrossRef]
- Watt, L.J.; Clark, C.E.F.; Albornoz, R.I.; Krebs, G.L.; Petzel, C.E.; Utsumi, S.A. Effects of Grain-Based Concentrate Feeding and Rumination Frequency on the Milk Production, Methane and Carbon Dioxide Fluxes, and Activity of Dairy Cows in a Pasture-Based Automatic Milking System. Livest. Sci. 2021, 245, 104420. [Google Scholar] [CrossRef]
- Beauchemin, K.A. Invited Review: Current Perspectives on Eating and Rumination Activity in Dairy Cows. J. Dairy Sci. 2018, 101, 4762–4784. [Google Scholar] [CrossRef]
- Beauchemin, K.A. Ingestion and Mastication of Feed by Dairy Cattle. Vet. Clin. N. Am. Food Anim. Pract. 1991, 7, 439–463. [Google Scholar] [CrossRef]
- Aditya, S.; Bahutala, M.B.; Hibatullah, D.N.; Pourazad, P.; Wahyono, T.; Qumar, M.; Penagos-Tabares, F.; Wulansari, N. Evaluation of Milk Yield and Composition, Feed Intake, Chewing Activities, and Clinical Variables in Dairy Cows under Hot-Humid Climate of Tropical Zone. J. Therm. Biol. 2023, 114, 103608. [Google Scholar] [CrossRef]
- Llonch, P.; Somarriba, M.; Duthie, C.-A.; Haskell, M.J.; Rooke, J.A.; Troy, S.; Roehe, R.; Turner, S.P. Association of Temperament and Acute Stress Responsiveness with Productivity, Feed Efficiency, and Methane Emissions in Beef Cattle: An Observational Study. Front. Vet. Sci. 2016, 3, 43. [Google Scholar] [CrossRef]
- Dumortier, P.; Gourlez de la Motte, L.; Andriamandroso, A.L.H.; Aubinet, M.; Beckers, Y.; Bindelle, J.; De Cock, N.; Lebeau, F.; Heinesch, B. Beef Cattle Methane Emission Estimation Using the Eddy Covariance Technique in Combination with Geolocation. Agric. For. Meteorol. 2021, 297, 108249. [Google Scholar] [CrossRef]
- Salles, M.S.V.; da Silva, S.C.; Salles, F.A.; Roma, L.C.; El Faro, L.; Bustos Mac Lean, P.A.; Lins de Oliveira, C.E.; Martello, L.S. Mapping the Body Surface Temperature of Cattle by Infrared Thermography. J. Therm. Biol. 2016, 62, 63–69. [Google Scholar] [CrossRef]
- Türkgeldi, B.; Koç, F.; Lackner, M.; Okuyucu, B.; Okur, E.; Palangi, V.; Esen, S. Infrared Thermography Assessment of Aerobic Stability of a Total Mixed Ration: An Innovative Approach to Evaluating Dairy Cow Feed. Animals 2023, 13, 2225. [Google Scholar] [CrossRef]
- Wang, F.-K.; Shih, J.-Y.; Juan, P.-H.; Su, Y.-C.; Wang, Y.-C. Non-Invasive Cattle Body Temperature Measurement Using Infrared Thermography and Auxiliary Sensors. Sensors 2021, 21, 2425. [Google Scholar] [CrossRef]
- Guadagnin, A.R.; Matiello, J.P.; Ribeiro, R.S.; Pereira, L.G.R.; Machado, F.S.; Tomich, T.R.; Campos, M.M.; Heisler, G.; Fischer, V. Assessment of Heat Production and Methane Emission Using Infrared Thermography in Lactating Holstein and Gyrolando-F1 (½ Holstein ½ Gyr) Crossbreed Cows. J. Therm. Biol. 2023, 115, 103628. [Google Scholar] [CrossRef]
- Montanholi, Y.R.; Odongo, N.E.; Swanson, K.C.; Schenkel, F.S.; McBride, B.W.; Miller, S.P. Application of Infrared Thermography as an Indicator of Heat and Methane Production and Its Use in the Study of Skin Temperature in Response to Physiological Events in Dairy Cattle (Bos Taurus). J. Therm. Biol. 2008, 33, 468–475. [Google Scholar] [CrossRef]
- Gabbi, A.M.; Kolling, G.J.; Fischer, V.; Pereira, L.G.R.; Tomich, T.R.; Machado, F.S.; Campos, M.M.; da Silva, M.V.G.B.; Cunha, C.S.; Santos, M.K.R.; et al. Use of Infrared Thermography to Estimate Enteric Methane Production in Dairy Heifers. Quant. InfraRed Thermogr. J. 2022, 19, 187–195. [Google Scholar] [CrossRef]
- Henchion, M.M.; Regan, Á.; Beecher, M.; MackenWalsh, Á. Developing ‘Smart’ Dairy Farming Responsive to Farmers and Consumer-Citizens: A Review. Animals 2022, 12, 360. [Google Scholar] [CrossRef] [PubMed]
- Giagnoni, G.; Friggens, N.C.; Johansen, M.; Maigaard, M.; Wang, W.; Lund, P.; Weisbjerg, M.R. How Much Can Performance Measures Explain of the Between-Cow Variation in Enteric Methane? J. Dairy Sci. 2024, in press. [Google Scholar] [CrossRef]
- González-Recio, O.; López-Paredes, J.; Ouatahar, L.; Charfeddine, N.; Ugarte, E.; Alenda, R.; Jiménez-Montero, J.A. Mitigation of Greenhouse Gases in Dairy Cattle via Genetic Selection: 2. Incorporating Methane Emissions into the Breeding Goal. J. Dairy Sci. 2020, 103, 7210–7221. [Google Scholar] [CrossRef] [PubMed]
