
Movement and Home Range of Amur Soft-Shell Turtle (Pelodiscus maackii) in the Ussuri River, Heilongjiang Province, China
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
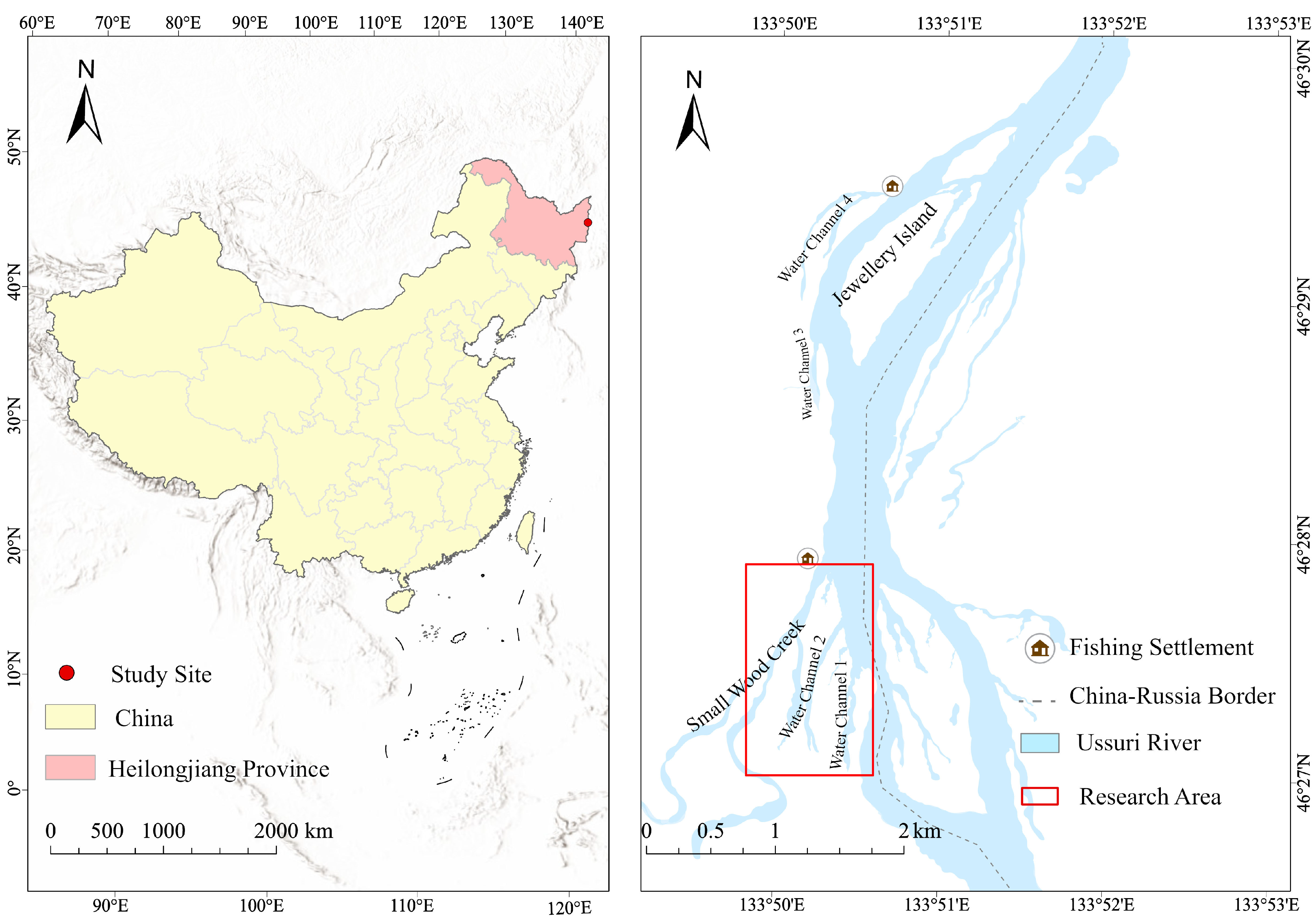
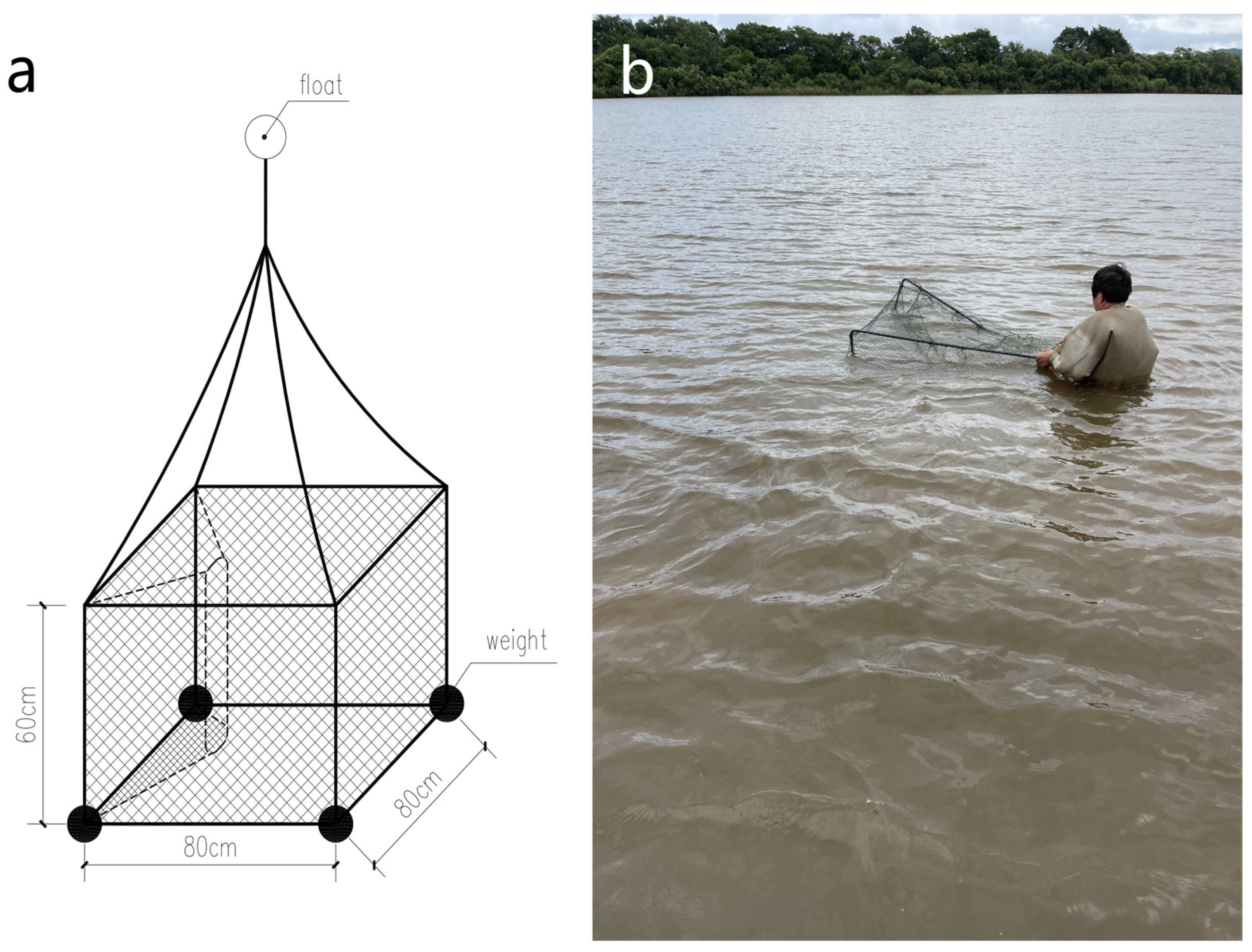
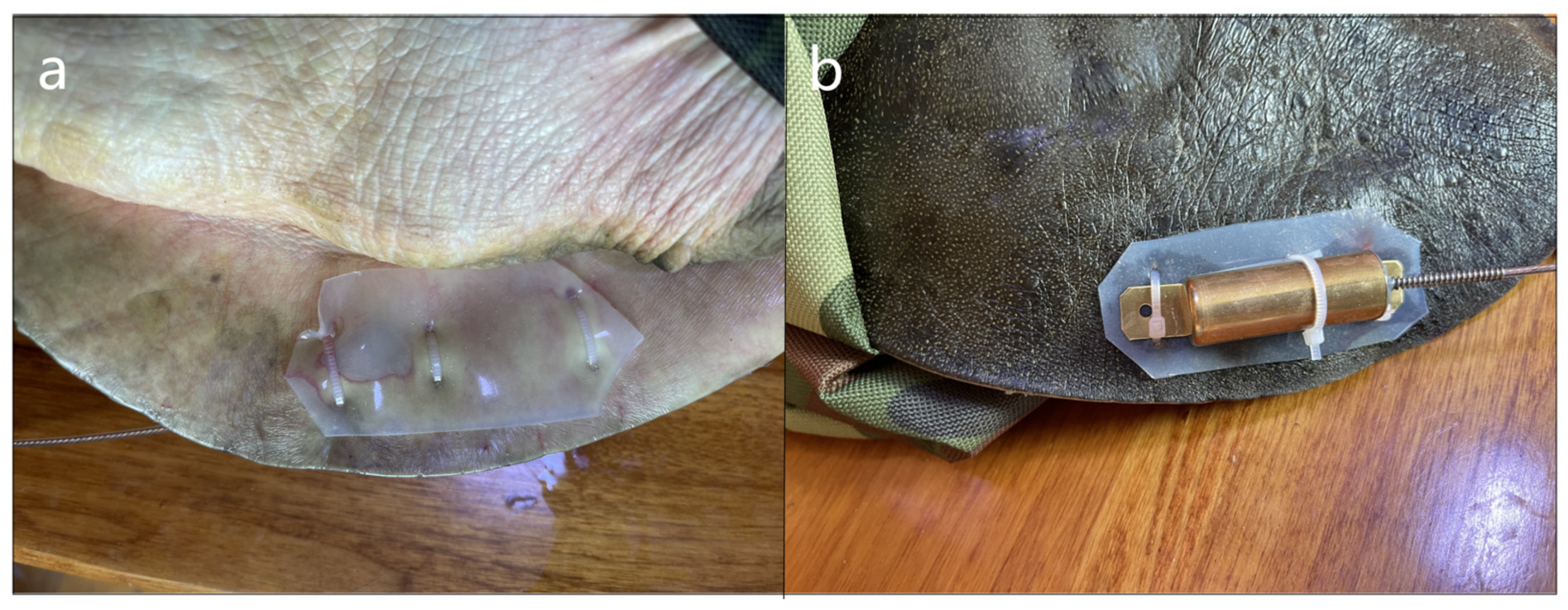
2.1. Study Site, Radio Telemetry, and Data Collection
2.2. Home Range Analysis
2.3. Movement Analysis
3. Results
3.1. Home Range Analysis
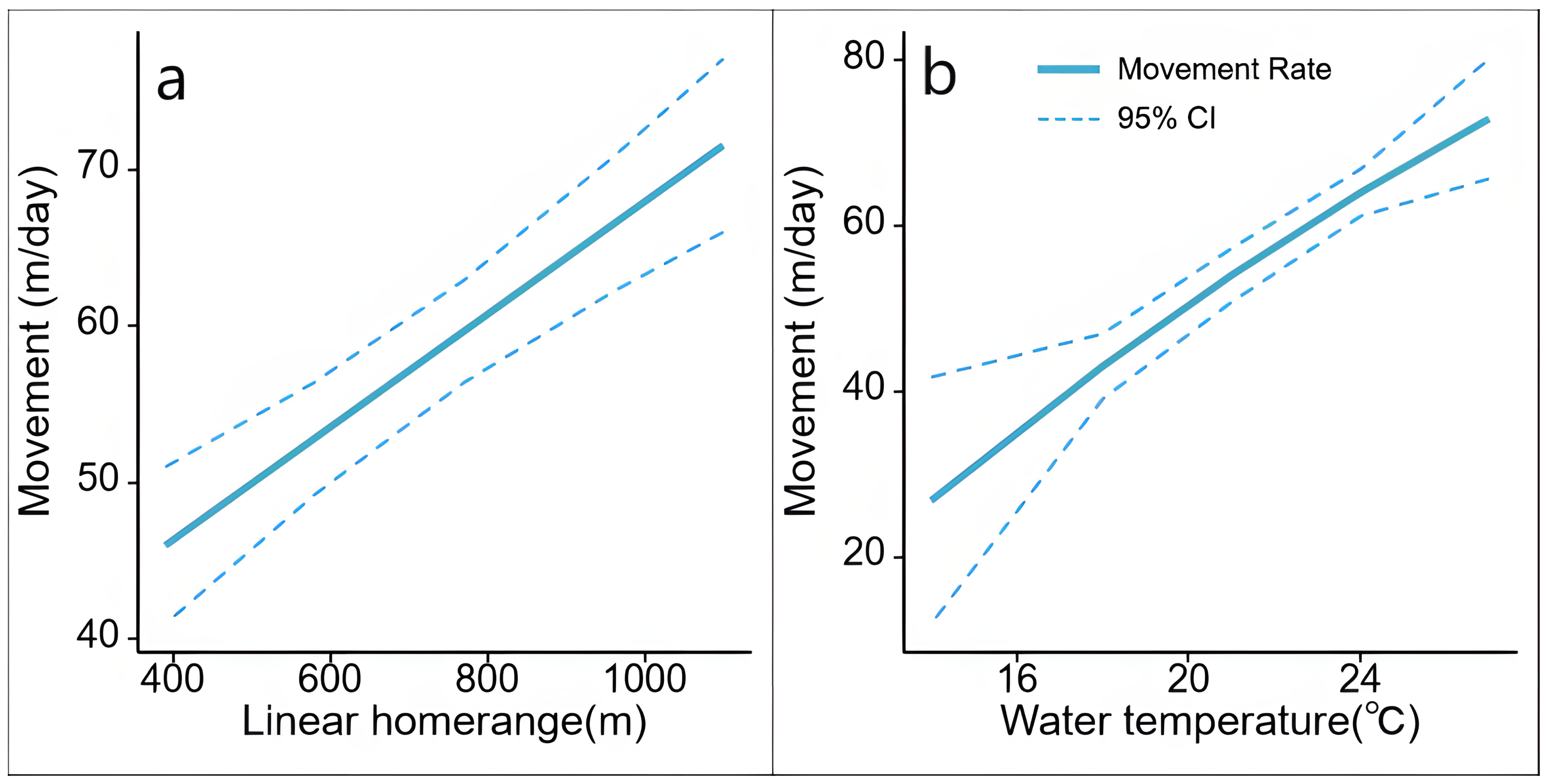
3.2. Movement Analysis
4. Discussion
4.1. Home Range
4.2. Movement
4.3. Conservation and Management
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Age Group | Sex | PL | Body Mass | Locations | Linear Range | Min Move | Max Move | Mean Move | Move SD | MCP | KDE 95% | KDE 50% | Prop. Asy. | End |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Subadult | Male | 13.8 | 560 | 68 | 615 | 14 | 137.4 | 60.25 | 28.77 | 1.79 | 0.86 | 0.09 | 1.01 | Complete home range, released at the end |
| 2 | Subadult | Male | 14 | 590 | 55 | 672 | 10.8 | 118.9 | 61.58 | 26.08 | 1.77 | 0.68 | 0.08 | 1.01 | Complete home range, caught by fish net and released |
| 3 | Subadult | Male | 13 | 520 | 59 | 624 | 16.1 | 113.8 | 57.86 | 23.83 | 1.65 | 0.97 | 0.11 | 1.01 | Complete home range, released at the end |
| 4 | Subadult | Female | 13.6 | 580 | 67 | 1145 | 18.6 | 275.6 | 72.45 | 29.36 | 3.65 | 2.07 | 0.18 | 0.96 | Complete home range, released at the end |
| 5 | Subadult | Female | 14.3 | 617 | 62 | 388 | 10.8 | 89.9 | 48.12 | 19.64 | 1.30 | 0.66 | 0.07 | 1.01 | Complete home range, released at the end |
| 6 | Subadult | Female | 13.5 | 550 | 70 | 995 | 15.2 | 158.5 | 62.25 | 26.41 | 1.80 | 0.78 | 0.08 | 1.03 | Complete home range, released at the end |
| 7 | Subadult | Male | 14.2 | 615 | 75 | 632 | 9.2 | 133.7 | 49.77 | 22.58 | 2.71 | 1.32 | 0.11 | 0.92 | Complete home range, released at the end |
| 8 | Subadult | Female | 13.9 | 600 | 65 | 473 | 8.5 | 94.6 | 39.18 | 20.04 | 1.58 | 0.78 | 0.07 | 1.01 | Complete home range, released at the end |
| 9 | Subadult | Female | 14.6 | 680 | 29 | * | * | * | * | * | * | * | * | 0.58 | |
