

Effect of Human Activity and Presence on the Behavior of Long-Tailed Macaques (Macaca fascicularis) in an Urban Tourism Site in Kuala Selangor, Malaysia
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
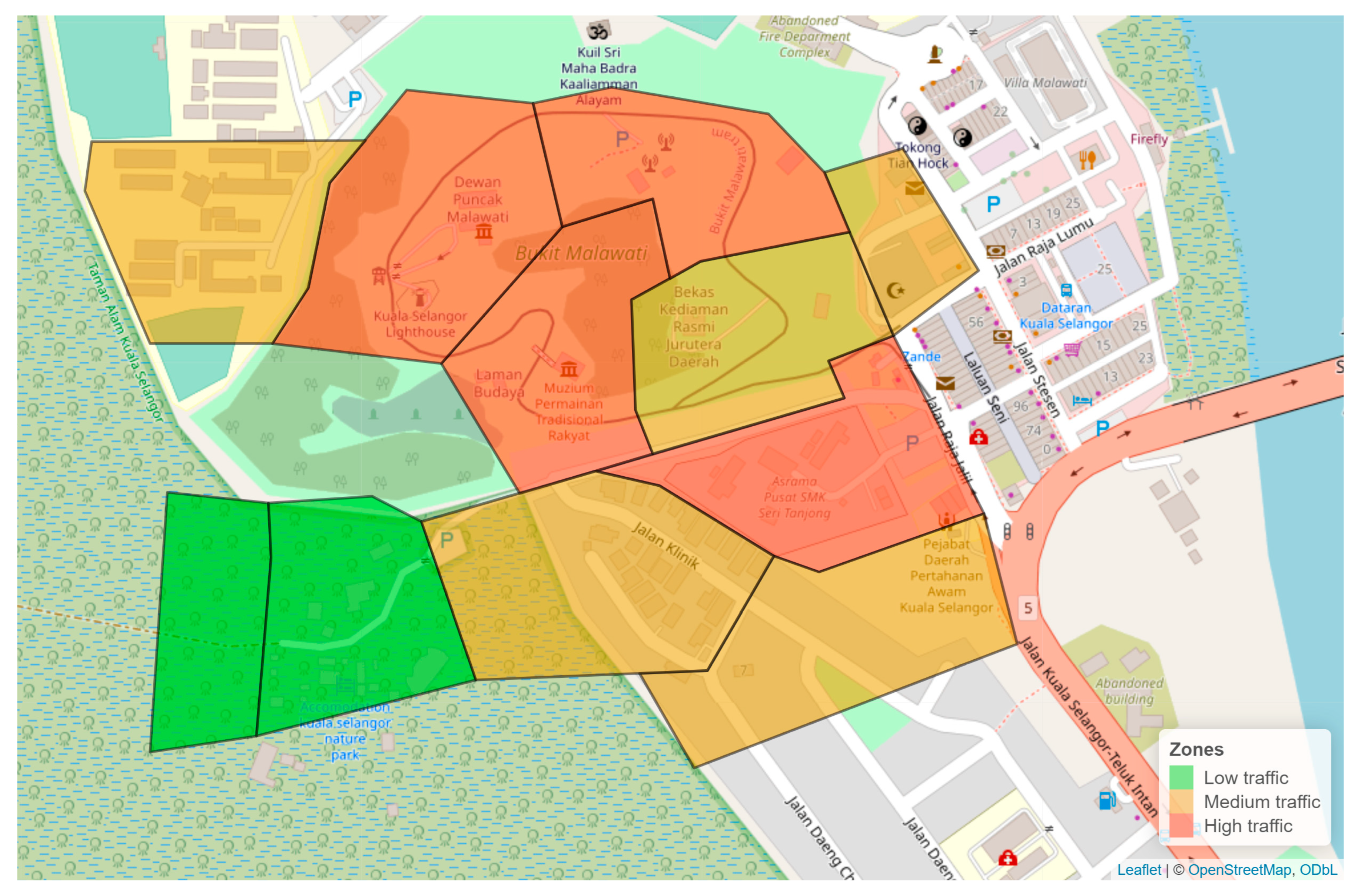
2.1. Study Site and Study Subjects
2.2. Behavior Sampling Method
2.3. Data Analysis
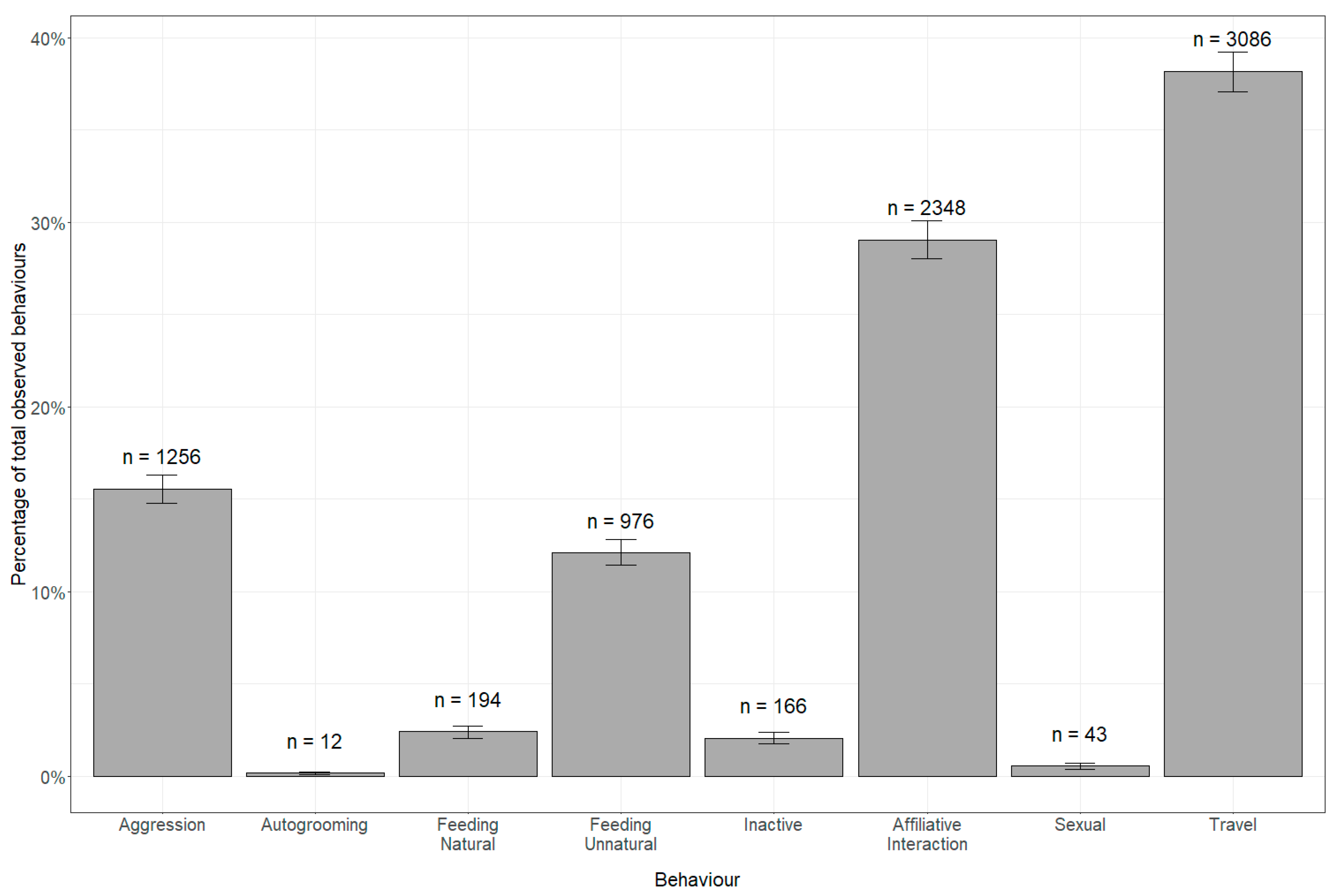
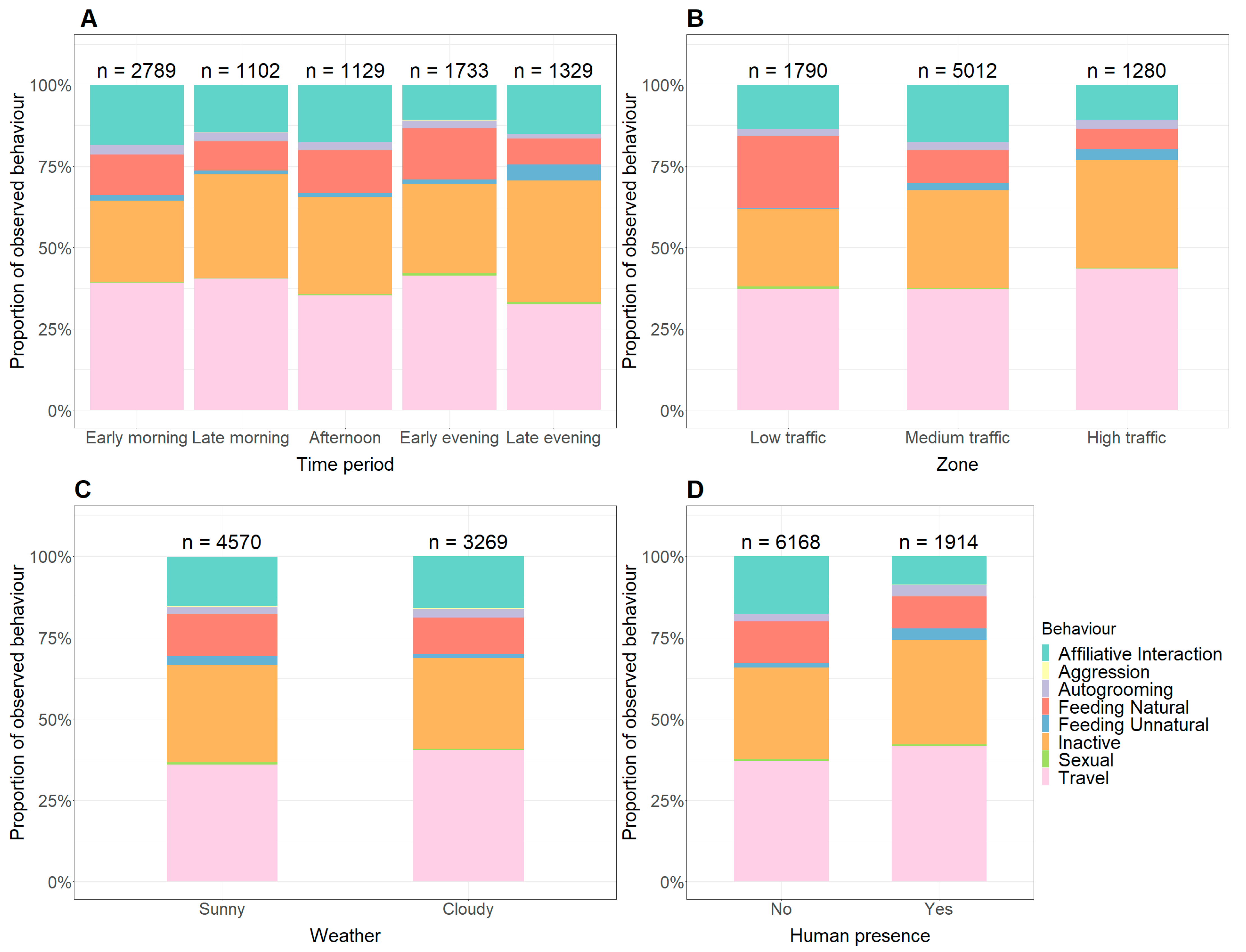
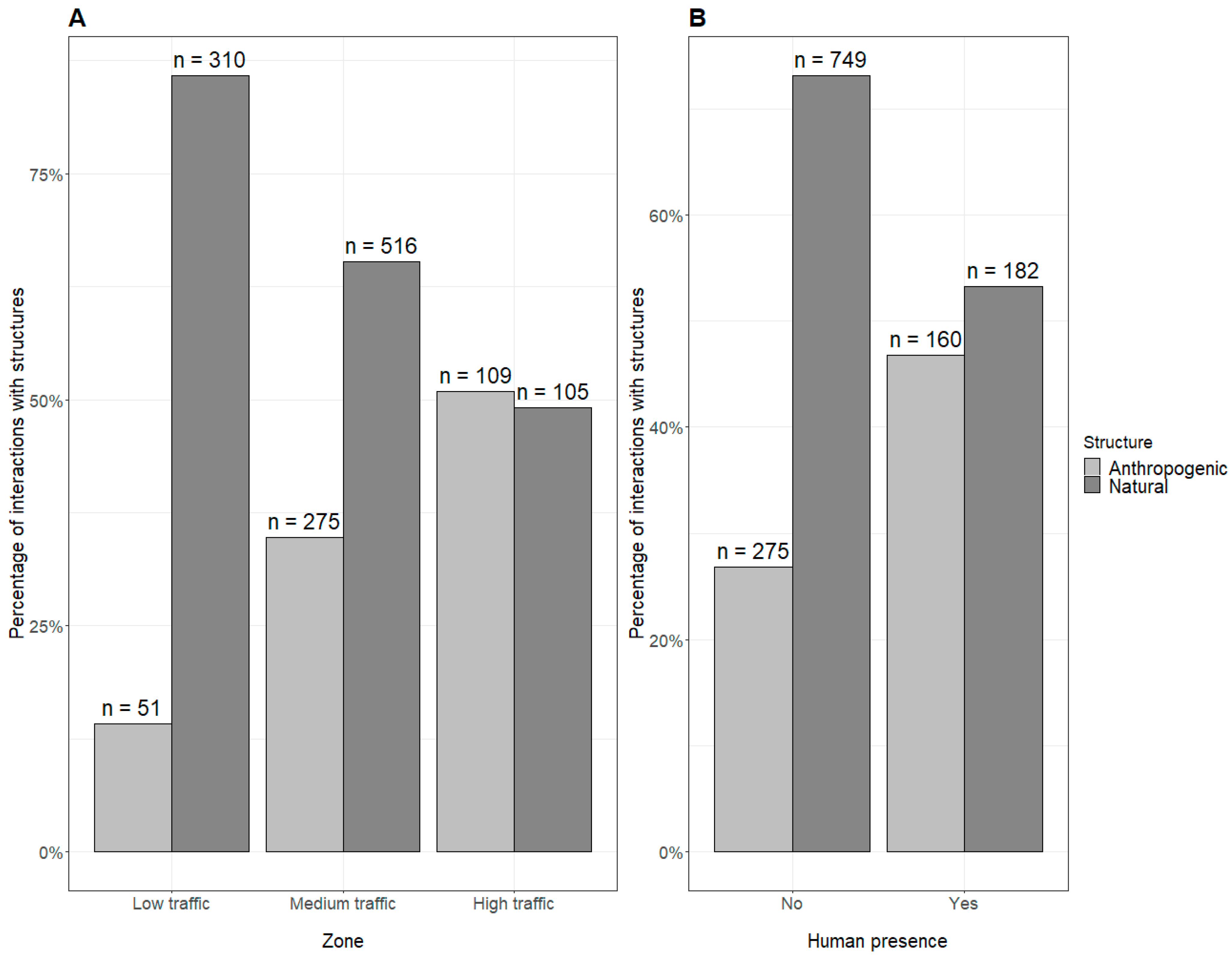
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elmqvist, T.; Zipperer, W.C.; Giineralp, B. Urbanization, Habitat Loss, Biodiversity Decline: Solution Pathways to Break the Cycle. In Routledge Handbook of Urbanization and Global Environmental