- Saborío-Montero, A.; López-García, A.; Gutiérrez-Rivas, M.; Atxaerandio, R.; Goiri, I.; García-Rodriguez, A.; Jiménez-Montero, J.A.; González, C.; Tamames, J.; Puente-Sánchez, F.; et al. A Dimensional Reduction Approach to Modulate the Core Ruminal Microbiome Associated with Methane Emissions via Selective Breeding. J. Dairy Sci. 2021, 104, 8135–8151. [Google Scholar] [CrossRef]
- Manzanilla-Pech, C.I.V.; Difford, G.F.; Løvendahl, P.; Stephansen, R.B.; Lassen, J. Genetic (Co-)Variation of Methane Emissions, Efficiency, and Production Traits in Danish Holstein Cattle along and across Lactations. J. Dairy Sci. 2022, 105, 9799–9809. [Google Scholar] [CrossRef] [PubMed]
- Pszczola, M.; Strabel, T.; Mucha, S.; Sell-Kubiak, E. Genome-Wide Association Identifies Methane Production Level Relation to Genetic Control of Digestive Tract Development in Dairy Cows. Sci. Rep. 2018, 8, 15164. [Google Scholar] [CrossRef] [PubMed]
- Buddle, B.M.; Denis, M.; Attwood, G.T.; Altermann, E.; Janssen, P.H.; Ronimus, R.S.; Pinares-Patiño, C.S.; Muetzel, S.; Neil Wedlock, D. Strategies to Reduce Methane Emissions from Farmed Ruminants Grazing on Pasture. Vet. J. 2011, 188, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Obergassel, W.; Mersmann, F.; Ott, H.E.; Wang-Helmreich, H. Phoenix from the Ashes: An Analysis of the Paris Agreement to the United Nations Framework Convention on Climate Change—Part I; Environmental Law; Wuppertal Institute for Climate, Environment and Energy: Wuppertal, Germany, 2016. [Google Scholar]
- Olijhoek, D.W.; Løvendahl, P.; Lassen, J.; Hellwing, A.L.F.; Höglund, J.K.; Weisbjerg, M.R.; Noel, S.J.; McLean, F.; Højberg, O.; Lund, P. Methane Production, Rumen Fermentation, and Diet Digestibility of Holstein and Jersey Dairy Cows Being Divergent in Residual Feed Intake and Fed at 2 Forage-to-Concentrate Ratios. J. Dairy Sci. 2018, 101, 9926–9940. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, T.; Lima, M.A.; dos Santos, G.B.; Pereira, L.G.R.; Machado, F.S.; Tomich, T.R.; Campos, M.M.; Jonker, A.; Rodrigues, P.H.M.; Brandao, V.L.N.; et al. Effects of Feeding Level and Breed Composition on Intake, Digestibility, and Methane Emissions of Dairy Heifers. Animals 2021, 11, 586. [Google Scholar] [CrossRef]
- Lakamp, A.D.; Weaber, R.L.; Bormann, J.M.; Rolf, M.M. Relationships between Enteric Methane Production and Economically Important Traits in Beef Cattle. Livest. Sci. 2022, 265, 105102. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Economic Cost | Milk Yield | c Possible Impact on GHG per Unit of Milk or Meat Produce | References |
|---|---|---|---|---|
| Bovine viral diarrhoea virus | b Up to EUR 294.48 per cow (with an average of €54.34 per cow) | Reduced | Moderate | [43] |
| Infectious bovine rhinotracheitis | b EUR 233.71 per sub-clinically infected cow | Reduced | High | [43] |
| Bovine respiratory disease | EUR 21.77 per case | N/A | Low | [43,44] |
| Johne’s disease | b EUR 30.38 per dairy cow and EUR 19.87 per beef cow | Reduced | High | [43] |
| Mastitis | Chronic EUR 118. Quartile range EUR 106–132 per case, clinical EUR 240/lactating cow per year | Reduced | Moderate | [43,45,46] |
| Lameness | From EUR 214.51 to 992.10 per case | Reduced | Moderate | [43,47,48] |
| Subclinical ketosis | EUR 179.37 per calving cow | Reduced | Low | [49,50,51,52] |
| Clinical ketosis | single clinical ketosis case averaged EUR 709 | Reduced | N/A | [42,53,54] |
| SARA | a of EUR 368.59 to EUR 437.71 lost income per cow per year | Reduced | N/A | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Džermeikaitė, K.; Krištolaitytė, J.; Antanaitis, R. Relationship between Dairy Cow Health and Intensity of Greenhouse Gas Emissions. Animals 2024, 14, 829. https://doi.org/10.3390/ani14060829
Džermeikaitė K, Krištolaitytė J, Antanaitis R. Relationship between Dairy Cow Health and Intensity of Greenhouse Gas Emissions. Animals. 2024; 14(6):829. https://doi.org/10.3390/ani14060829
Chicago/Turabian StyleDžermeikaitė, Karina, Justina Krištolaitytė, and Ramūnas Antanaitis. 2024. "Relationship between Dairy Cow Health and Intensity of Greenhouse Gas Emissions" Animals 14, no. 6: 829. https://doi.org/10.3390/ani14060829
APA StyleDžermeikaitė, K., Krištolaitytė, J., & Antanaitis, R. (2024). Relationship between Dairy Cow Health and Intensity of Greenhouse Gas Emissions. Animals, 14(6), 829. https://doi.org/10.3390/ani14060829