| 10 | Subadult | Female | 14 | 600 | 18 | * | * | * | * | * | * | * | * | 0.68 | Incomplete home range, radio transmitter peeled off by fish net and turtle released |
| 11 | Subadult | Female | 13.7 | 578 | 30 | * | * | * | * | * | * | * | * | 0.70 | Incomplete home range, radio transmitter peeled off by fish net and turtle escaped |
| 12 | Subadult | Male | 14.5 | 620 | 32 | * | * | * | * | * | * | * | * | 0.69 | Incomplete home range, radio transmitter peeled off for unknown reason |
| 13 | Adult | Female | 16.8 | 1000 | 14 | * | 98.2 | 345.3 | 178.2 | * | * | * | 0.45 | Incomplete home range, signal stayed on Russia side | |
| 14 | Adult | Male | 17 | 1230 | 11 | * | 89.5 | 420.2 | 159.6 | * | * | * | 0.50 | Incomplete home range, signal disappeared after 11 locations | |
| 15 | Adult | Female | 22 | 2000 | 1 | * | * | * | * | * | * | * | * | * | Signal disappeared on the second day |
| 16 | Adult | Female | 26 | 2720 | 1 | * | * | * | * | * | * | * | * | * | Signal disappeared on the second day |
| 17 | Adult | Male | 19.5 | 1880 | 3 | * | * | * | * | * | * | * | * | * | Signal disappeared on the fourth day |
| 18 | Adult | Male | 24.5 | 2450 | 1 | * | * | * | * | * | * | * | * | * | Signal disappeared on the second day |
| 19 | Adult | Female | 29 | 2950 | 1 | * | * | * | * | * | * | * | * | * | Signal disappeared on the second day |
Appendix B
| Model Name | Structure |
|---|---|
| Global | 1 + W + W2 + LR + Sex + PL + W × LR + W × PL + W × Sex + 1|id |
| Temp. and linear home range interaction | 1 + W + W2 + LR + W × LR + 1|id |
| Temp., linear home range, and sex interaction | 1 + W + W2 + LR + Sex + W × LR + W × Sex + 1|id |
| Temp., linear home range, and body size interaction | 1 + W + W2 + LR + PL + W × LR + W × PL + 1|id |
| Temp., sex, and body size interaction | 1 + W + W2 + Sex + PL + W × Sex + W × PL + 1|id |
| Temp. and linear home range | 1 + W + W2 + LR + 1|id |
| Temp. and linear home range linear effect and interaction | 1 + W + LR + W × LR + 1|id |
| Temp. | 1 + W + W2 + 1|id |
| Temp., sex, and body size | 1 + W + W2 + Sex + PL + 1|id |
| Main linear effect | 1 + W + LR + Sex + PL + 1|id |
| Temp. and sex interaction | 1 + W + W2 + Sex + W × Sex + 1|id |
