
Dietary Artemisia arborescens Supplementation Effects on Growth, Oxidative Status, and Immunity of Gilthead Seabream (Sparus aurata L.)

 , , ,
, , ,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Diets
2.3. Animals and Experimental Design
2.4. Growth-Performance Parameters
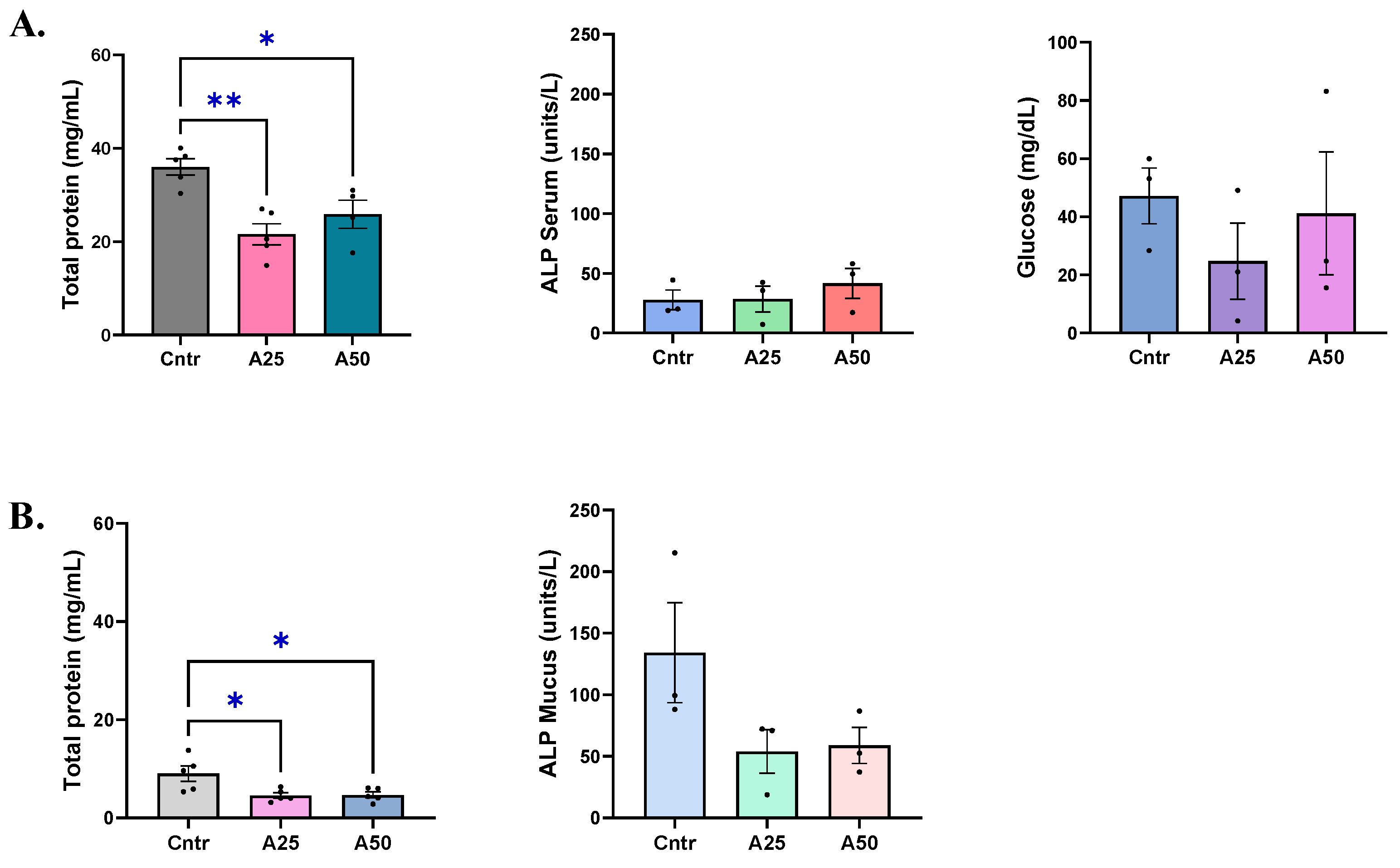
2.5. Biochemical Parameters
2.5.1. Total Protein Levels
