Simple Summary
Outbreaks of white spot disease in Pacific white shrimp pose a significant threat to the major shrimp farming industry. Understanding the mechanisms of the shrimp against the causative virus is crucial. C-type lectins are important pattern recognition receptors that can be involved in the response against viral infections. This study identified the C-type lectin domain family 4 member F in shrimp as an important receptor gene that could promote replication of the causative virus and affect the survival rate of shrimp. This study will provide a theoretical basis for understanding the resistance mechanisms of shrimp against the virus.
Abstract
White spot disease (WSD) outbreaks pose a significant threat to the Pacific white shrimp (Litopenaeus vannamei) farming industry. The causative agent is the white spot syndrome virus (WSSV). There are no effective treatments for WSD so far. Therefore, understanding the resistance mechanisms of L. vannamei against the WSSV is crucial. C-type lectins (CTLs) are important pattern recognition receptors (PRRs) that promote agglutination, phagocytosis, encapsulation, bacteriostasis, and antiviral infections. This study cloned the C-type lectin domain family 4 member F (LvCLEC4F) from L. vannamei. LvCLEC4F contains a 492 bp open reading frame (ORF) encoding a protein of 163 amino acids, including a carbohydrate recognition domain (CRD). Following a challenge with the WSSV, the expression profile of LvCLEC4F was significantly altered. Using RNA interference (RNAi) technology, it was found that LvCLEC4F promotes WSSV replication and affects the expression levels of genes related to the regulation of apoptosis, signaling and cellular stress response, and immune defense. Meanwhile, the hemolymph agglutination phenomenon in vivo was weakened when LvCLEC4F was knocked down. These results indicated that LvCLEC4F may play an important role in the interaction between L. vannamei and WSSV.
1. Introduction
The Pacific white shrimp Litopenaeus vannamei has become a major shrimp farming species worldwide because of its rapid growth, strong disease resistance, and economic benefits [1]. With the expansion of aquaculture, sudden outbreaks of bacterial, fungal, and viral diseases pose a significant threat to the global shrimp farming industry. White spot disease (WSD) is one of the most serious diseases in shrimp, and the causative agent is the white spot syndrome virus (WSSV). WSSV, an enveloped double-stranded DNA virus, ranges from 250 to 380 nm in length and 80 to 120 nm in diameter and is classified in the genus Whispovirus of the family Nimaviridae [2]. Within 7–10 days, the mortality rate of shrimp can reach 100% following WSSV infection [3,4]. Despite considerable research into treatment approaches and various methods to prevent WSD in recent years, the development of effective control strategies and treatments remains an ongoing challenge [5]. Therefore, understanding the resistance mechanisms of L. vannamei against the WSSV is crucial.
Invertebrates lack an acquired immune system and rely mainly on the innate immune system to resist foreign bodies and pathogens [6]. Following bacterial or viral infection, host pattern recognition receptors (PRRs) recognize pathogen-associated molecular patterns (PAMPs), which trigger the initiation of innate immune defense [7,8]. Various PRRs have been identified in invertebrates, including toll-like receptors (TLRs), scavenger receptors (SRs), gram-negative binding proteins (GNBPs), and C-type lectins (CTLs). PRRs elicit various immune responses. Therefore, functional research on PRRs has attracted widespread attention [9].
CTLs are important PRRs and are widespread in both vertebrates and invertebrates. CTLs typically contain one or more carbohydrate recognition domains (CRDs). The CRD includes 110–130 amino acids and has a conserved structural motif arranged in two protein loops stabilized by two disulfide bridges [10]. CTLs specifically bind to glycoproteins on the surface of pathogens via CRDs, which triggers a series of immune responses [11,12]. CTLs mediate intercellular interactions, regulate cytokine expression, promote phagocytosis and encapsulation, activate inflammatory responses, and play important roles in cell apoptosis [13,14,15].
CTLs play important roles in the invertebrate immune processes. Stomach virus-associated CTLs capture viral protein post-WSSV infection and present it to receptors on the surfaces of related shrimp host cells, facilitating WSSV infection in shrimp [16]. When recombinant CTL (rLvLec) was injected into L. vannamei infected with Vibrio, LvLec regulated the immune response of hemocytes via the cGMP-PKA signaling pathway [17]. LvCTL5 from L. vannamei was found to have a direct bacteriostatic and immunoregulatory function [18]. A shrimp CTL, as the PRR of bacteria and the ligand of the dome, mediates the activation of the Jak/STAT pathway [19]. LvCTL 4.2 in L. vannamei inhibited Vibrio but facilitated WSSV infection [20].
This study identified the C-type lectin domain family 4 member F (LvCLEC4F) in L. vannamei. To determine whether LvCLEC4F is involved in the response of L. vannamei to WSSV infection, the expression profile of LvCLEC4F was determined in the hepatopancreas, gill, muscle, and eyestalk of healthy and WSSV-infected L. vannamei. Using RNA interference (RNAi) technology to knockdown the expression of LvCLEC4F, the specific function of LvCLEC4F in the anti-WSSV response was analyzed. Hemolymph agglutination in vivo of L. vannamei, WSSV viral load, L. vannamei survival rate, and the expression levels of genes related to the regulation of apoptosis, signaling and cellular stress response, and immune defense were detected.
2. Materials and Methods
2.1. Experimental Animals
All pathogenic-free L. vannamei were obtained from BLUP Aquabreed Co., Ltd. (Weifang, China). Samples of L. vannamei were randomly selected and subjected to diagnostic screening for seven pathogens, including WSSV, acute hepatopancreatic necrosis disease-causing Vibrio parahaemolyticus (VpAHPND), covert mortality nodavirus (CMNV), infectious hypodermal and hematopoietic necrosis virus (IHHNV), infectious myonecrosis virus (IMNV), Decapod iridescent virus 1 (DIV1), and Enterocytozoon hepatopenaei (EHP). No pathogens were identified. Experiments were conducted at the Aquatic Genetic Breeding Center, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences (Qingdao, China). Before beginning animal experiments, L. vannamei were acclimatized in sterile seawater (salinity 29‰, temperature 26 ± 1°C) for three days. L. vannamei were subjected to the same daily management, including changing water once a day (each time 1/2 vol), supplying sufficient dissolved oxygen, and providing a consistent diet of commercial feed. L. vannamei with 2.5–3.5 g body weight and 6.0–6.6 cm body length were used to clone the LvCLEC4F sequence and analyze the LvCLEC4F expression profiles post-WSSV infection. L. vannamei with 2.7–3.3 g body weight and 5.5–6.3 cm body length were used for the RNAi experiment.
2.2. RNA Extraction and cDNA Synthesis
Total RNA was extracted using the RNA-Easy isolation reagent (Vazyme Biotech Co., Ltd., Nanjing, China) following the manufacturer’s instructions. The quality and concentration of RNA were assessed by 1% agarose gel electrophoresis using a NanoPhotometer® spectrophotometer (IMPLEN, Munchen, Germany). For the real-time PCR (RT-PCR) assay, cDNA was synthesized from total RNA using a HiScript® III RT SuperMix for qPCR (+gDNA wiper) kit (Vazyme Biotech Co., Ltd., Nanjing, China) following the manufacturer’s instructions.