| Temp. and body size interaction | 1 + W + W2 + PL + W × PL + 1|id |
| Temp. and linear home range linear effect | 1 + W + LR + 1|id |
| Temp., sex, and body size linear effect | 1 + W + Sex + PL + 1|id |
| Temp. and sex | 1 + W + W2 + Sex + 1|id |
| Temp. and body size | 1 + W + W2 + PL + 1|id |
| Null | 1 + 1|id |
References
- Brandt, J.F. Observationes quaedam ad generis trionychum species duas novas spectantes. Bull. Acad. Imper. Sci. St. Petersbourg Cl. Phys.-Mathemat. 1857, 16, 110–111. [Google Scholar]
- Stejneger, L. Herpetology of Japan and Adjacent Territory; Government Printing Office: Washington, DC, USA, 1907. [Google Scholar]
- Baek, H.-J.; Kim, P.; Kim, Y.-C.; Kim, A.; Kim, S.; Min, M.-S.; Lee, H. The complete mitochondrial genome of the Amur soft-shelled turtle (Pelodiscus maackii Brandt, 1858), from South Korea. Mitochondrial DNA Part B 2022, 7, 498–500. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.H.; Song, J.Y.; Koo, K.S. The Status of Distribution for Native Freshwater Turtles in Korea, with Remarks on Taxonomic Position. Korean J. Environ. Biol. 2012, 30, 151–155. [Google Scholar]
- Suzuki, D.; Hikida, T. Taxonomic status of the soft-shell turtle populations in Japan: A molecular approach. Curr. Herpetol. 2014, 33, 171–179. [Google Scholar] [CrossRef]
- Moll, D.; Moll, E.O. The Ecology, Exploitation and Conservation of River Turtles; Oxford University Pres: New York, NY, USA, 2004; pp. 1–393. [Google Scholar]
- ESRI. ArcGis Pro 3.0.1; Environmental Systems Research Institute: Redlands, CA, USA, 2022. [Google Scholar]
- Sexton, O.J. Spatial and temporal movements of a population of the painted turtle, Chrysemys picta marginata (Agassiz). Ecol. Monogr. 1959, 29, 113–140. [Google Scholar] [CrossRef]
- Pluto, T.G.; Bellis, E.D. Seasonal and annual movements of riverine map turtles, Graptemys geographica. J. Herpetol. 1988, 22, 152–158. [Google Scholar] [CrossRef]
- Nilsen, E.B.; Pedersen, S.; Linnell, J.D.C. Can minimum convex polygon home ranges be used to draw biologically meaningful conclusions? Ecol. Res. 2008, 23, 635–639. [Google Scholar] [CrossRef]