2.5.2. Alkaline Phosphatase Levels
2.5.3. Glucose Levels
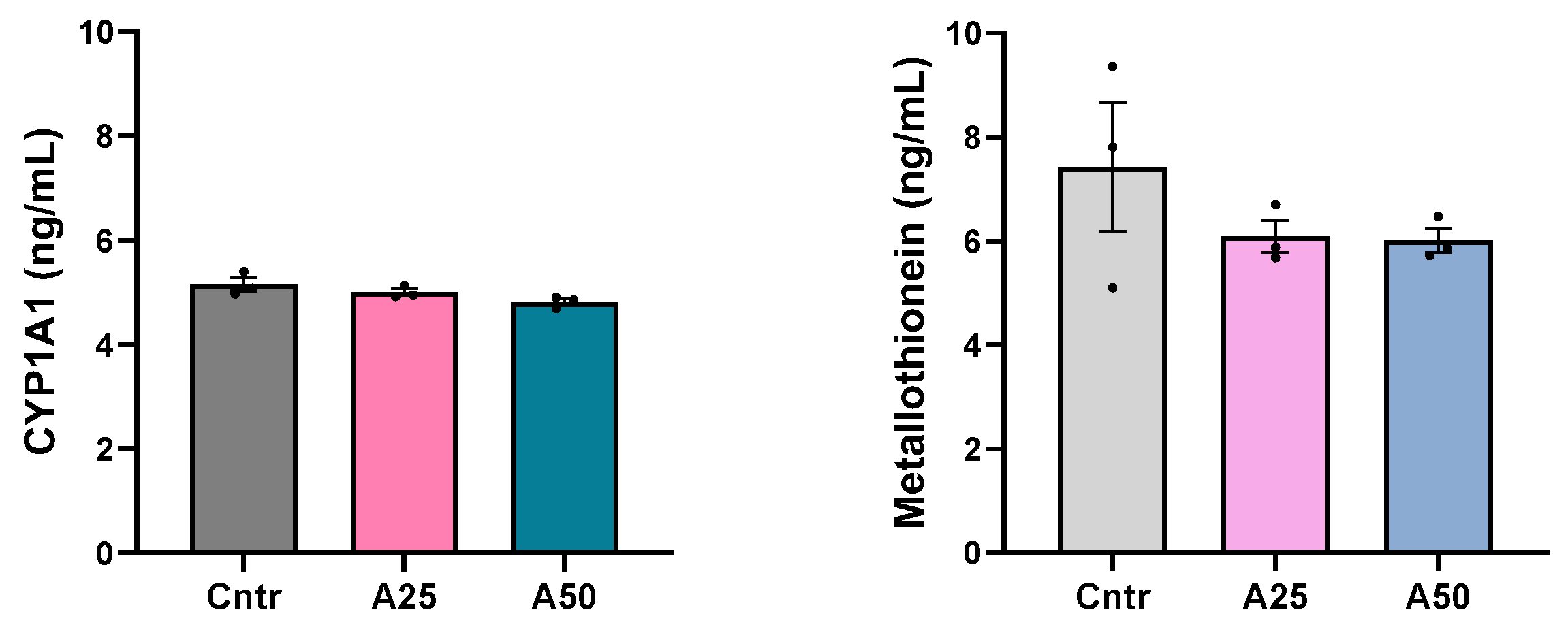
2.6. Oxidative Stress Parameters
2.6.1. Cytochrome P450 1A1 Levels
2.6.2. Metallothionein Levels
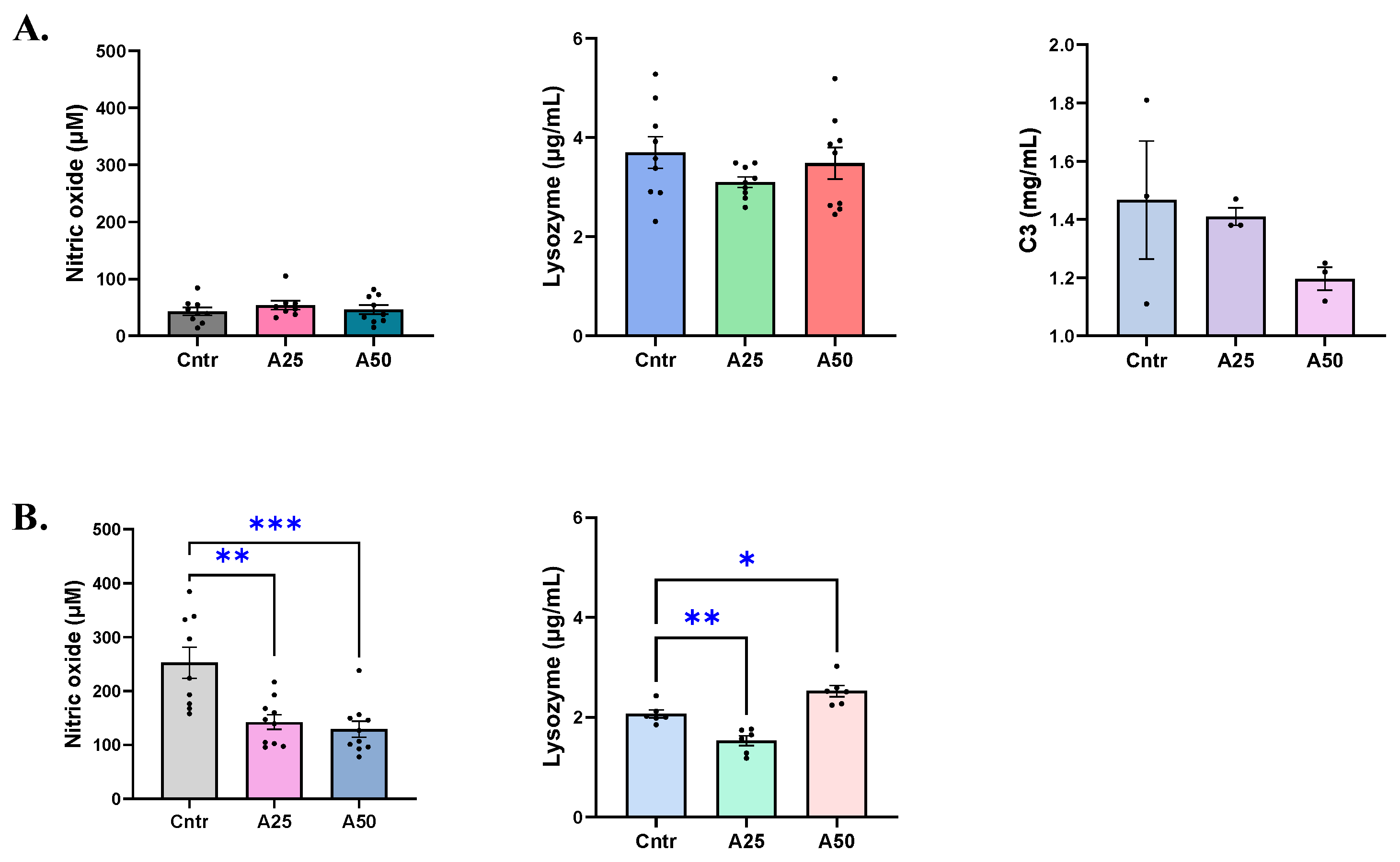
2.7. Immunological Parameters
2.7.1. Nitric Oxide Levels
2.7.2. Lysozyme Activity
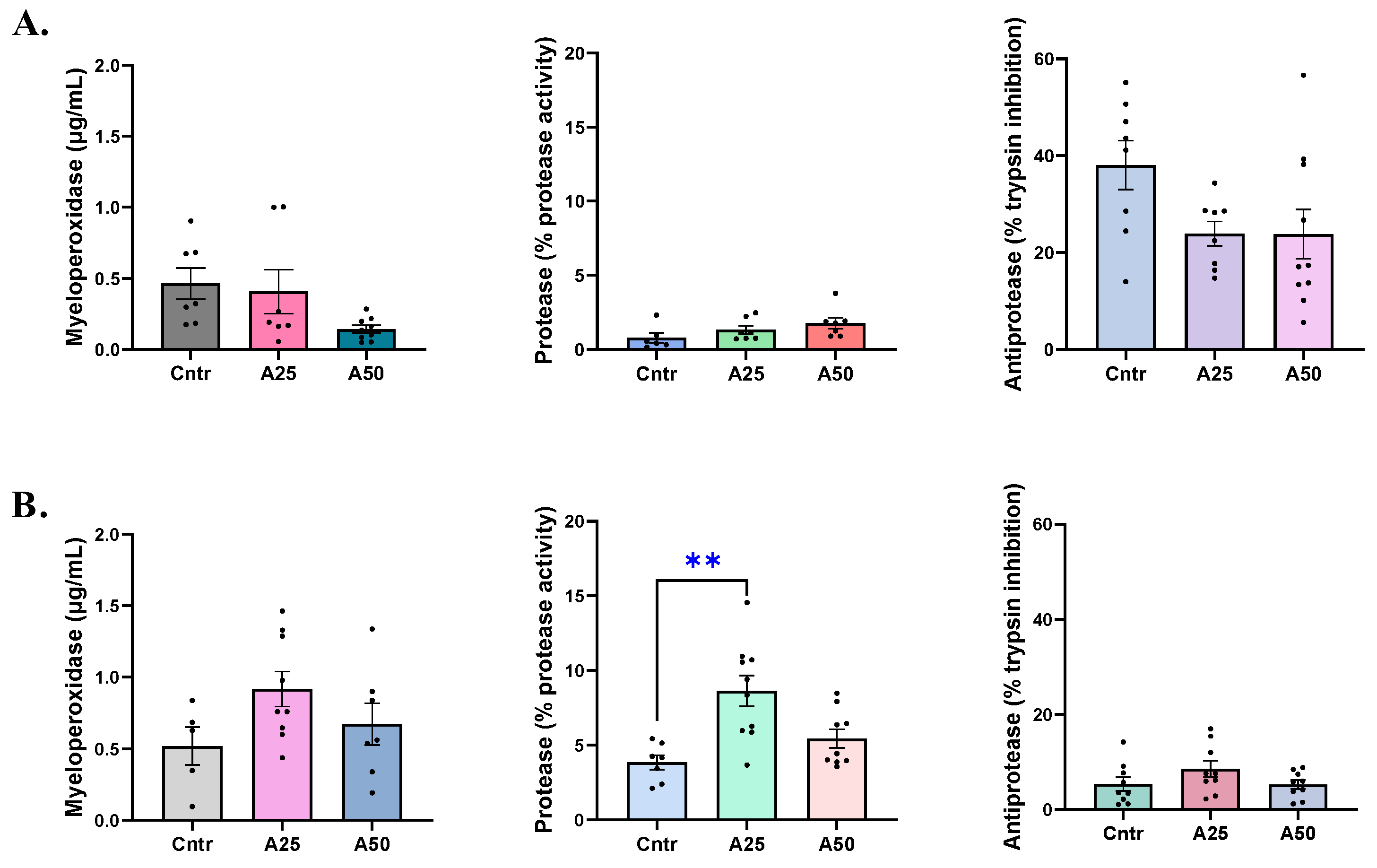