2.3. Molecular Cloning and Sequencing of LvCLEC4F
Based on the reference sequence of LvCLEC4F in the NCBI database (XM_027356524.1), primer3 (https://primer3.ut.ee/, accessed on 26 March 2023) was used to design the primers (LvCLEC4F-F and LvCLEC4F-R) (Table 1) for cDNA sequence amplification of LvCLEC4F. The cDNA solution was acquired from the hepatopancreas in L. vannamei. The ORF sequence of LvCLEC4F was amplified using Quick Taq HS DyeMix (TOYOBO, Shanghai, China). PCR was performed in a 50 μL volume containing 4 μL cDNA solution (100 ng/μL), 25 μL 2x Quick Taq HS DyeMix, and 1 μL of each primer (10 μM). The PCR reaction was performed as follows: 94 °C for 2 min; 94 °C for 30 s, 55 °C for 30 s, and 68 °C for 1 min for 35 cycles; and it was extended at 68 °C for 5 min. PCR products were sequenced by Sangon Biotech Co., Ltd. (Shanghai, China).

Table 1.
The sequences of primers and probes used in this study.
2.4. Bioinformatics Analysis
The ORF sequence of LvCLEC4F was translated into an amino acid sequence using EditSeq 7.1 software (https://www.dnastar.com/, accessed on 18 April 2023). Protein molecular weight (MW) and the theoretical isoelectric point (PI) instability coefficient were predicted using the ExPASy-ProtParam tool (https://web.expasy.org/protparam/, accessed on 18 April 2023). The transmembrane region of the protein was analyzed using TMHMM 2.0 software (http://www.cbs.dtu.dk/services/TMHMM/, accessed on 18 April 2023). SignalP 4.1 software (https://services.healthtech.dtu.dk/services/SignalP-4.1/, accessed on 20 April 2023) was used to predict the signal peptide. Glycosylation sites were analyzed using NetNGlyc 1.0 online software (https://services.healthtech.dtu.dk/services/NetNGlyc-1.0/, accessed on 21 April 2023). The phosphorylation site analysis used NetPhos 3.1 software (https://services.healthtech.dtu.dk/services/NetPhos-3.1/, accessed on 21 April 2023). The SMART online tool (https://smart.embl.de/, accessed on 21 April 2023) was used for protein functional domain prediction analysis. The secondary structure of the protein was predicted using SOPMA software (https://npsa-prabi.ibcp.fr/, accessed on 21 April 2023). Alignment with homologous sequences from other species was performed using DNAMAN 6.0 (https://www.lynnon.com/, accessed on 28 April 2023). The phylogenetic tree for LvCLEC4F was constructed using the neighbor-joining (NJ) method of MEGA 7.0 (https://www.megasoftware.net/, accessed on 28 April 2023).
2.5. WSSV Challenge Experiment
The experiment was divided into three groups, with fifty shrimp per group. To detect the distribution of LvCLEC4F, the hepatopancreas, gill, muscle, and eyestalk of healthy L. vannamei were collected. Three shrimp from every group were taken. The remaining L. vannamei were individually orally infected with the WSSV bait. The viral load of the WSSV bait was 1 × 107 copies. The hepatopancreas, gill, muscle, and eyestalk of L. vannamei at 24, 48, 72, 96, 144, 192, and 228 h post-WSSV infection were collected and stored in RNAstore reagent (Tiangen Biotech Co., Ltd., Beijing, China) at −80 °C. Three individual shrimp from each group were taken at every sampling time. The collected samples were used for total RNA extraction and cDNA synthesis to assess the expression level of LvCLEC4F.
2.6. RT-PCR
The expression level of LvCLEC4F was detected using an RT-PCR assay with SYBR Green Real-time PCR Master Mix (TOYOBO, Shanghai, China) and the Applied BiosystemsTM QuantStudio 1 Real-Time PCR quantifier (Applied Biosystems, Foster City, CA, USA) with primers (qLvCLEC4F-F, qLvCLEC4F-R, 18S-F, and 18S-R) (Table 1). 18S ribosomal RNA (18S rRNA) was used as an internal control. RT-PCR was performed in a 20 μL volume containing 10 μL SYBR Green Realtime PCR Master Mix, 0.8 μL of each primer (10 μM), and 2 μL cDNA solution (100 ng/μL). RT-PCR was performed as follows: 95 °C for 60 s; 95 °C for 15 s, 60 °C for 15 s, and 72 °C for 45 s for 40 cycles. All experiments were performed in triplicate, and the data were analyzed using the 2–ΔΔCt method. An unpaired two-tailed t-test was used to compare the results.
2.7. Synthesis of Double-Stranded RNA
Double-stranded RNAs (dsRNAs) targeting LvCLEC4F (designated dsLvCLEC4F) were synthesized by in vitro transcription. dsRNAs targeting GFP (designated dsGFP) were also synthesized and used as negative controls. The DNA template for dsLvCLEC4F preparation was generated by PCR using the primers dsLvCLEC4F-Fi and dsLvCLEC4F-Ri (Table 1), with cDNA from the hepatopancreas of L. vannamei as a template. The DNA template for dsGFP preparation was generated by PCR using the primers dsGFP-Fi and dsGFP-Ri (Table 1), with the plasmid pET28a containing the GFP sequence as a template. The PCR products contained the T7 promoter sequence. According to the manufacturer’s instructions, dsRNAs were synthesized using a T7 in vitro transcription kit (Takara, Dalian, China). The in vitro transcription was performed in a 20 μL volume, which contained 2 μL 10 × transcription buffer, 2 μL ATP solution, 2 μL GTP solution, 2 μL CTP solution, 2 μL UTP solution, 0.5 μL Rnase inhibitor, 2 μL T7 RNA polymerase, and 3 μL DNA templates (1 μg). The reaction mixture was incubated at 42 °C for 2 h and then was incubated at 37 °C for 30 min after being added to 2 μL of Rnase-free Dnase I. The dsRNAs were stored at −80 °C.
2.8. Knockdown of LvCLEC4F by dsRNA
To assess the RNAi-mediated knockdown efficiency of the dsRNAs, 60 L. vannamei were randomly divided into two groups: dsGFP+WSSV and dsLvCLEC4F+WSSV. L. vannamei was injected at the third abdominal segment with dsGFP (3 μg/g shrimp) (dsGFP+WSSV group) and dsLvCLEC4F (3 μg/g shrimp) (dsLvCLEC4F+WSSV group), respectively. A total of 24 h after the first injection, the same dose of dsRNAs was injected again to enhance RNAi efficiency, followed by the injection of WSSV virus suspension (4.7 × 106 copies) in a 20 μL volume for the dsGFP+WSSV and dsLvCLEC4F+WSSV groups. The hepatopancreas of L. vannamei in the two groups were collected at 36 and 48 h post-WSSV injection. The expression level of LvCLEC4F in the hepatopancreas was determined using RT-PCR with three replicates.