- Jennrich, R.I.; Turner, F.B. Measurement of non-circular home range. J. Theor. Biol. 1969, 22, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Worton, B.J. A review of models of home range for animal movement. Ecol. Model. 1987, 38, 277–298. [Google Scholar] [CrossRef]
- Worton, B.J. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 1989, 70, 164–168. [Google Scholar] [CrossRef]
- Silverman, B.W. Density Estimation for Statistics and Data Analysis; Chapman & Hall: London, UK, 1986. [Google Scholar]
- Worton, B.J. Using Monte Carlo simulation to evaluate kernel-based home range estimators. J. Wildl. Manag. 1995, 59, 794–800. [Google Scholar] [CrossRef]
- Park, B.U.; Marron, J.S. Comparison of data-driven bandwidth selectors. J. Am. Stat. Assoc. 1990, 85, 66–72. [Google Scholar] [CrossRef]
- Sain, S.R.; Baggerly, K.A.; Scott, D.W. Cross-validation of multivariate densities. J. Am. Stat. Assoc. 1994, 89, 807–817. [Google Scholar] [CrossRef]
- Jones, M.C.; Marron, J.S.; Sheather, S.J. A brief survey of bandwidth selection for density estimation. J. Am. Stat. Assoc. 1996, 91, 401–407. [Google Scholar] [CrossRef]
- Horne, J.S.; Garton, E.O. Likelihood cross-validation versus least squares cross-validation for choosing the smoothing parameter in kernel home-range analysis. J. Wildl. Manag. 2006, 70, 641–648. [Google Scholar] [CrossRef]
- Horne, J.S.; Garton, E.O. Animal Space Use 1.3. 2015. Available online: http://www.webpages.uidaho.edu/population_ecology/animal_space_use.htm (accessed on 13 February 2015).
- Row, J.R.; Blouin-Demers, G. Kernels are not accurate estimators of home-range size for herpetofauna. Copeia 2006, 2006, 797–802. [Google Scholar] [CrossRef]
- Blundell, G.M.; Maier, J.A.K.; Debevec, E.M. Linear home ranges: Effects of smoothing, sample size, and autocorrelation on kernel estimates. Ecol. Monogr. 2001, 71, 469–489. [Google Scholar] [CrossRef]
- Hodder, K.H.; Masters, J.E.G.; Beaumont, W.R.C.; Gozlan, R.E.; Pinder, A.C.; Knight, C.M.; Kenward, R.E. Techniques for evaluating the spatial behaviour of river fish. Hydrobiologia 2007, 582, 257–269. [Google Scholar] [CrossRef]
- Carrière, M.A. Movement Patterns and Habitat Selection of Common Map Turtles (Graptemys geographica) in St. Lawrence Islands National Park, Ontario, Canada. Master’s Thesis, University of Ottawa, Ottawa, ON, Canada, 2007. [Google Scholar]