2.7.3. Myeloperoxidase Activity
2.7.4. Protease Activity
2.7.5. Anti-protease Activity
2.7.6. Complement C3 Levels
2.7.7. Total Immunoglobulins and Immunoglobulin M (IgM) Levels
2.8. RNA Isolation and Complementary DNA (cDNA) Synthesis
2.9. Gene-Expression Analysis
2.10. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Biochemical and Oxidative Stress Responses
3.3. Non-Specific Immune Responses
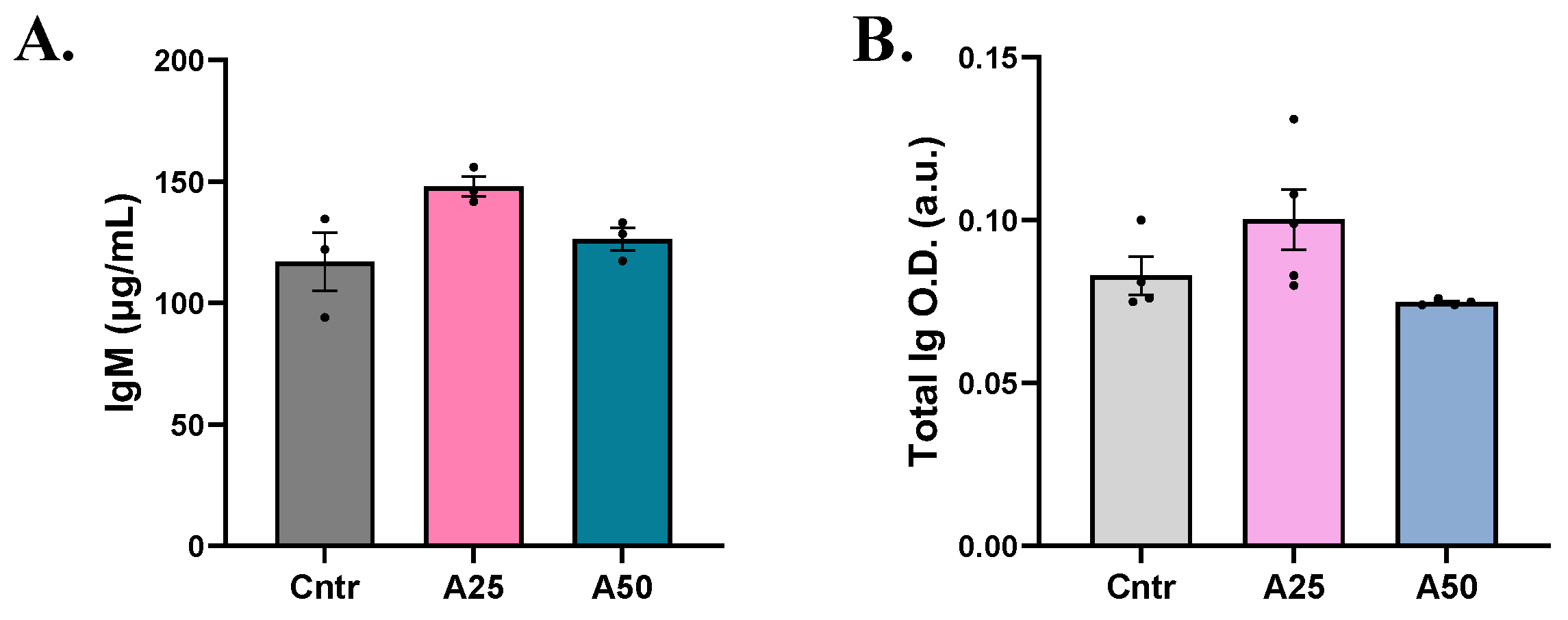
3.4. Immunoglobulin Response
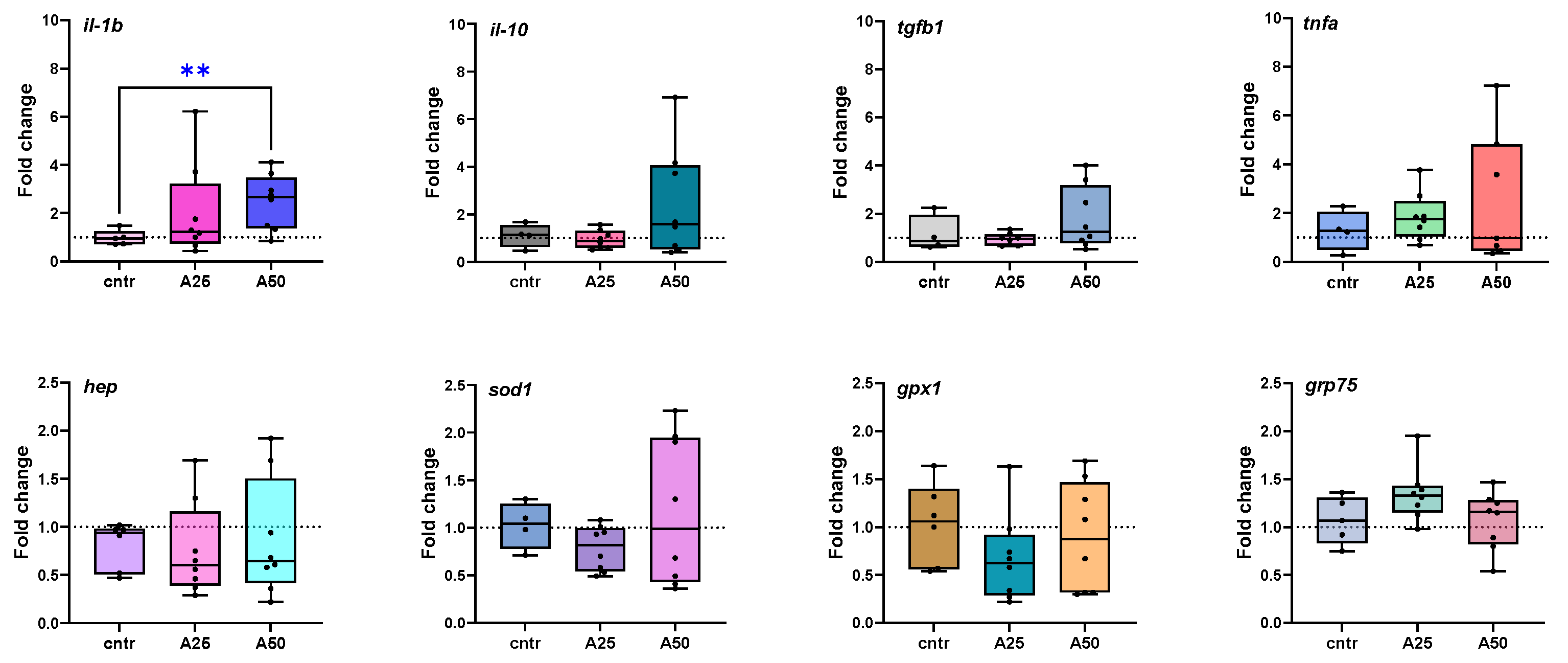
3.5. Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. Food and Agriculture Organization of the United Nations, Blue Transformation. Available online: https://www.fao.org/fishery/en/bluetransformation (accessed on 15 December 2023).