2.9. Survival Rate Analysis after LvCLEC4F Knockdown
The healthy L. vannamei were randomly divided into four groups (thirty individual shrimp per group), named the dsLvCLEC4F+WSSV group, dsGFP+WSSV group (as control), PBS group (as control), and WSSV group (as control). L. vannamei was injected at the third abdominal segment with dsLvCLEC4F (3 μg/g shrimp) (dsLvCLEC4F+WSSV group), dsGFP (3 μg/g shrimp) (dsGFP+WSSV group), or 1 × PBS (PBS group), respectively. A total of 24 h after the first injection, the same dose of dsRNAs was injected again, followed by the injection of WSSV virus suspension (4.7 × 106 copies) in a 20 μL volume for the dsLvCLEC4F+WSSV, dsGFP+WSSV, and WSSV groups. The survival rates of L. vannamei in four groups were determined at different time points (0, 12, 24, 36, 48, 72, and 96 h) after WSSV injection. The survival rates between different groups of L. vannamei were tested for statistical significance using the log-rank test. Statistical analyses were performed using GraphPad Prism software (version 8.0; https://www.graphpad.com/, accessed on 12 September 2023).
2.10. Investigating Immune Response and Gene Expression after LvCLEC4F Knockdown
2.10.1. Experimental Setting
Hemolymph agglutination in vivo, the viral load of WSSV, and the gene expression related to the innate immunity of L. vannamei were assessed after LvCLEC4F knockdown. L. vannamei were randomly divided into four groups (thirty individual shrimp per group), named the dsLvCLEC4F+WSSV group, dsGFP+WSSV group, PBS group, and WSSV group. L. vannamei was injected at the third abdominal segment with dsLvCLEC4F (3 μg/g shrimp) (dsLvCLEC4F+WSSV group), dsGFP (3 μg/g shrimp) (dsGFP+WSSV group), or 1 × PBS (PBS group), respectively. A total of 24 h after the first injection, the same dose of dsRNAs was injected again, followed by the injection of WSSV virus suspension (4.7 × 106 copies) in a 20 μL volume for the dsLvCLEC4F+WSSV, dsGFP+WSSV, and WSSV group. The hepatopancreas and muscle of L. vannamei were collected at 0, 36, 48, and 72 h post-WSSV infection and stored at −80 °C. The hemolymph of L. vannamei was collected at 48 h post-WSSV infection. All experiments were performed in triplicate.
2.10.2. Detection of Hemolymph Agglutination
The hemolymph agglutination phenomenon in vivo at 48 h post-WSSV infection was observed using an optical microscope. All experiments were performed in triplicate.
2.10.3. The WSSV Viral Load Analysis with TaqMan RT-PCR
DNA was extracted from the muscle of L. vannamei using a TIANamp Marine Animals DNA Kit (Tiangen Biotech Co., Ltd., Beijing, China). The quality and concentration of DNA were assessed by 1% agarose gel electrophoresis and using a NanoPhotometer® spectrophotometer (IMPLEN, Munchen, Germany). The copy number of WSSV in the muscle of L. vannamei was detected using TaqMan RT-PCR with an Applied BiosystemsTM QuantStudio 1 Real-Time PCR Quantifier (Applied Biosystems, Foster City, CA, USA) and the THUNDERBIRDTM Probe qPCR Mix kit (TOYOBO, Shanghai, China) with primers WSSV-F and WSSV-R and the WSSV probe (Table 1) [21]. TaqMan RT-PCR was performed in a 20 μL volume containing 10 μL THUNDERBIRD Probe qPCR Mix, 0.6 μL of each primer (10 μM), 0.4 μL WSSV probe (10 μM), 0.1 μL ROX reference dye, and 2 μL DNA template. TaqMan RT-PCR was performed as follows: 95 °C for 30 s, 40 cycles of 95 °C for 5 s, and 60 °C for 34 s. All experiments were performed in triplicate. The WSSV viral load in L. vannamei was measured by quantifying the concentration of WSSV DNA.
2.10.4. Gene Expression Analysis
To further confirm the role of the LvCLEC4F in anti-WSSV innate immunity, RT-PCR was used to examine the expression levels of genes related to innate immunity, including B cell leukemia (Bcl-2) (XM_027353493.1), caspase 3 (KC660103.1), caspase 8 (XM_027383230.1), mitogen-activated protein kinase p38b-like (p38MAPK) (XM_027367740.1), and lysozyme (Lyz) (AY170126) in the above-mentioned RNAi assay. Total RNA was extracted from the hepatopancreas of L. vannamei infected with WSSV at different time points (0, 36, 48, and 72 h) in the dsGFP+WSSV and dsLvCLEC4F+WSSV groups. The RNA extraction, cDNA synthesis, and RT-PCR methods are as described above. Gene expression was detected with primers (Bcl-2-F, Bcl-2-R, caspase 3-F, caspase 3-R, caspase 8-F, caspase 8-R, p38MAPK-F, p38MAPK-R, Lyz-F, Lyz-R, 18S-F, and 18S-R) (Table 1) with three replicates.
3. Results
3.1. Characterization of LvCLEC4F
The complete ORF sequence of LvCLEC4F was 492 bp, encoding 163 amino acids (Figure 1). The MW was 18.70 kDa, the theoretical PI was 4.88, and the instability index was 40.53, which was classified as an unstable protein. There were 20 negatively and 11 positively charged amino acid residues. The aliphatic index was 65.21, and the average hydrophilicity was -0.273. This protein was predicted to contain eight serine (Ser), three threonine (Thr), and four tyrosine (Tyr) phosphorylation sites. A signal peptide consisting of 16 amino acids was present at the N-terminus. The CRD was located at positions 30–162 (Figure 1). Among the predicted secondary structures of LvCLEC4F after the removal of the signal peptide, the α-helix (26 amino acids) accounted for 17.69%, the β-strand (40 amino acids) accounted for 27.21%, the beta-turn (12 amino acids) accounted for 8.16%, and the random coil (69 amino acids) accounted for 46.94%.
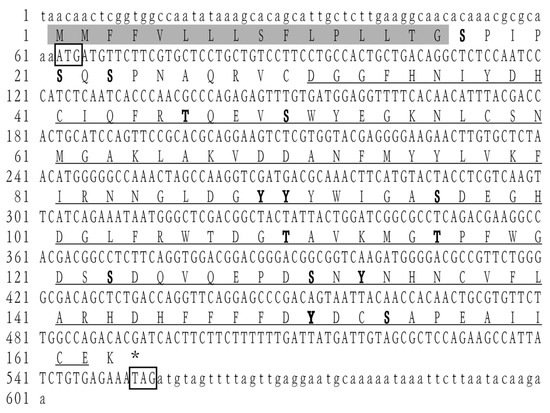
Figure 1.
cDNA and amino acid sequence of LvCLEC4F. The start and stop codons are indicated by boxes. The predicted carbohydrate recognition domain (CRD) is indicated by an underline. The predicted phosphorylation sites are indicated in bold font. The predicted signal peptide structure is indicated by a gray shadow.