- Geffen, E.; Mendelssohn, H. Home range use and seasonal movements of the Egyptian tortoise (Testudo kleinmanni) in the northwestern Negev, Israel. Herpetologica 1988, 44, 354–359. [Google Scholar]
- Calenge, C. The package adehabitat for the R software: A tool for the analysis of space and habitat use by animals. Ecol. Model 2006, 197, 516–519. [Google Scholar] [CrossRef]
- Harris, S.; Cresswell, W.J.; Forde, P.G.; Trewhella, W.J.; Woollard, T.; Wray, S. Home-range analysis using radio-tracking data–a review of problems and techniques particularly as applied to the study of mammals. Mammal Rev. 1990, 20, 97–123. [Google Scholar] [CrossRef]
- Kranstauber, B.; Smolla, M.; Scharf, A. Move: Visualizing and Analyzing Animal Track Data, R Package Version 4.2.2; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Koya, P.R.; Goshu, A.T. Solutions of rate-state equation describing biological growths. Am. J. Math. Stat. 2013, 3, 305–311. [Google Scholar]
- Ernst, C.H.; Lovich, J.E. Turtles of the United States and Canada, 2nd ed.; The Johns Hopkins University Pres: Baltimore, MD, USA, 2009; pp. 1–827. [Google Scholar]
- Bodie, J.R.; Semlitsch, R.D. Spatial and temporal use of floodplain habitats by lentic and lotic species of aquatic turtles. Oecologia 2000, 122, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Naimi, B.; Hamm, N.A.S.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where is positional uncertainty a problem for species distribution modelling? Ecography 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 3 September 2021).
- Bates, M.M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Mazerolle, M.J. Model selection and multimodel inference using the AICcmodavg package. R Vignette 2020, 2020, 22. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information—Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; pp. 1–488. [Google Scholar]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Barton, B.K. MuMIn: Multi-Model Inference. R Package Version 1.42.1. 2019. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 6 April 2019).