- Stentiford, G.D.; Bateman, I.J.; Hinchliffe, S.J.; Bass, D.; Hartnell, R.; Santos, E.M.; Devlin, M.J.; Feist, S.W.; Taylor, N.G.H.; Verner-Jeffreys, D.W.; et al. Sustainable aquaculture through the One Health lens. Nat. Food 2020, 1, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Koshio, S.; Esteban, M.Á. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquacult. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Reverter, M.; Sarter, S.; Caruso, D.; Avarre, J.C.; Combe, M.; Pepey, E.; Pouyaud, L.; Vega-Heredía, S.; de Verdal, H.; Gozlan, R.E. Aquaculture at the crossroads of global warming and antimicrobial resistance. Nat. Commun. 2020, 11, 1870. [Google Scholar] [CrossRef] [PubMed]
- Reverter, M.; Tapissier-Bontemps, N.; Sarter, S.; Sasal, P.; Caruso, D. Moving towards more sustainable aquaculture practices: A meta-analysis on the potential of plant-enriched diets to improve fish growth, immunity and disease resistance. Rev. Aquacult. 2021, 13, 537–555. [Google Scholar] [CrossRef]
- Aguiar, G.A.C.C.d.; Carneiro, C.L.d.S.; Campelo, D.A.V.; Rusth, R.C.T.; Maciel, J.F.R.; Baldisserotto, B.; Zuanon, J.A.S.; Oliveira, A.V.d.; Oliveira, M.G.d.A.; Freitas, M.B.D.d.; et al. Effects of Dietary Peppermint (Mentha piperita) Essential Oil on Growth Performance, Plasma Biochemistry, Digestive Enzyme Activity, and Oxidative Stress Responses in Juvenile Nile Tilapia (Oreochromis niloticus). Fishes 2023, 8, 374. [Google Scholar] [CrossRef]
- Militello, M.; Settanni, L.; Aleo, A.; Mammina, C.; Moschetti, G.; Giammanco, G.M.; Blàzquez, M.A.; Carrubba, A. Chemical composition and antibacterial potential of Artemisia arborescens L. essential oil. Curr. Microbiol. 2011, 62, 1274–1281. [Google Scholar] [CrossRef]
- Yousefi, M.; Zahedi, S.; Reverter, M.; Adineh, H.; Hoseini, S.M.; Van Doan, H.; El-Haroun, E.R.; Hoseinifarm, S.H. Enhanced growth performance, oxidative capacity and immune responses of common carp, Cyprinus carpio fed with Artemisia absinthium extract-supplemented diet. Aquaculture 2021, 545, 737167. [Google Scholar] [CrossRef]
- Ekiert, H.; Klimek-Szczykutowicz, M.; Rzepiela, A.; Klin, P.; Szopa, A. Artemisia Species with High Biological Values as a Potential Source of Medicinal and Cosmetic Raw Materials. Molecules 2022, 27, 6427. [Google Scholar] [CrossRef]
- Lantzouraki, D.Z.; Amerikanou, C.; Karavoltsos, S.; Kafourou, V.; Sakellari, A.; Tagkouli, D.; Zoumpoulakis, P.; Makris, D.P.; Kalogeropoulos, N.; Kaliora, A.C. Artemisia arborescens and Artemisia inculta from Crete; Secondary Metabolites, Trace Metals and in Vitro Antioxidant Activities. Life 2023, 13, 1416. [Google Scholar] [CrossRef] [PubMed]
- Gholamhosseini, A.; Hosseinzadeh, S.; Soltanian, S.; Banaee, M.; Sureda, A.; Rakhshaninejad, M.; Ali Heidari, A.; Anbazpour, H. Effect of dietary supplements of Artemisia dracunculus extract on the haemato-immunological and biochemical response, and growth performance of the rainbow trout (Oncorhynchus mykiss). Aquac. Res. 2021, 52, 2097–2109. [Google Scholar] [CrossRef]
- Sarhadi, I.; Alizadeh, E.; Ahmadifar, E.; Adineh, H.; Dawood, M.A. Skin mucosal, serum immunity and antioxidant capacity of common carp (Cyprinus carpio) fed artemisia (Artemisia annua). Ann. Anim. Sci. 2020, 20, 1011–1027. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Aydın, B.; Hoseinifar, S.H.; Moonmanee, T.; Van Doan, H. Dietary Artemisia, Artemisia annua, supplementation improves common carp welfare under high stocking density. Aquac. Res. 2022, 53, 3494–3503. [Google Scholar] [CrossRef]
- Soares, M.P.; Cardoso, I.L.; Ishikawa, M.M.; de Oliveira, A.D.S.S.; Sartoratto, A.; Jonsson, C.M.; de Queiroz, S.C.D.N.; Duarte, M.C.T.; Rantin, F.T.; Sampaio, F.G. Effects of Artemisia annua alcohol extract on physiological and innate immunity of Nile tilapia (Oreochromis niloticus) to improve health status. Fish Shellfish Immunol. 2020, 105, 369–377. [Google Scholar] [CrossRef]
- He, G.; Sun, H.; Liao, R.; Wei, Y.; Zhang, T.; Chen, Y.; Lin, S. Effects of herbal extracts (Foeniculum vulgare and Artemisia annua) on growth, liver antioxidant capacity, intestinal morphology and microorganism of juvenile largemouth bass, Micropterus salmoides. Aquac. Rep. 2022, 23, 101081. [Google Scholar] [CrossRef]
- Ji, S.C.; Jeong, G.S.; Im, G.S.; Lee, S.W.; Yoo, J.H.; Takii, K. Dietary medicinal herbs improve growth performance, fatty acid utilization, and stress recovery of Japanese flounder. Fish. Sci. 2007, 73, 70–76. [Google Scholar] [CrossRef]
- Ji, S.C.; Takaoka, O.; Jeong, G.S.; Lee, S.W.; Ishimaru, K.; Seoka, M.; Takii, K. Dietary medicinal herbs improve growth and some non-specific immunity of red sea bream Pagrus major. Fish. Sci. 2007, 73, 63–69. [Google Scholar] [CrossRef]
- Takaoka, O.; Ji, S.C.; Ishimaru, K.; Lee, S.W.; Jeong, G.S.; Biswas, A.; Takii, K. Dietary medicinal herbs and enzyme treated fish meal improve stress resistances and growth performance at early juvenile stage of red sea bream Pagrus major. Aquac. Res. 2016, 47, 390–397. [Google Scholar] [CrossRef]
- Mbokane, E.M.; Moyo, N.A.G. A preliminary investigation into the potential effect of Artemisia afra on growth and disease resistance in sub-adults of Oreochromis mossambicus. Aquaculture 2018, 482, 197–202. [Google Scholar] [CrossRef]