Change; Karen, S., Solecki, W.D., Griffith, C.A., Eds.; Routledge: London, UK; New York, NY, USA, 2016; Volume 2016, pp. 139–151. [Google Scholar]
- Hughes, A.C. Mapping Priorities for Conservation in Southeast Asia. Biol. Conserv. 2017, 209, 395–405. [Google Scholar] [CrossRef]
- Sodhi, N.S.; Koh, L.P.; Brook, B.W.; Ng, P.K.L. Southeast Asian Biodiversity: An Impending Disaster. Trends Ecol. Evol. 2004, 19, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Wierucka, K.; Hatten, C.E.; Murphy, D.; Allcock, J.A.; Andersson, A.A.; Bojan, J.W.; Kong, T.C.; Kwok, J.K.; Lam, J.Y.; Ma, C.H.; et al. Human-Wildlife Interactions in Urban Asia. Glob. Ecol. Conserv. 2023, 46, e02596. [Google Scholar] [CrossRef]
- Ngo, H.T.; Guèze, M.; Agard Trinidad, J.; Arneth, A.; Balvanera, P.; Brauman, K.; Watson, R.; Baste, I.; Larigauderie, A.; Leadley, P.; et al. Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on; Watson, R., Baste, I., Larigauderie, A., Leadley, P., Pascual, U., Baptiste, B., Demissew, S., Dziba, L., Eds.; IPBES Secretariat: Bonn, Germany, 2019. [Google Scholar]
- Erinjery, J.J.; Kumar, S.; Kumara, H.N.; Mohan, K.; Dhananjaya, T.; Sundararaj, P.; Kent, R.; Singh, M. Losing Its Ground: A Case Study of Fast Declining Populations of a ‘Least-Concern’ Species, the Bonnet Macaque (Macaca radiata). PLoS ONE 2017, 12, e0182140. [Google Scholar] [CrossRef] [PubMed]
- Passamani, M. Activity Budget of Geoffroy’s Marmoset (Callithrix Geoffroyi) in an Atlantic Forest in Southeastern Brazil. Am. J. Primatol. 1998, 46, 333–340. [Google Scholar] [CrossRef]
- Peres, C.A. Diet and Feeding Ecology of Saddle-Back (Saguinus fuscicollis) and Moustached (S. mystax) Tamarins in an Amazonian Terrafirme Forest. J. Zool. 1993, 230, 567–592. [Google Scholar] [CrossRef]
- Hansen, M.F.; Ang, A.; Trinh, T.; Sy, E.; Paramasiwam, S.; Ahmed, T.; Dimalibot, J.; Jones-Engel, L.; Ruppert, N.; Griffioen, C.; et al. Macaca Fascicularis. The IUCN Red List of Threatened Species 2022: e.T12551A199563077. 2022. Available online: https://dx.doi.org/10.2305/IUCN.UK.2022-1.RLTS.T12551A199563077.en (accessed on 10 April 2024).