3.2. Multiple Alignments and Phylogenetic Analysis
To further investigate the amino acid sequence similarity between LvCLEC4F and CLEC4F in other species, several vertebrates and invertebrates were selected for multiple comparisons of CLEC4F. Sequence conservation of the CLEC4F protein is relatively low, with significant differences in amino acid sequences among different species. The homology between LvCLEC4F and CLEC4F of the American lobster Homarus americanus was the highest, with a similarity of 49.12%. This was followed by some vertebrates (40.30–21.93%) and invertebrates (23.73–20.31%) (Figure 2).
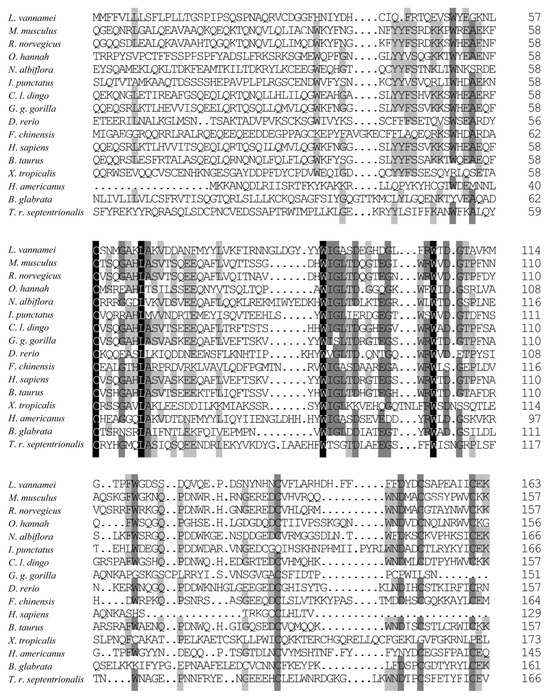
Figure 2.
The amino acid sequence alignment of CLEC4F. One hundred percent identical residues are indicated by a black shadow. Seventy-five percent identical residues are indicated by a dark gray shadow. Fifty percent identical residues are indicated by a light gray shadow. The GenBank accession numbers of CLEC4F are as follows: L. vannamei (XP_027212325.1), Rattus norvegicus (NP_446205.1), Canis lupus dingo (XP_025326743.1), Gorilla gorilla gorilla (XP_055234888.1), Danio rerio (XP_009299422.1), Fenneropenaeus chinensis (XP_047498904.1), Homo sapiens (KAI4034935.1), Bos taurus (XP_027410747.1), Xenopus tropicalis (XP_031750858.1), Homarus americanus (XP_042233235.1), Toxorhynchites rutilus septentrionalis (XP_055620181.1), Mus musculus (NP_058031.2), Ophiophagus hannah (ETE57245.1), Nibea albiflora (KAG8004890.1), Ictalurus punctatus (NP_001187725.1), Biomphalaria glabrata (XP_055887399.1).
A phylogenetic evolutionary analysis was conducted to study the evolutionary relationship between LvCLEC4F and the selected vertebrates and invertebrates further. The phylogenetic tree was consistent with the taxonomic status of the species. L. vannamei clustered preferentially with H. americanus (Figure 3).
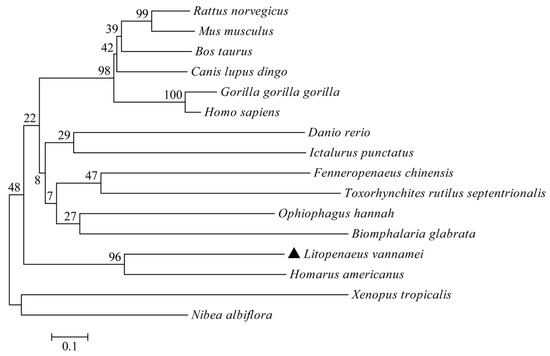
Figure 3.
Phylogenetic tree analysis of CLEC4F. The GenBank accession numbers of CLEC4F sequences are identical to those listed in Figure 2. The LvCLEC4F marker of L. vannamei is ▲.
3.3. The Expression Profiles of LvCLEC4F Post-WSSV Infection
RT-PCR demonstrated that LvCLEC4F was expressed in the hepatopancreas, gill, muscle, and eyestalk of healthy L. vannamei. The expression level of LvCLEC4F in the hepatopancreas was significantly higher than in the gill, muscle, and eyestalk. The expression level of LvCLEC4F in the gill was approximately 0.021-fold (p < 0.05) the level in the hepatopancreas. The expression level of LvCLEC4F in the muscle was approximately 0.005-fold (p < 0.05) the level in the hepatopancreas. The expression level of LvCLEC4F in the eyestalk was approximately 0.014-fold (p < 0.05) the level in the hepatopancreas (Figure 4).
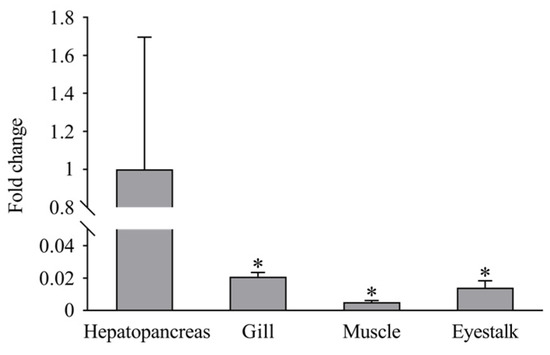
Figure 4.
Expression profiles of LvCLEC4F in the hepatopancreas, gill, muscle, and eyestalk of healthy L. vannamei. *: p < 0.05.
Post-WSSV infection, the expression levels of LvCLEC4F in the hepatopancreas of L. vannamei at 24, 48, 72, 96, 144, 192, and 228 h were approximately 1.78-fold, 11.32-fold (p < 0.01), 15.29-fold (p < 0.05), 45.19-fold, 17.82-fold (p < 0.01), 147.69-fold (p < 0.05), and 39.58-fold (p < 0.01) the level in the control, respectively (Figure 5A). The expression levels of LvCLEC4F in the gill at 24, 48, 72, 96, 144, 192, and 228 h were approximately 0.75-fold (p < 0.05), 0.93-fold, 1.82-fold (p < 0.05), 0.52-fold (p < 0.01), 0.45-fold (p < 0.01), 0.35-fold (p < 0.01), and 0.94-fold the level in the control, respectively (Figure 5B). The expression levels of LvCLEC4F in the muscle at 24, 48, 72, 96, 144, 192, and 228 h were approximately 0.92-fold, 1.21-fold, 0.99-fold, 2.04-fold (p < 0.01), 1.49-fold, 1.32-fold (p < 0.05), and 2.51-fold the level in the control, respectively (Figure 5C). The expression levels of LvCLEC4F in the eyestalk at 24, 48, 72, 96, 144, 192, and 228 h were approximately 0.74-fold, 0.63-fold, 0.57-fold (p < 0.05), 0.85-fold, 0.43-fold (p < 0.01), 0.53-fold (p < 0.05), and 1.67-fold the level in the control, respectively (Figure 5D).