- Plummer, M.V.; Shirer, H.W. Movement Patterns in A River Population of the Softshell Turtle, Trionyx Muticus; Occasional Papers of the Museum of Natural History 1975; The University of Kansas: Lawrence, KS, USA, 1975; Volume 43, pp. 1–26. [Google Scholar]
- Ross, J.P.; Bluett, R.D.; Dreslik, M.J. Movement and home range of the smooth softshell turtle (Apalone mutica): Spatial ecology of a river specialist. Diversity 2019, 11, 124. [Google Scholar] [CrossRef]
- Powell, R.A. Animal home ranges and territories and home range estimators. Res. Tech. Anim. Ecol. Controv. Conseq. 2000, 442, 65–110. [Google Scholar]
- Kenward, R.E. A Manual for Wildlife Radio Tagging; Academic Press: London, UK, 2001; pp. 1–311. [Google Scholar]
- Steury, T.D.; Mccarthy, J.E.; Roth, T.C.; Lima, S.L.; Murray, D.L. Evaluation of Root-n Bandwidth Selectors for Kernel Density Estimation. J. Wildl. Manag. 2010, 74, 539–548. [Google Scholar] [CrossRef]
- Börger, L.; Dalziel, B.D.; Fryxell, J.M. Are there general mechanisms of animal home range behaviour? A review and prospects for future research. Ecol. Lett. 2008, 11, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Mueller, T.; Fagan, W.F. Search and navigation in dynamic environments–from individual behaviors to population distributions. Oikos 2008, 117, 654–664. [Google Scholar] [CrossRef]
- Svanbäck, R.; Bolnick, D.I. Intraspecific competition affects the strength of individual specialization: An optimal diet theory method. Evol. Ecol. Res. 2005, 7, 993–1012. [Google Scholar]
- Swihart, R.K.; Slade, N.A. Influence of sampling interval on estimates of home-range size. J. Wildl. Manag. 1985, 49, 1019–1025. [Google Scholar] [CrossRef]
- Kernohan, B.J.; Gitzen, R.A.; Millspaugh, J.J. Analysis of animal space use and movements. In Radio Tracking and Animal Populations; Millspaugh, J.J., Marzluff, J.M., Eds.; Academic Press: San Diego, CA, USA, 2001; pp. 125–166. [Google Scholar]
- Woolnough, D.A.; Downing, J.A.; Newton, T.J. Fish movement and habitat use depends on water body size and shape. Ecol. Freshw. Fish 2009, 18, 83–91. [Google Scholar] [CrossRef]
- Duncan, M.B. Distributions of small nongame fishes in the lower Yellowstone River. Am. Midl. Nat. 2016, 175, 1–23. [Google Scholar] [CrossRef]