- Mbokane, E.M.; Moyo, N.A.G. Effects of dietary levels of essential oil extracts from Moringa oleifera and Artemisia afra on kidney histology, haemato-immunological parameters and disease resistance in Clarias gariepinus. Aquac. Res. 2020, 51, 410–425. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). The State of Mediterranean and Black Sea Fisheries; General Fisheries Commission for the Mediterranean: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). The State of World Fisheries and Aquaculture 2022: Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Mhalhel, K.; Levanti, M.; Abbate, F.; Laurà, R.; Guerrera, M.C.; Aragona, M.; Porcino, C.; Briglia, M.; Germanà, A.; Montalbano, G. Review on Gilthead Seabream (Sparus aurata) Aquaculture: Life Cycle, Growth, Aquaculture Practices and Challenges. J. Mar. Sci. Eng. 2023, 11, 2008. [Google Scholar] [CrossRef]
- Mladineo, I.; Volpatti, D.; Beraldo, P.; Rigos, G.; Katharios, P.; Padros, F. Monogenean Sparicotyle chrysophrii: The major pathogen of the Mediterranean gilthead seabream aquaculture. Rev. Aquac. 2024, 16, 287–308. [Google Scholar] [CrossRef]
- Cunniff, P.A.; AOAC International. Official Methods of Analysis of AOAC International, 16th ed.; AOAC International: Arlington, TX, USA, 1995. [Google Scholar]
- Wilkinson, P.C. Methods for Studying Mononuclear Phagocytes. Immunology 1983, 49, 569. [Google Scholar]
- Quade, M.J.; Roth, J.A. A rapid, direct assay to measure degranulation of bovine neutrophil primary granules. Vet. Immunol. Immunopathol. 1997, 58, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Ross, N.W.; Firth, K.J.; Wang, A.; Burka, J.F.; Johnson, S.C. Changes in hydrolytic enzyme activities of naïve Atlantic salmon Salmo salar skin mucus due to infection with the salmon louse Lepeophtheirus salmonis and cortisol implantation. Dis. Aquat. Organ. 2000, 41, 43–51. [Google Scholar] [CrossRef]
- Henry, M.; Fountoulaki, E. Optimal dietary protein/lipid ratio for improved immune status of a newly cultivated Mediterranean fish species, the shi drum Umbrina cirrosa. Fish Shellfish Immunol. 2014, 37, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Becerril, M.; Salinas, I.; Cuesta, A.; Meseguer, J.; Tovar-Ramirez, D.; Ascencio-Valle, F.; Esteban, M.A. Oral delivery of live yeast Debaryomyces hansenii modulates the main innate immune parameters and the expression of immune-relevant genes in the gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2008, 25, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Sitjà-Bobadilla, A.; Calduch-Giner, J.; Saera-Vila, A.; Palenzuela, O.; Alvarez-Pellitero, P.; Pérez-Sánchez, J. Chronic exposure to the parasite Enteromyxum leei (Myxozoa: Myxosporea) modulates the immune response and the expression of growth, redox and immune relevant genes in gilthead sea bream, Sparus aurata L. Fish Shellfish Immunol. 2008, 24, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Pozo, E.; Liarte, S.; Fernández-Alacid, L.; Abellán, E.; Meseguer, J.; Mulero, V.; García-Ayala, A. Pattern of expression of immune-relevant genes in the gonad of a teleost, the gilthead seabream (Sparus aurata L.). Mol. Immunol. 2008, 45, 2998–3011. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Guardiola, F.A.; Cuesta, A.; Arizcun, M.; Meseguer, J.; Esteban, M.A. Comparative skin mucus and serum humoral defence mechanisms in the teleost gilthead seabream (Sparus aurata). Fish Shellfish Immunol. 2014, 36, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Lallès, J.P. Biology, environmental and nutritional modulation of skin mucus alkaline phosphatase in fish: A review. Fish Shellfish Immunol. 2019, 89, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Iger, Y.; Abraham, M. The process of skin healing in experimentally wounded carp. J. Fish Biol. 1990, 36, 421–437. [Google Scholar] [CrossRef]
- de Araujo, F.C.T.; Ribeiro, R.P.; Campos, E.C.; Todesco, H.; Tsujii, K.M.; Mantovani, L.S.C.; Ribeiro, R.F.; Carvalho, J.C.; Casetta, J.; Lopera-Barrero, N.M.; et al. Could serum glucose be a selection criterion in Nile tilapia breeding programs? Aquaculture 2022, 548, 737573. [Google Scholar] [CrossRef]
- Eisenman, S.W.; Poulev, A.; Struwe, L.; Raskin, I.; Ribnicky, D.M. Qualitative variation of anti-diabetic compounds in different tarragon (Artemisia dracunculus L.) cytotypes. Fitoterapia 2011, 82, 1062–1074. [Google Scholar] [CrossRef] [PubMed]
- Wadhwa, R.; Taira, K.; Kaul, S.C. An Hsp70 family chaperone, mortalin/mthsp70/PBP74/Grp75: What, when, and where? Cell Stress Chaperones 2002, 7, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Bermejo-Nogales, A.; Benedito-Palos, L.; Saera-Vila, A.; Calduch-Giner, J.A.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J. Confinement exposure induces glucose regulated protein 75 (GRP75/mortalin/mtHsp70/PBP74/HSPA9B) in the hepatic tissue of gilthead sea bream (Sparus aurata L.). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 149, 428–438. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Capó, X.; Alomar, C.; Compa, M.; Sole, M.; Sanahuja, I.; Soliz Rojas, D.L.; González, G.P.; Garcinuño Martínez, R.M.; Deudero, S. Quantification of differential tissue biomarker responses to microplastic ingestion and plasticizer bioaccumulation in aquaculture reared sea bream Sparus aurata. Environ. Res. 2022, 211, 113063. [Google Scholar] [CrossRef]