- Huffman, M.A. The Sacred Monkeys of Bali. Primates 2000, 41, 235. [Google Scholar] [CrossRef]
- Brandon-Jones, D.; Eudey, A.A.; Geissmann, T.; Groves, C.P.; Melnick, D.J.; Morales, J.C.; Shekelle, M.; Stewart, C.B. Asian Primate Classification. Int. J. Primatol. 2004, 25, 97–164. [Google Scholar] [CrossRef]
- Brent, L.; Veira, Y. Social Behavior of Captive Indochinese and Insular Long-Tailed Macaques (Macaca fascicularis) Following Transfer to a New Facility. Int. J. Primatol. 2002, 23, 147–159. [Google Scholar] [CrossRef]
- Hamada, Y.; Suryobroto, B.; Goto, S.; Malaivijitnond, S. Morphological and Body Color Variation in Thai Macaca Fascicularis Fascicularis North and South of the Isthmus of Kra. Int. J. Primatol. 2008, 29, 1271–1294. [Google Scholar] [CrossRef]
- Hambali, K.; Ismail, A.; Md Zain, B.M. Daily Activity Budget of Long-Tailed Macaques (Macaca fascicularis) in Kuala Selangor Nature Park. Int. J. Basic Appl. Sci. IJBAS-IJENS 2012, 12, 124404–126767. [Google Scholar]
- Mohammad, M.; Wong, A. The Daily Activity Budgets of Long-Tailed Macaque (Macaca fascicularis) at Padang Teratak Wildlife Sanctuary, Beaufort, Sabah, Malaysia. J. Trop Biol. Conserv. 2019, 16, 165–183. [Google Scholar]
- Ruslin, F.; Yaakop, S.; Zain, B.M.M. A Preliminary Study on Activity Budget, Daily Travel Distance and Feeding Behaviour of Long-Tailed Macaques and Spectacled Dusky Leaf Monkey in Bangi Campus of Universiti Kebangsaan Malaysia, Selangor. In AIP Conference Proceedings; American Institute of Physics: College Park, MD, USA, 2014. [Google Scholar] [CrossRef]
- Fitriyah, A.; Wahyuningsih, E.; Syaputra, M.; Lestari, A.T.; Isyaturriyadhah. Survey of Long-Tailed Macaque’s Behaviour in Mount Rinjani National Park, Lombok Timur. In IOP Conference Series: Earth and Environmental Science; IOP Publishing Ltd.: Bristol, UK, 2021; Volume 891. [Google Scholar] [CrossRef]
- Ruslin, F.; Matsuda, I.; Md-Zain, B.M. The Feeding Ecology and Dietary Overlap in Two Sympatric Primate Species, the Long-Tailed Macaque (Macaca fascicularis) and Dusky Langur (Trachypithecus obscurus obscurus), in Malaysia. Primates 2019, 60, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.C.; Priston, N.E.C. Human Attitudes to Primates: Perceptions of Pests, Conflict and Consequences for Primate Conservation. In Human Attitudes to Primates; Wiley: Hoboken, NJ, USA, 2005; pp. 1–23. [Google Scholar]
- Priston, N.E.C.; McLennan, M.R. Managing Humans, Managing Macaques: Human-Macaque Conflict in Asia and Africa. In The Macaque Connection: Cooperation and Conflict between Humans and Macaques; Springer: New York, NY, USA, 2013; pp. 225–250. [Google Scholar]
- Burton, F.D. Monkey King in China: Basis for a Conservation Policy? In Primates Face to Face; Cambridge Univeristy Press: Cambridge, UK, 2002; pp. 137–162. [Google Scholar] [CrossRef]
- Knight, J. Monkeys on the Move: The Natural Symbolism of People-Macaque Conflict in Japan. J. Asian Stud. 1999, 58, 622–647. [Google Scholar] [CrossRef]
- Southwick, C.H.; Siddiqi, M.F. India’s Rhesus Populations: Protectionism versus Conservation Management. Monkeys Edge 2011, 297, 275–292. [Google Scholar] [CrossRef]
- Marty, P.R.; Beisner, B.; Kaburu, S.S.K.; Balasubramaniam, K.; Bliss-Moreau, E.; Ruppert, N.; Mohd Sah, S.A.; Ismail, A.; Arlet, M.E.; Atwill, E.R.; et al. Time Constraints Imposed by Anthropogenic Environments Alter Social Behaviour in Longtailed Macaques. Anim. Behav. 2019, 150, 157–165. [Google Scholar] [CrossRef]
- Gilhooly, L.J.; Burger, R.; Sipangkui, S.; Colquhoun, I.C. Tourist Behavior Predicts Reactions of Macaques (Macaca fascicularis and M. nemestrina) at Sepilok Orang-Utan Rehabilitation Centre, Sabah, Malaysia. Int. J. Primatol. 2021, 42, 349–368. [Google Scholar] [CrossRef]
- Jayapali, U.; Perera, P.; Cresswell, J.; Dayawansa, N. Does Anthropogenic Influence on Habitats Alter the Activity Budget and Home Range Size of Toque Macaques (Macaca sinica)? Insight into the Human-Macaque Conflict. Trees For. People 2023, 13, 100412. [Google Scholar] [CrossRef]