- Gibbons, J.W. Movement patterns among turtle populations: Applicability to management of the desert tortoise. Herpetologica 1986, 42, 104–113. [Google Scholar]
- Plummer, M.V. Activity, habitat and population structure in the turtle. Trionyx muticus. Copeia 1977, 1977, 431. [Google Scholar] [CrossRef]
- Lovich, J.E. Geographic variation in the seasonal activity cycle of spotted turtles. Clemmys Guttata. J. Herpetol. 1988, 22, 482–485. [Google Scholar] [CrossRef]
- Ernst, C.H. Environmental temperatures and activities in wild spotted turtles. Clemmys Guttata. J. Herpetol. 1982, 16, 112–120. [Google Scholar] [CrossRef]
- Plummer, M.V.; Mills, N.E.; Allen, S.L. Activity, habitat, and movement patterns of softshell turtles (Trionyx spiniferus) in a small stream. Chelonian Conserv. Biol. 1997, 2, 514–520. [Google Scholar]
- Gibbons, J.W. The slider turtle. In Life History and Ecology of the Slider Turtle; Gibbons, J.W., Ed.; Smithsonian Institution Press: Washington, DC, USA, 1990; pp. 3–18. [Google Scholar]

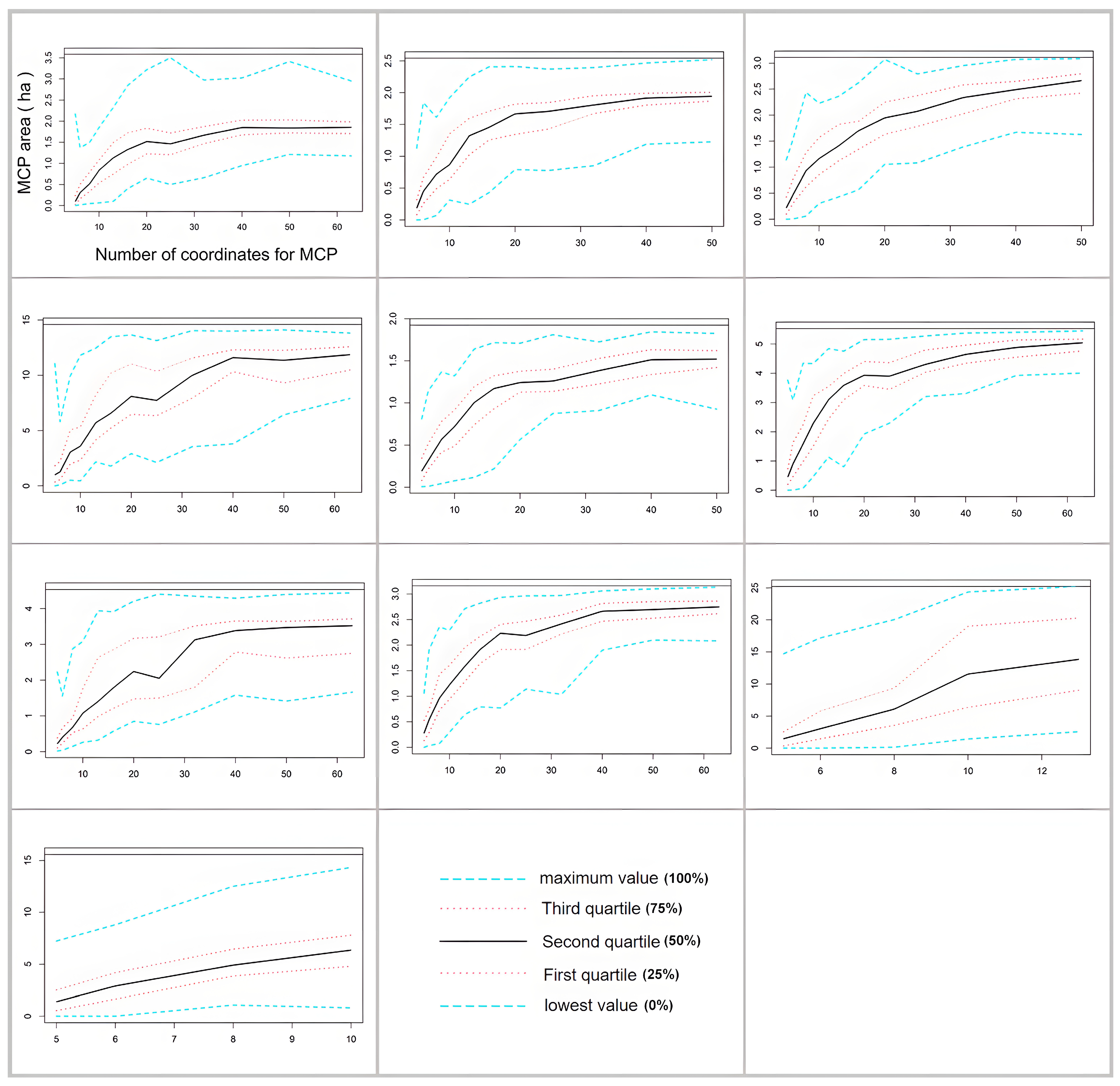
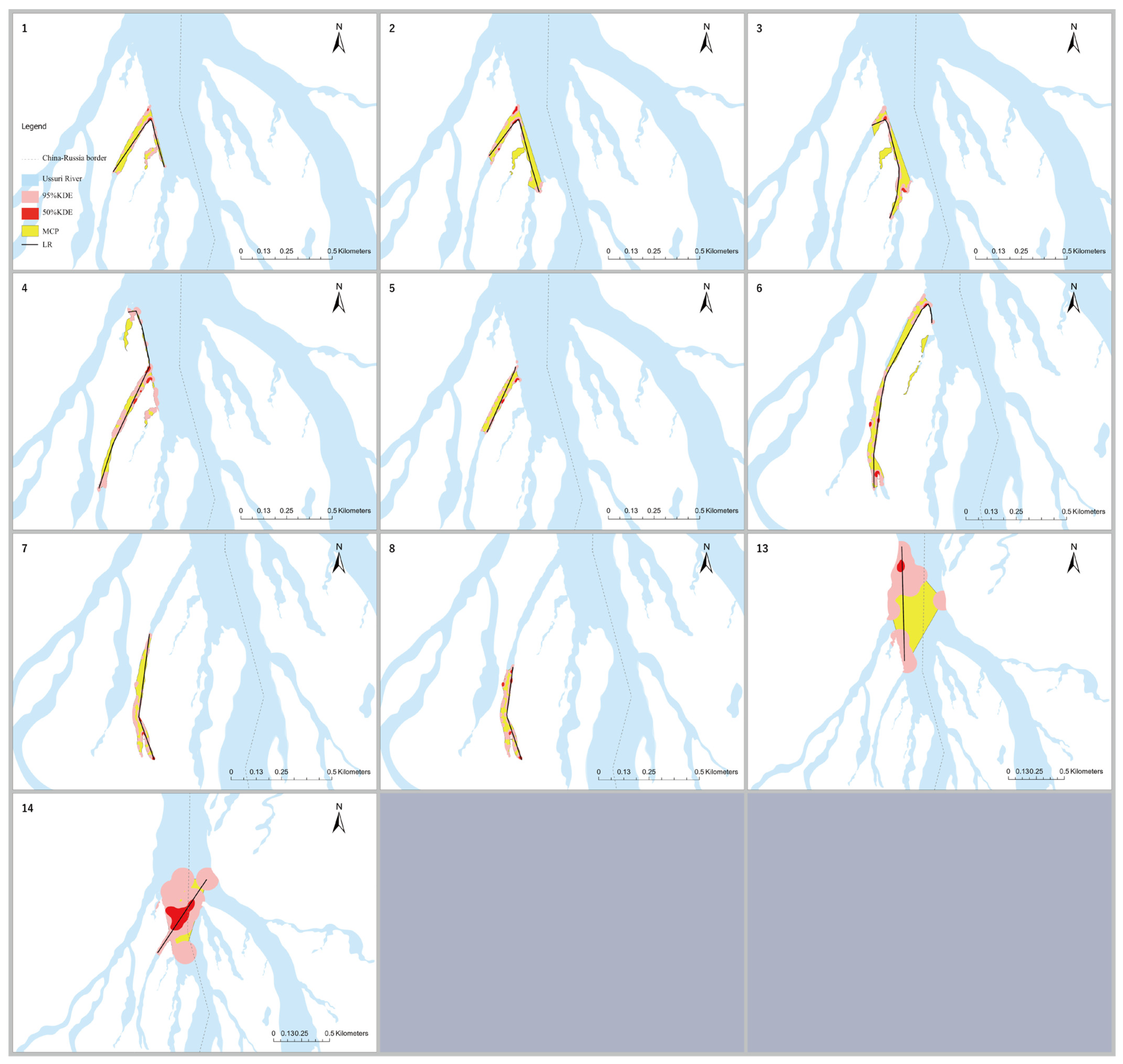
| MCP | ||||||||
| ID | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| 1 | 88.89 | 55.84 | 20.53 | 84.04 | 38.5 | 0 | 0 | |
| 2 | 63.01 | 80.14 | 17.38 | 73.08 | 30.81 | 0 | 0 | |
| 3 | 48.42 | 31.97 | 15.96 | 17.4 | 15.67 | 0 | 0 | |
| 4 | 83.63 | 70.84 | 37.42 | 100 | 64.68 | 41.49 | 8.55 | |
| 5 | 38.12 | 45.89 | 10.77 | 13.18 | 29.86 | 0 | 0 | |
| 6 | 0 | 35.77 | 12.7 | 76.91 | 82.82 | 81.05 | 80.52 | |
| 7 | 0 | 0 | 0 | 6.67 | 0 | 56.49 | 90.47 | |
| 8 | 0 | 0 | 0 | 1.85 | 0 | 43.03 | 69.31 | |
| 95% KDE | ||||||||
| ID | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| 1 | 64.75 | 54.78 | 36.92 | 68.86 | 29.85 | 0 | 0 | |
| 2 | 0.73 | 49.79 | 31.49 | 76.58 | 31.14 | 0 | 0 | |
| 3 | 0.56 | 44.96 | 20.07 | 30.41 | 22.45 | 0 | 0 | |
| 4 | 0.78 | 86.75 | 61.26 | 97 | 43.09 | 24.1 | 19.83 | |
| 5 | 0.65 | 79.9 | 31.87 | 34.4 | 22.8 | 0 | 0 | |
| 6 | 0.35 | 42.38 | 0 | 40.31 | 45.56 | 82.19 | 72.53 | |
| 7 | 0 | 0 | 0 | 9.21 | 0 | 54.55 | 86.94 | |
| 8 | 0 | 0 | 0 | 7.46 | 0 | 47.11 | 69.66 | |
| 50% KDE | ||||||||
| ID | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| 1 | 51.3 | 43.31 | 28.63 | 35.97 | 24.32 | 0 | 0 | |
| 2 | 88.54 | 42.42 | 43.39 | 63.79 | 23.89 | 0 | 0 | |
| 3 | 80.45 | 47.29 | 23.3 | 28.81 | 22.76 | 0 | 0 | |