- Taheri Mirghaed, A.; Hamed Paknejad, H.; Mirzargar, S.S. Hepatoprotective effects of dietary Artemisia (Artemisia annua) leaf extract on common carp (Cyprinus carpio) exposed to ambient ammonia. Aquaculture 2020, 527, 735443. [Google Scholar] [CrossRef]
- El-Houseiny, W.; Anter, R.G.A.; Arisha, A.H.; Mansour, A.T.; Safhi, F.A.; Alwutayd, K.M.; Elshopakey, G.E.; Abd El-Hakim, Y.M.; Mohamed, E.M.M. Growth Retardation, Oxidative Stress, Immunosuppression, and Inflammatory Disturbances Induced by Herbicide Exposure of Catfish, Clarias gariepinus, and the Alleviation Effect of Dietary Wormwood, Artemisia cina. Fishes 2023, 8, 297. [Google Scholar] [CrossRef]
- Dai, Q.Q.; Zhou, X.Q.; Jiang, W.D.; Wu, P.; Liu, Y.; Shi, H.Q.; Zhang, L.; Mi, H.F.; Tang, J.Y.; Zhang, R.N.; et al. Application of enzymatically treated Artemisia annua L. on adult grass carp (Ctenopharyngodon idella): Improved growth performance, intestinal antioxidant capacity and apical junctional complex. Aquaculture 2023, 575, 739612. [Google Scholar] [CrossRef]
- Ortiz-Delgado, J.B.; Segner, H.; Sarasquete, C. Cellular distribution and induction of CYP1A following exposure of gilthead seabream, Sparus aurata, to waterborne and dietary benzo(a)pyrene and 2,3,7,8-tetrachlorodibenzo-p-dioxin: An immunohistochemical approach. Aquat. Toxicol. 2005, 75, 144–161. [Google Scholar] [CrossRef] [PubMed]
- Guinot, D.; Ureña, R.; Pastor, A.; Varó, I.; del Ramo, J.; Torreblanca, A. Long-term effect of temperature on bioaccumulation of dietary metals and metallothionein induction in Sparus aurata. Chemosphere 2012, 87, 1215–1221. [Google Scholar] [CrossRef]
- Tolkou, A.K.; Toubanaki, D.K.; Kyzas, G.Z. Detection of Arsenic, Chromium, Cadmium, Lead, and Mercury in Fish: Effects on the Sustainable and Healthy Development of Aquatic Life and Human Consumers. Sustainability 2023, 15, 16242. [Google Scholar] [CrossRef]
- Župan, I.; Tkalčić, S.; Čož-Rakovac, R.; Strunjak-Perović, I.; Topić-Popović, N.; Babić, S.; Bujak, M.; Šarić, T. Biochemical parameters in the blood of gilthead sea bream (Sparus aurata Linnaeus, 1758) supplemented with commercially available β-glucan-based product (IMUNO-2865®). Aquac. Res. 2018, 49, 786–792. [Google Scholar] [CrossRef]
- Subramanian, S.; MacKinnon, S.L.; Ross, N.W. A comparative study on innate immune parameters in the epidermal mucus of various fish species. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 148, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Holland, M.C.; Lambris, J.D. The complement system in teleosts. Fish Shellfish Immunol. 2002, 12, 399–420. [Google Scholar] [CrossRef]
- Locascio, A.; Annona, G.; Caccavale, F.; D’Aniello, S.; Agnisola, C.; Palumbo, A. Nitric Oxide Function and Nitric Oxide Synthase Evolution in Aquatic Chordates. Int. J. Mol. Sci. 2023, 24, 11182. [Google Scholar] [CrossRef]
- Teixeira, C.; Peixoto, D.; Hinzmann, M.; Santos, P.; Ferreira, I.; Pereira, G.V.; Dias, J.; Costas, B. Dietary Strategies to Modulate the Health Condition and Immune Responses in Gilthead Seabream (Sparus aurata) Juveniles Following Intestinal Inflammation. Animals 2022, 12, 3019. [Google Scholar] [CrossRef] [PubMed]
- Guluarte, C.; Reyes-Becerril, M.; Gonzalez-Silvera, D.; Cuesta, A.; Angulo, C.; Esteban, M.Á. Probiotic properties and fatty acid composition of the yeast Kluyveromyces lactis M3. In vivo immunomodulatory activities in gilthead seabream (Sparus aurata). Fish Shellfish Immunol. 2019, 94, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Ceballos-Francisco, D.; Guardiola, F.A.; Esteban, M.Á. Influence of skin wounds on the intestinal inflammatory response and barrier function: Protective role of dietary Shewanella putrefaciens SpPdp11 administration to gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2020, 99, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Ortuño, J.; Cuesta, A.; Rodríguez, A.; Esteban, M.A.; Meseguer, J. Oral administration of yeast, Saccharomyces cerevisiae, enhances the cellular innate immune response of gilthead seabream (Sparus aurata L.). Vet. Immunol. Immunopathol. 2002, 85, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Rombout, J.H.; Yang, G.; Kiron, V. Adaptive immune responses at mucosal surfaces of teleost fish. Fish Shellfish Immunol. 2014, 40, 634–643. [Google Scholar] [CrossRef]
- Lin, Z.X.; Pan, L.; Xie, R.T.; Li, L.X.; Wen, J.S.; Zhou, X.Q.; Dong, X.H.; Xie, S.W.; Tan, B.P.; Liu, H.Y. Effects of dietary artemisinin on growth performance, digestive enzyme activity, intestinal microbiota, antioxidant capacity and immune biomarkers of Coral trout (Plectropomus leopardus). Aquac. Rep. 2023, 29, 101525. [Google Scholar] [CrossRef]
- Natnan, M.E.; Low, C.F.; Chong, C.M.; Bunawan, H.; Baharum, S.N. Integration of Omics Tools for Understanding the Fish Immune Response Due to Microbial Challenge. Front. Mar. Sci. 2021, 8, 668771. [Google Scholar] [CrossRef]
- Zapata, A.G. The fish spleen. Fish Shellfish Immunol. 2024, 144, 109280. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Xu, H.; Yu, Y.; Xu, Z. Regulatory roles of cytokines in T and B lymphocytes-mediated immunity in teleost fish. Dev. Comp. Immunol. 2023, 144, 104621. [Google Scholar] [CrossRef]
- Zou, J.; Secombes, C.J. The Function of Fish Cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef]