- Hambali, K.; Ismail, A.; Munir Md-Zain, B.; Amir, A.; Abdul Karim, F.; Munir, M.-Z.B. Diet of Long-Tailed Macaques (Macaca fascicularis) at the Entrance of Kuala Selangor Nature Park (Anthropogenic habitat): Food Selection That Leads to Human-Macaque Conflict. Acta Biol. Malays. 2014, 3, 58–68. [Google Scholar]
- Brandt, J.S.; Buckley, R.C. A Global Systematic Review of Empirical Evidence of Ecotourism Impacts on Forests in Biodiversity Hotspots. Curr. Opin. Environ. Sustain. 2018, 32, 112–118. [Google Scholar] [CrossRef]
- Muehlenbein, M.P.; Ancrenaz, M. Minimizing Pathogen Transmission at Primate Ecotourism Destinations: The Need for Input from Travel Medicine. J. Travel. Med. 2009, 16, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Muehlenbein, M.P.; Martinez, L.A.; Lemke, A.A.; Ambu, L.; Nathan, S.; Alsisto, S.; Sakong, R. Unhealthy Travelers Present Challenges to Sustainable Primate Ecotourism. Travel. Med. Infect. Dis. 2010, 8, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Russon, A.E.; Wallis, J. Primate Tourism as a Conservation Tool. Primate Tour. 2014, 42, 313–332. [Google Scholar] [CrossRef]
- Williamson, E.A.; Feistner, A.T.C. Habituating Primates: Processes, Techniques, Variables and Ethics. In Field and Laboratory Methods in Primatology; Cambridge University Press: Cambridge, UK, 2011; pp. 33–50. [Google Scholar] [CrossRef]
- Woodford, M.H.; Butynski, T.M.; Karesh, W.B. Habituating the Great Apes: The Disease Risks. Oryx 2002, 36, 153–160. [Google Scholar] [CrossRef]
- McKinney, T. The Effects of Provisioning and Crop-raiding on the Diet and Foraging Activities of Human-commensal White-faced Capuchins (Cebus capucinus). Am. J. Primatol. 2011, 73, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Maréchal, L.; MacLarnon, A.; Majolo, B.; Semple, S. Primates’ Behavioural Responses to Tourists: Evidence for a Trade-off between Potential Risks and Benefits. Sci. Rep. 2016, 6, 32465. [Google Scholar] [CrossRef] [PubMed]
- Goldthorpe, G.; Neo, S.H.A. Preliminary Investigation into the Effects of Hunting on Three Large Ungulate Species in Peninsular Malaysia, with Implications for Tiger Conservation. Malay Nat. J. 2011, 63, 549–560. [Google Scholar]
- Department of Wildlife and National Parks Peninsular Malaysia. Annual Report by Department of Wildlife and National Parks Peninsular Malaysia; Kuala Lumpur, Malaysia, 2018. [Google Scholar]
- Azlan, J.M. Mammal Diversity and Conservation in a Secondary Forest in Peninsular Malaysia. Biodivers. Conserv. 2006, 15, 1013–1025. [Google Scholar] [CrossRef]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef]
- Bateson, P.; Martin, P. Measuring Behaviour: An Introductory Guide, 3rd ed.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 24 February 2024).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Ruppert, N.; Holzner, A.; See, K.W.; Gisbrecht, A.; Beck, A. Activity Budgets and Habitat Use of Wild Southern Pig-Tailed Macaques (Macaca nemestrina) in Oil Palm Plantation and Forest. Int. J. Primatol. 2018, 39, 237–251. [Google Scholar] [CrossRef]
- Sha, J.C.M.; Hanya, G. Diet, Activity, Habitat Use, and Ranging of Two Neighboring Groups of Food-Enhanced Long-Tailed Macaques (Macaca fascicularis). Am. J. Primatol. 2013, 75, 581–592. [Google Scholar] [CrossRef]
- Knight, J. Making Wildlife Viewable: Habituation and Attraction. Soc. Anim. 2009, 17, 167–184. [Google Scholar] [CrossRef]
- Brotcorne, F.; Giraud, G.; Gunst, N.; Fuentes, A.; Wandia, I.N.; Beudels-Jamar, R.C.; Poncin, P.; Huynen, M.-C.; Leca, J.-B. Intergroup Variation in Robbing and Bartering by Long-Tailed Macaques at Uluwatu Temple (Bali, Indonesia). Primates 2017, 58, 505–516. [Google Scholar] [CrossRef]
- Taufet-Rosdi, N.E.A.; Md-Zain, B.M. Human Long-tailed Macaque Interactions and Ecotourism Potential in Gunung Keriang Recreational Park, Kedah Peninsular Malaysia. J. Anim. Plant Sci. 2023, 33, 1021–1032. [Google Scholar] [CrossRef]
- Murali, R.S.N. Uproar over Macaque Culling. Available online: https://www.thestar.com.my/news/nation/2013/03/18/uproar-over-macaque-culling (accessed on 10 November 2023).