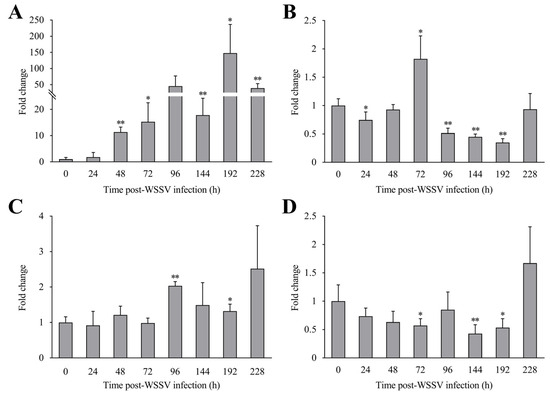
Figure 5.
Expression profiles of LvCLEC4F in the hepatopancreas, gill, muscle, and eyestalk of L. vannamei post-WSSV infection. (A) The expression level of LvCLEC4F in the hepatopancreas at different time points post-WSSV infection. (B) The expression level of LvCLEC4F in the gill at different time points post-WSSV infection. (C) The expression level of LvCLEC4F in the muscle at different time points post-WSSV infection. (D) The expression level of LvCLEC4F in the eyestalk at different time points post-WSSV infection. *: p < 0.05, **: p < 0.01.
3.4. WSSV Infection Was Suppressed after Knocking down LvCLEC4F
An RT-PCR assay was conducted to confirm RNAi efficiency. The results showed that the LvCLEC4F expression level in the dsLvCLEC4F+WSSV group was reduced by approximately 73.94% and 60.18% at 36 and 48 h post-WSSV infection, respectively, compared with that in the dsGFP+WSSV group (Figure 6A), indicating the successful in vivo knockdown of LvCLEC4F.
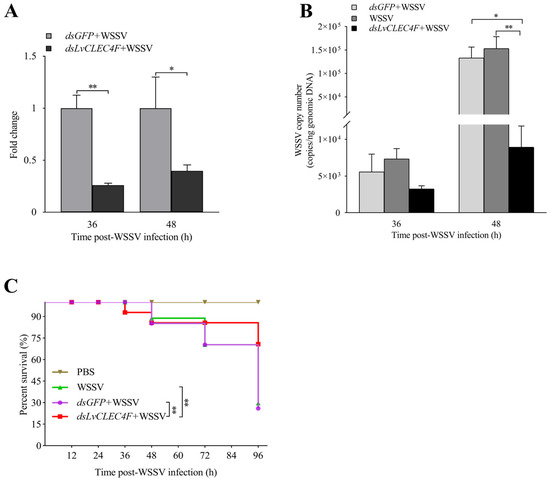
Figure 6.
WSSV replication was suppressed after knocking down LvCLEC4F. (A) The knockdown efficiency of LvCLEC4F in the hepatopancreas at 36 and 48 h post-WSSV infection. (B) The WSSV viral load in the dsGFP+WSSV group (as control), the WSSV group (as control), and the dsLvCLEC4F+WSSV group after knocking down LvCLEC4F. (C) The survival curve of L. vannamei post-WSSV infection. *: p < 0.05, **: p < 0.01.
A TaqMan RT-PCR assay was performed to investigate the characteristics of WSSV replication in the muscle of L. vannamei after knocking down LvCLEC4F. The results showed that the WSSV copy number in L. vannamei in the dsGFP+WSSV group was 5.57 × 103 copies/ng at 36 h post-WSSV infection. The WSSV copy number in L. vannamei in the WSSV group was 7.34 × 103 copies/ng at 36 h post-WSSV infection. The WSSV copy number in L. vannamei in the dsLvCLEC4F+WSSV group was 3.25 × 103 copies/ng at 36 h post-WSSV infection (Figure 6B). The WSSV copy number in L. vannamei in the dsGFP+WSSV group was 1.33 × 105 copies/ng at 48 h post-WSSV infection. The WSSV copy number in L. vannamei in the WSSV group was 1.53 × 105 copies/ng at 48 h post-WSSV infection. The WSSV copy number in L. vannamei in the dsLvCLEC4F+WSSV group was 8.93 × 103 copies/ng at 48 h post-WSSV infection. The WSSV copy number in L. vannamei in the dsLvCLEC4F+WSSV group was significantly lower (0.067-fold) (p < 0.05) than that in the dsGFP+WSSV group and significantly lower (0.058-fold) (p < 0.01) than that in the WSSV group at 48 h post-WSSV infection (Figure 6B). Subsequently, the survival rates of L. vannamei at 0, 12, 24, 36, 48, 72, and 96 h post-WSSV infection were analyzed. The survival rate of L. vannamei in the PBS group was 100%. There was no significant difference in the survival rate of L. vannamei between the dsGFP+WSSV and WSSV groups. The survival rate of L. vannamei in the dsLvCLEC4F+WSSV group was significantly higher than that in the dsGFP+WSSV and WSSV groups (p < 0.01) (Figure 6C).
3.5. Hemolymph Agglutination In Vivo Assay
Optical microscopy was performed to investigate the effect of LvCLEC4F knockdown on hemolymph agglutination in L. vannamei. The results showed that there was no apparent agglutination in the PBS group (Figure 7A,E). The hemolymph agglutination phenomenon was weakened in the dsLvCLEC4F+WSSV group (Figure 7D,H) compared with the WSSV (Figure 7B,F) and dsGFP+WSSV (Figure 7C,G) groups after knocking down LvCLEC4F at 48 h post-WSSV infection.
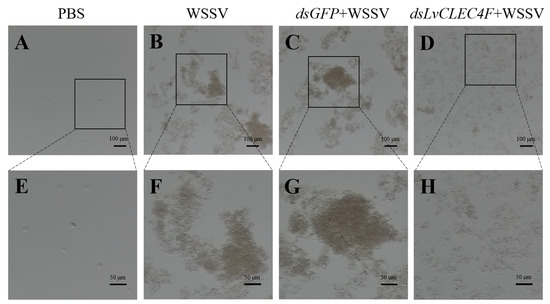
Figure 7.