- Toubanaki, D.K.; Efstathiou, A.; Tzortzatos, O.P.; Valsamidis, M.A.; Papaharisis, L.; Bakopoulos, V.; Karagouni, E. Nervous Necrosis Virus Modulation of European Sea Bass (Dicentrarchus labrax, L.) Immune Genes and Transcriptome towards Establishment of Virus Carrier State. Int. J. Mol. Sci. 2023, 24, 16613. [Google Scholar] [CrossRef]
- Cuesta, A.; Meseguer, J.; Esteban, M.A. The antimicrobial peptide hepcidin exerts an important role in the innate immunity against bacteria in the bony fish gilthead seabream. Mol. Immunol. 2008, 45, 2333–2342. [Google Scholar] [CrossRef]
- Shen, Y.; Zhao, Z.; Zhao, J.; Chen, X.; Cao, M.; Wu, M. Expression and Functional Analysis of Hepcidin from Mandarin Fish (Siniperca chuatsi). Int. J. Mol. Sci. 2019, 20, 5602. [Google Scholar] [CrossRef]
- D’Andrea, P.; Giampieri, F.; Battino, M. Nutritional Modulation of Hepcidin in the Treatment of Various Anemic States. Nutrients 2023, 15, 5081. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.R.; Wu, P.; Feng, L.; Jiang, W.D.; Liu, Y.; Kuang, S.Y.; Tang, L.; Zhou, X.Q. Dietary phytic acid weakened the antimicrobial activity and aggravated the inflammatory status of head kidney, spleen and skin in on-growing grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2020, 103, 256–265. [Google Scholar] [CrossRef]
- Costas, B.; Couto, A.; Azeredo, R.; Machado, M.; Krogdahl, A.; Oliva-Teles, A. Gilthead seabream (Sparus aurata) immune responses are modulated after feeding with purified antinutrients. Fish Shellfish Immunol. 2014, 41, 70–79. [Google Scholar] [CrossRef]
- Burrells, C.; Williams, P.D.; Southgate, P.J.; Crampton, V.O. Immunological, physiological and pathological responses of rainbow trout (Oncorhynchus mykiss) to increasing dietary concentrations of soybean proteins. Vet. Immunol. Immunopathol. 1999, 72, 277–288. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Peña-Llopis, S.; Gómez-Requeni, P.; Médale, F.; Kaushik, S.; Pérez-Sánchez, J. Effect of fish meal replacement by plant protein sources on non-specific defence mechanisms and oxidative stress in gilthead sea bream (Sparus aurata). Aquaculture 2005, 249, 387–400. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw Materials | Control Diet (cntr) | Artemisia 0.25% (A25) | Artemisia 0.50% (A50) |
|---|---|---|---|
| Fish meal 67% | 30.00 | 30.00 | 30.00 |
| Soybean meal 45% | 20.00 | 20.00 | 20.00 |
| Wheat flour | 14.38 | 14.38 | 14.38 |
| Wheat gluten meal | 6.00 | 6.00 | 6.00 |
| Fish oil | 13.24 | 12.99 | 12.74 |
| Corn gluten meal | 13.8 | 13.8 | 13.8 |
| Monocalcium phosphate (MCP) | 2.33 | 2.33 | 2.33 |
| Vitamins and minerals premix | 0.25 | 0.25 | 0.25 |
| Artemisia E.O. | 0 | 0.25 | 0.50 |
| Proximate Composition (%) | |||
| Crude Protein * | 44.0 | 44.0 | 44.0 |
| Crude lipid * | 17.7 | 17.7 | 17.7 |
| Carbohydrates ** | 1.8 | 1.8 | 1.8 |
| Moisture * | - | - | - |
| Ash * | 8.7 | 8.7 | 8.7 |
| Starch † | 11.4 | 11.4 | 11.4 |
| Phosphorus † | 1.4 | 1.4 | 1.4 |
| Calcium † | 1.3 | 1.3 | 1.3 |
| Lysine † | 2.4 | 2.4 | 2.4 |
| Methionine + Cysteine † | 1.6 | 1.6 | 1.6 |
| Gross Energy (MJ/kg) *** | 20.8 | 20.8 | 20.8 |
| Parameters | Control (cntr) | 0.25% (A25) | 0.5% (A50) | Significance |
|---|---|---|---|---|
| Mean Initial weight (g) | 108.93 ± 4.21 | 109.94 ± 4.45 | 109.43 ± 2.53 | p1 = 0.53/p2 = 0.84/p3 = 0.84 |
| Mean Final weight (g) | 208.00 ± 11.96 | 207.28 ± 14.31 | 201.91 ± 22.39 | p1 = 0.24/p2 = 2.01/p3 = 1.76 |
| Mean Weight gain (g) | 99.06 | 97.34 | 92.47 | N/A |
| FR (%) | 1.5 | 1.5 | 1.5 | N/A |
| SGR (%/day) | 1.03 ± 0.04 a | 0.95 ± 0.02 b | 0.82 ± 0.02 c | p1 = 0.029/p2 = 0.001/p3 = 0.006 |
| FCR (%) | 1.65 ± 0.03 | 1.82 ± 0.04 | 2.15 ± 0.13 a | p1 = 0.093/p2 = 0.001/p3 = 0.008 |
| Survival rate (%) | 96.7 | 93.9 | 87.88 | p1 = 0.83/p2 = 0.054/p3 = 0.111 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tzortzatos, O.-P.; Toubanaki, D.K.; Kolygas, M.N.; Kotzamanis, Y.; Roussos, E.; Bakopoulos, V.; Chatzopoulos, A.; Athanassopoulou, F.; Karagouni, E. Dietary Artemisia arborescens Supplementation Effects on Growth, Oxidative Status, and Immunity of Gilthead Seabream (Sparus aurata L.). Animals 2024, 14, 1161. https://doi.org/10.3390/ani14081161
Tzortzatos O-P, Toubanaki DK, Kolygas MN, Kotzamanis Y, Roussos E, Bakopoulos V, Chatzopoulos A, Athanassopoulou F, Karagouni E. Dietary Artemisia arborescens Supplementation Effects on Growth, Oxidative Status, and Immunity of Gilthead Seabream (Sparus aurata L.). Animals. 2024; 14(8):1161. https://doi.org/10.3390/ani14081161
Chicago/Turabian StyleTzortzatos, Odysseas-Panagiotis, Dimitra K. Toubanaki, Markos N. Kolygas, Yannis Kotzamanis, Efstratios Roussos, Vasileios Bakopoulos, Achilleas Chatzopoulos, Fotini Athanassopoulou, and Evdokia Karagouni. 2024. "Dietary Artemisia arborescens Supplementation Effects on Growth, Oxidative Status, and Immunity of Gilthead Seabream (Sparus aurata L.)" Animals 14, no. 8: 1161. https://doi.org/10.3390/ani14081161
APA StyleTzortzatos, O.-P., Toubanaki, D. K., Kolygas, M. N., Kotzamanis, Y., Roussos, E., Bakopoulos, V., Chatzopoulos, A., Athanassopoulou, F., & Karagouni, E. (2024). Dietary Artemisia arborescens Supplementation Effects on Growth, Oxidative Status, and Immunity of Gilthead Seabream (Sparus aurata L.). Animals, 14(8), 1161. https://doi.org/10.3390/ani14081161