- Gumert, M.D.; Fuentes, A.; Lisa, J.E. Monkeys on the Edge Ecology and Management of Long-Tailed Macaques and Their Interface with Humans; Cambridge University Press: Cambridge, UK, 2011; p. 360. [Google Scholar]
- Łopucki, R.; Klich, D.; Kiersztyn, A. Changes in the Social Behavior of Urban Animals: More Aggression or Tolerance? Mamm. Biol. 2021, 101, 1–10. [Google Scholar] [CrossRef]
- Gaynor, K.M.; Cords, M. Antipredator and Social Monitoring Functions of Vigilance Behaviour in Blue Monkeys. Anim. Behav. 2012, 84, 531–537. [Google Scholar] [CrossRef]
- Stankowich, T.; Blumstein, D.T. Fear in Animals: A Meta-Analysis and Review of Risk Assessment. Proc. R. Soc. B Biol. Sci. 2005, 272, 2627. [Google Scholar] [CrossRef]
- Munir Md-Zain, B.; Mohd-Daut, N.; Md-Nor, S. Characterizing Silvered Leaf Monkey Visitor Interaction at Bukit Melawati, Kuala Selangor, Malaysia. J. Wildl. Parks 2009, 26, 83–94. [Google Scholar]
- Špinka, M.; Newberry, R.C.; Bekoff, M. Mammalian Play: Training for the Unexpected. Q. Rev. Biol. 2001, 76, 141–168. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, H.; Qi, X.; Wang, C.; Yang, B.; Li, B. Behavioral Patterns, Influencing Factors, Functions and Risks of Social Play in Primates. Acta Ecol. Sin. 2012, 32, 2910–2917. [Google Scholar] [CrossRef]
- Symons, D. Play and Aggression: A Study of Rhesus Monkeys; Columbia University Press: New York, NY, USA, 1978. [Google Scholar] [CrossRef]
- Ciani, F.; Dall’Olio, S.; Stanyon, R.; Palagi, E. Social Tolerance and Adult Play in Macaque Societies: A Comparison with Different Human Cultures. Anim. Behav. 2012, 84, 1313–1322. [Google Scholar] [CrossRef]
- Kappeler, P.M.; Cremer, S.; Nunn, C.L. Sociality and Health: Impacts of Sociality on Disease Susceptibility and Transmission in Animal and Human Societies. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140116. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, M.; Lane, M.A. Sociality, Hierarchy, Health: Comparative Biodemography: A Collection of Papers; National Academies Press: Washington, DC, USA, 2014. [Google Scholar] [CrossRef]
- Schülke, O.; Bhagavatula, J.; Vigilant, L.; Ostner, J. Social Bonds Enhance Reproductive Success in Male Macaques. Curr. Biol. 2010, 20, 2207–2210. [Google Scholar] [CrossRef] [PubMed]
- Silk, J.B.; Beehner, J.C.; Bergman, T.J.; Crockford, C.; Engh, A.L.; Moscovice, L.R.; Wittig, R.M.; Seyfarth, R.M.; Cheney, D.L. The Benefits of Social Capital: Close Social Bonds among Female Baboons Enhance Offspring Survival. Proc. R. Soc. B Biol. Sci. 2009, 276, 3099–3104. [Google Scholar] [CrossRef]
- Silk, J.B.; Beehner, J.C.; Bergman, T.J.; Crockford, C.; Engh, A.L.; Moscovice, L.R.; Wittig, R.M.; Seyfarth, R.M.; Cheney, D.L. Strong and Consistent Social Bonds Enhance the Longevity of Female Baboons. Curr. Biol. 2010, 20, 1359–1361. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, R.I.M. Time: A Hidden Constraint on the Behavioural Ecology of Baboons. Behav. Ecol. Sociobiol. 1992, 31, 35–49. [Google Scholar] [CrossRef]
- Kappeler, P.M.; Van Schaik, C.P. Evolution of Primate Social Systems. Int. J. Primatol. 2002, 23, 707–740. [Google Scholar] [CrossRef]