The observation of hemolymph agglutination in L. vannamei following WSSV infection after knocking down LvCLEC4F. (A) PBS group (scale bar = 100 μm). (B) WSSV group (scale bar = 100 μm). (C) dsGFP+WSSV group (scale bar = 100 μm). (D) dsLvCLEC4F+WSSV group (scale bar = 100 μm). (E) PBS group (scale bar = 50 μm). (F) WSSV group (scale bar = 50 μm). (G) dsGFP+WSSV group (scale bar = 50 μm). (H) dsLvCLEC4F+WSSV group (scale bar = 50 μm).
3.6. The Expression Profiles of Genes Related to Innate Immunity after Knocking down LvCLEC4F
RT-PCR was used to detect the expression levels of genes related to innate immunity in L. vannamei after knocking down LvCLEC4F. The expression levels of Bcl-2 in the dsLvCLEC4F+WSSV group at 36, 48, and 72 h were approximately 0.63-fold, 0.30-fold (p < 0.01), and 0.28-fold (p < 0.05), respectively, compared to those in the dsGFP+WSSV group (Figure 8A). The expression levels of caspase 3 in the dsLvCLEC4F+WSSV group at 36, 48, and 72 h were approximately 1.01-fold, 0.37-fold (p < 0.05), and 0.32-fold (p < 0.01), respectively, compared to those in the dsGFP+WSSV group (Figure 8B). The expression levels of caspase 8 in the dsLvCLEC4F+WSSV group at 36, 48, and 72 h were approximately 0.46-fold, 0.26-fold (p < 0.05), and 0.18-fold (p < 0.01), respectively, compared to those in the dsGFP+WSSV group (Figure 8C). The expression levels of p38MAPK in the dsLvCLEC4F+WSSV group at 36, 48, and 72 h were approximately 1.09-fold, 0.28-fold (p < 0.01), and 0.30-fold (p < 0.01), respectively, compared to those in the dsGFP+WSSV group (Figure 8D). The expression levels of Lyz in the dsLvCLEC4F+WSSV group at 36, 48, and 72 h were approximately 0.08-fold (p < 0.05), 0.23-fold (p < 0.01), and 1.00-fold, respectively, compared to those in the dsGFP+WSSV group (Figure 8E).
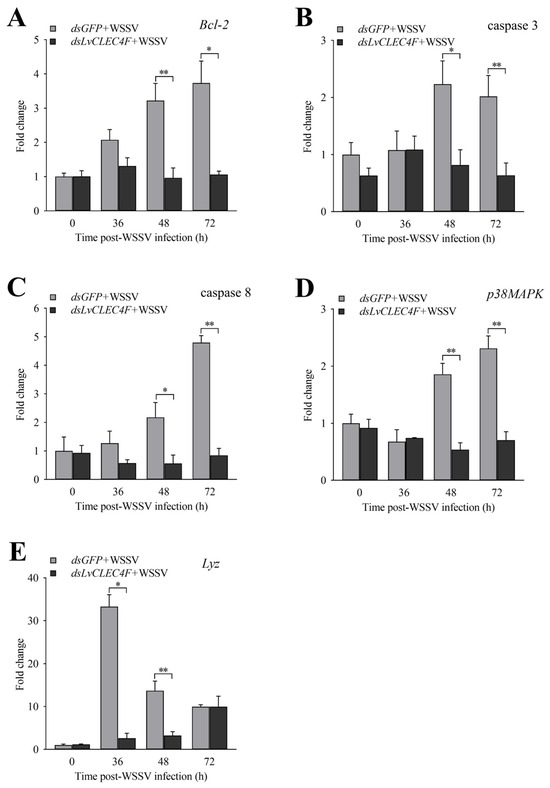
Figure 8.
Expression profiles of immune-related genes after knocking down LvCLEC4F. (A) The expression level of Bcl-2 at different time points post-WSSV infection after knocking down LvCLEC4F. (B) The expression level of caspase 3 at different time points post-WSSV infection after knocking down LvCLEC4F. (C) The expression level of caspase 8 at different time points post-WSSV infection after knocking down LvCLEC4F. (D) The expression level of p38MAPK at different time points post-WSSV infection after knocking down LvCLEC4F. (E) The expression level of Lyz at different time points post-WSSV infection after knocking down LvCLEC4F. *: p < 0.05, **: p < 0.01.
4. Discussion
CTLs serve as crucial PRRs and immune modulators and play a significant role in innate immune responses [22]. Studies have confirmed that CTLs exhibit pathogen-binding and agglutinating activities. Three CTLs (MjLecA, MjLecB, and MjLecC) of M. japonicus bind to viral envelope proteins, preventing WSSV infection of hemocytes [23]. The CTLs (PcLec-1 and PcLec-2) of crayfish participate in immune responses against bacterial and viral infections [24,25]. A shrimp CTL displays a bacterial agglutination ability [26]. The present study cloned LvCLEC4F from L. vannamei. LvCLEC4F contains a 492 bp ORF encoding 163 amino acids, comprising a signal peptide structure and CRD (Figure 1). The CRD of CTLs can bind to carbohydrate molecules on the pathogen surface, triggering immune responses [27]. LvCTL 4.2 in L. vannamei was a C-type mannose-binding lectin with a CRD containing a mutated mannose-binding motif that could facilitate WSSV pathogenesis [20]. A crayfish CTL contains CRD that can bind to Vibrio and WSSV [22].
The present study found that LvCLEC4F is expressed in the hepatopancreas, gill, muscle, and eyestalk of healthy L. vannamei. The expression level of LvCLEC4F was the highest in the hepatopancreas compared to all other organs and tissues examined (Figure 4). A previous report stated that LvCTL7 expression levels in L. vannamei were higher in the hepatopancreas, muscle, gill, and eyestalk, but lower in the brain, epidermis, thoracic ganglion, intestine, heart, and hemocytes [6]. In the current study, the expression level of LvCLEC4F in the hepatopancreas was greatly influenced by WSSV infection, showing significant upregulation (Figure 5A). This was consistent with the upregulation of hepatopancreas CTL of a related shrimp following bacterial and WSSV challenges [28].
Knocking down the expression of LvCLEC4F in L. vannamei via RNAi resulted in a significant decrease in the WSSV viral load (Figure 6B) and a significant increase in the survival rate of L. vannamei (Figure 6C), indicating that LvCLEC4F might promote WSSV infection. Studies have reported that soluble CTLs capture and present virions to cell surface receptors to facilitate viral infection. The replication of WSSV was inhibited after the knockdown of a CTL expression of a related shrimp [16]. Moreover, the WSSV viral load in L. vannamei significantly decreased after knocking down PvDnaJC16 [29]. Like the lectin family, the toll receptor family is a group of conserved PRRs that primarily control the initiation of innate immune responses. Silencing Toll2 significantly increases the survival rate of shrimp post-WSSV infection and reduces the viral load, suggesting that Toll2 can promote WSSV infection in shrimp [30]. Based on the above results, it was speculated that LvCLEC4F could promote the replication of WSSV and affect the survival rate of L. vannamei.