| 4 | 100 | 92.16 | 42.06 | 96.5 | 28.43 | 0 | 0 | |
| 5 | 77.09 | 76.88 | 41.43 | 39.47 | 23.45 | 0 | 0 | |
| 6 | 0 | 30.38 | 34.43 | 27.71 | 34.33 | 74.64 | 80.79 | |
| 7 | 0 | 0 | 0 | 0 | 0 | 33.17 | 45.04 | |
| 8 | 0 | 0 | 0 | 0 | 0 | 52.96 | 53.93 | |
| Rank | Model Name | K | −2LL | AICc | ∆AICc | wi | R2(m) | R2(c) |
|---|---|---|---|---|---|---|---|---|
| 1 | Global | 11 | −2284.17 | 4590.87 | 0 | 0.83 | 0.26 | 0.26 |
| 2 | Temp., linear home range, and sex interaction | 9 | −2288.68 | 4595.73 | 4.86 | 0.07 | 0.26 | 0.26 |
| 3 | Main linear effect | 7 | −2291.24 | 4596.71 | 5.84 | 0.04 | 0.25 | 0.26 |
| 4 | Temp. and linear home range interaction | 7 | −2292.5 | 4599.23 | 8.36 | 0.01 | 0.26 | 0.26 |
| 5 | Temp., linear home range, and body size interaction | 9 | −2290.54 | 4599.45 | 8.57 | 0.01 | 0.26 | 0.26 |
| 17 | Null | 3 | −2345.27 | 4696.58 | 105.71 | 0 | 0 | 0.15 |
| Effect | Estimate | SE | LCI | UCI |
|---|---|---|---|---|
| Intercept | 54.83 | 2.49 | 49.94 | 59.72 |
| Water Temp. | 12.6 | 1.92 | 8.84 | 16.37 |
| (Water Temp.)2 | −0.49 | 1.19 | −2.81 | 1.84 |
| Linear Home Range (LR) | 8.71 | 1.45 | 5.87 | 11.55 |
| Sex | 5.5 | 4.51 | −3.35 | 14.35 |
| Plastron Length (PL) | −2.35 | 1.97 | −6.22 | 1.51 |
| Water Temp × LR | −1.73 | 1.28 | −4.24 | 0.78 |
| Water Temp × PL | 2.8 | 2.05 | −1.21 | 6.82 |
| Water Temp × Sex | −7.1 | 4.74 | −16.39 | 2.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, X.; Shi, H. Movement and Home Range of Amur Soft-Shell Turtle (Pelodiscus maackii) in the Ussuri River, Heilongjiang Province, China. Animals 2024, 14, 1088. https://doi.org/10.3390/ani14071088
Hou X, Shi H. Movement and Home Range of Amur Soft-Shell Turtle (Pelodiscus maackii) in the Ussuri River, Heilongjiang Province, China. Animals. 2024; 14(7):1088. https://doi.org/10.3390/ani14071088
Chicago/Turabian StyleHou, Xiaochen, and Haitao Shi. 2024. "Movement and Home Range of Amur Soft-Shell Turtle (Pelodiscus maackii) in the Ussuri River, Heilongjiang Province, China" Animals 14, no. 7: 1088. https://doi.org/10.3390/ani14071088
APA StyleHou, X., & Shi, H. (2024). Movement and Home Range of Amur Soft-Shell Turtle (Pelodiscus maackii) in the Ussuri River, Heilongjiang Province, China. Animals, 14(7), 1088. https://doi.org/10.3390/ani14071088