- Oliveira, A.F.S.; Rossi, A.O.; Silva, L.F.R.; Lau, M.C.; Barreto, R.E. Play Behaviour in Nonhuman Animals and the Animal Welfare Issue. J. Ethol. 2009, 28, 1–5. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavior | Variable | Odds Ratio | p-Value | McFadden’s R2 |
|---|---|---|---|---|
| Travel | Intercept | 0.00 | <0.001 | 0.0398 |
| Anthropogenic structure | 1.39 | <0.001 | ||
| Sunny weather | 0.85 | <0.001 | ||
| Human presence | 1.06 | NS | ||
| Medium traffic area | 0.94 | NS | ||
| High traffic area | 1.07 | NS | ||
| Inactive | Intercept | 0.30 | <0.001 | 0.0359 |
| Anthropogenic structure | 0.82 | <0.001 | ||
| Sunny weather | 1.09 | <0.05 | ||
| Human presence | 1.12 | <0.05 | ||
| Medium traffic area | 1.49 | <0.001 | ||
| High traffic area | 0.56 | <0.001 | ||
| Affiliative Interaction | Intercept | 0.21 | <0.001 | 0.057 |
| Anthropogenic structure | 0.57 | <0.001 | ||
| Sunny weather | 0.90 | <0.05 | ||
| Human presence | 0.49 | <0.001 | ||
| Medium traffic area | 1.67 | <0.001 | ||
| High traffic area | 1.54 | <0.001 | ||
| Feeding Natural | Intercept | 0.26 | <0.001 | 0.0558 |
| Anthropogenic structure | 1.16 | <0.05 | ||
| Sunny weather | 1.16 | <0.01 | ||
| Human presence | 1.17 | <0.05 | ||
| Medium traffic area | 0.34 | <0.001 | ||
| High traffic area | 0.18 | <0.001 | ||
| Feeding Unnatural | Intercept | 0.00 | <0.001 | 0.1063 |
| Anthropogenic structure | 12.18 | <0.001 | ||
| Sunny weather | 2.61 | <0.001 | ||
| Human presence | 1.25 | <0.001 | ||
| Medium traffic area | 3.39 | <0.001 | ||
| High traffic area | 3.03 | NS | ||
| Self grooming | Intercept | 0.03 | <0.001 | 0.0513 |
| Anthropogenic structure | 0.80 | NS | ||
| Sunny weather | 0.84 | NS | ||
| Human presence | 1.93 | <0.001 | ||
| Medium traffic area | 0.98 | NS | ||
| High traffic area | 0.92 | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Entezami, M.; Mustaqqim, F.; Morris, E.; Lim, E.S.H.; Prada, J.M.; Paramasivam, S.J. Effect of Human Activity and Presence on the Behavior of Long-Tailed Macaques (Macaca fascicularis) in an Urban Tourism Site in Kuala Selangor, Malaysia. Animals 2024, 14, 1173. https://doi.org/10.3390/ani14081173
Entezami M, Mustaqqim F, Morris E, Lim ESH, Prada JM, Paramasivam SJ. Effect of Human Activity and Presence on the Behavior of Long-Tailed Macaques (Macaca fascicularis) in an Urban Tourism Site in Kuala Selangor, Malaysia. Animals. 2024; 14(8):1173. https://doi.org/10.3390/ani14081173
Chicago/Turabian StyleEntezami, Mahbod, Fiqri Mustaqqim, Elizabeth Morris, Erin Swee Hua Lim, Joaquín M. Prada, and Sharmini Julita Paramasivam. 2024. "Effect of Human Activity and Presence on the Behavior of Long-Tailed Macaques (Macaca fascicularis) in an Urban Tourism Site in Kuala Selangor, Malaysia" Animals 14, no. 8: 1173. https://doi.org/10.3390/ani14081173
APA StyleEntezami, M., Mustaqqim, F., Morris, E., Lim, E. S. H., Prada, J. M., & Paramasivam, S. J. (2024). Effect of Human Activity and Presence on the Behavior of Long-Tailed Macaques (Macaca fascicularis) in an Urban Tourism Site in Kuala Selangor, Malaysia. Animals, 14(8), 1173. https://doi.org/10.3390/ani14081173