To investigate the reason for the improved immunity of L. vannamei after knocking down LvCLEC4F, the hemolymph agglutination in L. vannamei following WSSV infection after knocking down LvCLEC4F was observed. The expression levels of genes related to the regulation of apoptosis, signaling and cellular stress response, and immune defense were analyzed. The results indicated an apparent trend of weakened hemolymph aggregation compared to that in the dsGFP+WSSV group after knocking down LvCLEC4F (Figure 7). LvLdlrCTL exhibits agglutination activity against bacteria and fungi and potentiates the phagocytosis of hemocytes [31]. Relevant signaling pathways, including the apoptotic pathway, and other immune pathways, can exert immune functions post-WSSV infection [32,33,34]. Experimental studies have shown that apoptosis induced by WSSV infection constitutes a pivotal host defense response against viral infections [35]. In the present study, the expression of genes in the apoptotic pathway, primarily Bcl-2 (Figure 8A), caspase 3 (Figure 8B), and caspase 8 (Figure 8C), showed a significant downward trend after knocking down LvCLEC4F. Additionally, a significant decrease in WSSV viral load was detected after knocking down LvCLEC4F. The findings were similar to a previous study, which demonstrated that intramuscular injection of rLvHSP70 in L. vannamei effectively extended the survival rate of WSSV-infected shrimp and led to a decreased WSSV viral load. Furthermore, a significant reduction in the expression of apoptosis-related genes was observed [36].
The expression of gene p38MAPK (Figure 8D) showed a significant downward trend after knocking down LvCLEC4F. p38MAPK was a vital gene induced in response to WSSV infection in a shrimp [34]. In this study, the expression of Lyz showed a significant downward trend following the knockdown of LvCLEC4F (Figure 8E). It was consistent with a previous report that after knocking down MrLec, the expression of lysozyme 2 was downregulated in a freshwater shrimp post-Vibrio challenge [37]. AMPs play crucial roles in innate immune defense responses [38]. Previous studies have also shown that knocking down LvLdlrCTL leads to the upregulation or downregulation of many immune effector genes in shrimp post-WSSV infection [31].
5. Conclusions
In conclusion, a LvCLEC4F has been cloned from L. vannamei. The expression profiles of LvCLEC4F in the hepatopancreas, gill, muscle, and eyestalk of L. vannamei were significantly altered post-WSSV infection. After knocking down LvCLEC4F, the survival rate of L. vannamei significantly increased, and the WSSV viral load significantly decreased. In addition, the hemolymph agglutination phenomenon was weakened. Finally, the expression of genes related to the regulation of apoptosis, signaling and cellular stress response, and immune defense (Bcl-2, caspase 3, caspase 8, p38MAPK, and Lyz) were significantly downregulated. Overall, LvCLEC4F was an important receptor gene that could promote WSSV replication and affect the survival rate of L. vannamei.
Author Contributions
Data curation, investigation, validation, and writing—original draft: Q.X., B.Y. and X.L.; resources: K.L., S.L., X.M. and J.K.; formal analysis and writing—review and editing: X.L. and X.M.; conceptualization and funding acquisition: X.L., S.L. and X.M.; project administration: X.L., S.L., X.M., J.K. and K.L.; supervision: J.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the National Key R&D Program of China (2022YFF1000304), the National Natural Science Foundation of China (32172960), the China Agriculture Research System of MOF and MARA (CARS-48), the Central Public-interest Scientific Institution Basal Research Fund, CAFS (2020TD26), and the Open Competition Program of Top Ten Critical Priorities of Agricultural Science and Technology Innovation for the 14th Five-Year Plan of Guangdong Province (2022SDZG01).
Institutional Review Board Statement
Under approval number YSFRI-2024005, this study was conducted with the approval of the committee at the Yellow Sea Fisheries Research Institute.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author/s.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hou, C.; Song, W.; Yuan, H.; Hu, N.; Tan, B.; Zhang, S. Comparative transcriptome analysis revealed that dietary zymosan-A improved the immunity of Penaeus vannamei by regulating the TLR signaling pathway. Aquaculture 2022, 561, 738603. [Google Scholar] [CrossRef]
- Lightner, D.V. Global transboundary disease politics: The OIE perspective. J. Invertebr. Pathol. 2012, 110, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Lightner, D.V. Virus diseases of farmed shrimp in the Western Hemisphere (the Americas): A review. J. Invertebr. Pathol. 2011, 106, 110–130. [Google Scholar] [CrossRef]
- Chen, J.; Li, Z.; Hew, C. Characterization of a novel envelope protein WSV010 of shrimp white spot syndrome virus and its interaction with a major viral structural protein VP24. Virology 2007, 364, 208–213. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Verbruggen, B.; Bickley, L.; van Aerle, R.; Bateman, K.S.; Stentiford, G.D.; Santos, E.M.; Tyler, C.R. Molecular Mechanisms of White Spot Syndrome Virus Infection and Perspectives on Treatments. Viruses 2016, 8, 23. [Google Scholar] [CrossRef]
- Luo, J.; Chen, Y.; Huang, Y.; Feng, J.; Yuan, Y.; Jian, J.; Cai, S.; Yang, S. A novel C-type lectin for Litopenaeus vannamei involved in the innate immune response against Vibrio infection. Fish Shellfish Immunol. 2023, 135, 108621. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J.A.; Kafatos, F.C.; Janeway, C.A., Jr.; Ezekowitz, R.A.B. Phylogenetic Perspectives in Innate Immunity. Science 1999, 284, 1313–1318. [Google Scholar] [CrossRef] [PubMed]
- De Gregorio, E.; Spellman, P.T.; Rubin, G.M.; Lemaitr, B. Genome-wide analysis of the Drosophila immune response by using oligonucleotide microarrays. Proc. Natl. Acad. Sci. USA 2001, 98, 12590–12595. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, J. Pattern recognition receptors acting in innate immune system of shrimp against pathogen infections. Fish Shellfish Immunol. 2013, 34, 981–989. [Google Scholar] [CrossRef]
- Sancho, D.; Reis e Sousa, C. Signaling by Myeloid C-type Lectin Receptors in Immunity and Homeostasis. Annu. Rev. Immunol. 2012, 30, 491–529. [Google Scholar] [CrossRef]
- Drickamer, K. Evolution of Ca2+-dependent Animal Lectins. Prog. Nucl. Res. Molec. Biol. 1993, 45, 207–232. [Google Scholar]
- Drickamer, K. Two Distinct Classes of Carbohydrate-recognition Domains in Animal Lectins. J. Biol. Chem. 1988, 263, 9557–9560. [Google Scholar] [CrossRef] [PubMed]
- Rogers, N.C.; Slack, E.C.; Edwards, A.D.; Nolte, M.A.; Schulz, O.; Schweighoffer, E.; Williams, D.L.; Gordon, S.; Tybulewicz, V.L.; Brown, G.D.; et al. Syk-Dependent Cytokine Induction by Dectin-1 Reveals a Novel Pattern Recognition Pathway for C Type Lectins. Immunity 2005, 22, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.C.; Azumi, K.; Nonaka, M. Complement systems in invertebrates. The ancient alternative and lectin pathways. Immunopharmacology 1999, 42, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Van Vliet, S.J.; García-Vallejo, J.J.; van Kooy, Y. Dendritic cells and C-type lectin receptors: Coupling innate to adaptive immune responses. Immunol. Cell Biol. 2008, 86, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, Y.; Xu, J.; Zhao, X.; Wang, J. Collaboration between a soluble C-type lectin and calreticulin facilitates white spot syndrome virus infection in shrimp. J. Immunol. 2014, 193, 2106–2117. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Pan, L.; Yu, J. The injection of one recombinant C-type lectin (LvLec) induced the immune response of hemocytes in Litopenaeus vannamei. Fish Shellfish Immunol. 2022, 124, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Yang, L.; Wang, Z.; Zuo, H.; Weng, S.; He, J.; Xu, X. A novel C-type lectin with microbiostatic and immune regulatory functions from Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 93, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Lan, J.; Zhao, X.; Vasta, G.R.; Wang, J. Binding of a C-type lectin’s coiled-coil domain to the Domeless receptor directly activates the JAK/STAT pathway in the shrimp immune response to bacterial infection. PLoS Pathog. 2017, 13, e1006626. [Google Scholar] [CrossRef]
- Huang, Y.H.; Kumar, R.; Liu, C.H.; Lin, S.S.; Wang, H.C. A novel C-type lectin LvCTL 4.2 has antibacterial activity but facilitates WSSV infection in shrimp (L. vannamei). Dev. Comp. Immunol. 2022, 126, 104239. [Google Scholar] [CrossRef]
- Durand, S.V.; Lightner, D.V. Quantitative real time PCR for the measurement of white spot syndrome virus in shrimp. J. Fish Dis. 2002, 25, 381–389. [Google Scholar] [CrossRef]
- Chen, D.; Meng, X.; Xu, J.; Yu, J.; Meng, M.; Wang, J. PcLT, a novel C-type lectin from Procambarus clarkii, is involved in the innate defense against Vibrio alginolyticus and WSSV. Dev. Comp. Immunol. 2013, 39, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Li, D.; Zhang, M.; Yang, H.; Ruan, L.; Xu, X. Cloning and characterization of three novel WSSV recognizing lectins from shrimp Marsupenaeus japonicus. Fish Shellfish Immunol. 2010, 28, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, H.; Li, X.; Zhao, X.; Wang, J. Characterization of a C-type lectin (PcLec2) as an upstream detector in the prophenoloxidase activating system of red swamp crayfish. Fish Shellfish Immunol. 2011, 30, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, X.; Sun, C.; Zhao, X.; Wang, J. C-type lectin from red swamp crayfish Procambarus clarkii participates in cellular immune response. Arch. Insect Biochem. Physiol. 2011, 76, 168–184. [Google Scholar] [CrossRef]
- Alenton, R.R.R.; Koiwai, K.; Nakamura, R.; Thawonsuwan, J.; Kondo, H.; Hirono, I. A Hint of Primitive Mucosal Immunity in Shrimp through Marsupenaeus japonicus Gill C-Type Lectin. J. Immunol. 2019, 203, 2310–2318. [Google Scholar] [CrossRef]
- Mahla, R.S.; Reddy, M.C.; Prasad, D.V.; Kumar, H. Sweeten PAMPs: Role of Sugar Complexed PAMPs in Innate Immunity and Vaccine Biology. Front. Immunol. 2013, 4, 248. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, W.; Wang, X.; Mu, Y.; Zhao, X.; Yu, X.; Wang, J. A novel C-type lectin with two CRD domains from Chinese shrimp Fenneropenaeus chinensis functions as a pattern recognition protein. Mol. Immunol. 2009, 46, 1626–1637. [Google Scholar] [CrossRef]
- Jaree, P.; Somboonwiwat, K. DnaJC16, the molecular chaperone, is implicated in hemocyte apoptosis and facilitates of WSSV infection in shrimp. Fish Shellfish Immunol. 2023, 137, 108770. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, H.; Zhu, J.; Liu, H.; Yang, Y.; Sun, B.; Wu, T.; Zhang, Y.; Yao, D. The transcription factor CEBP homolog of Penaeus vannamei contributes to WSSV replication. Fish Shellfish Immunol. 2023, 134, 108571. [Google Scholar] [CrossRef]
- Liang, Z.; Yang, L.; Zheng, J.; Zuo, H.; Weng, S.; He, J.; Xu, X. A low-density lipoprotein receptor (LDLR) class A domain-containing C-type lectin from Litopenaeus vannamei plays opposite roles in antibacterial and antiviral responses. Dev. Comp. Immunol. 2019, 92, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Dostert, C.; Jouanguy, E.; Irving, P.; Troxler, L.; Galiana-Arnoux, D.; Hetru, C.; Hoffmann, J.A.; Imler, J.L. The Jak-STAT signaling pathway is required but not sufficient for the antiviral response of drosophila. Nat. Immunol. 2005, 6, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Huang, Y.; Wei, S.; Huang, X.; Ye, F.; Fu, J.; Qin, Q. Characterization of p38 MAPKs from orange-spotted grouper, Epinephelus coioides involved in SGIV infection. Fish Shellfish Immunol. 2011, 31, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yao, W.; Liu, P.; Li, J.; Wang, Q. Expression profiles of the p38 MAPK signaling pathway from Chinese shrimp Fenneropenaeus chinensis in response to viral and bacterial infections. Gene 2018, 642, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Leu, J.H.; Lin, S.J.; Huang, J.Y.; Chen, T.C.; Lo, C.F. A model for apoptotic interaction between white spot syndrome virus and shrimp. Fish Shellfish Immunol. 2013, 34, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Janewanthanakul, S.; Supungul, P.; Tang, S.; Tassanakajon, A. Heat shock protein 70 from Litopenaeus vannamei (LvHSP70) is involved in the innate immune response against white spot syndrome virus (WSSV) infection. Dev. Comp. Immunol. 2020, 102, 103476. [Google Scholar] [CrossRef]
- Huang, X.; Li, W.; Jin, M.; Ma, F.T.; Huang, Y.; Shi, Y.R.; Zhao, L.L.; Feng, J.L.; Ren, Q.; Wang, W. Single CRD containing lectin from Macrobrachium rosenbergii (MrLec) participates in innate immunity against pathogen infections. Fish Shellfish Immunol. 2016, 51, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.L.; Li, W.; Meng, L.; Sha, Z.X.; Wang, Z.J.; Ren, G.C. Molecular cloning and expression analysis of a hepcidin antimicrobial peptide gene from turbot (Scophthalmus maximus). Fish Shellfish Immunol. 2007, 22, 172–181. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).