Simple Summary
In the process of domestication and urbanization, people and small companion animals share, to varying degrees, their dietary style and living environment, and they also encounter similar lifestyle challenges, such as obesity and other diseases. In our study, Lactobacillus acidophilus can improve the body’s lipid metabolism and immune and antioxidant capacity by regulating the relative abundance of intestinal bacterial community, which enables companion animals more easily adapt to the lifestyle of lower level of physical activity in the city, thereby improving animal health and well-being.
Abstract
Lactobacillus acidophilus (L. acidophilus), the most prevalent probiotic, has demonstrated the ability to improve the relative abundance of intestinal microorganisms and boost immunity. However, the underlying mechanisms of these effects remain unclear. This study evaluated body weight, nutrient apparent digestibility, serum indices, and bacterial communities in Chinese rural dogs from a L. acidophilus supplementation group (Lactobacillus acidophilus, n = 6) and a control group (CON, n = 6). The results indicated that L. acidophilus had no significant impact on the body weight and apparent nutrient digestibility of Chinese rural dogs. In comparison with the CON group, L. acidophilus significantly reduced the levels of cholesterol (CHO) and increased the levels of IgA, IFN-α, and T-AOC. Bacterial diversity indices were significantly reduced in the LAC group compared to the CON groups, and MetaStat analysis demonstrated notable distinctions in 14 bacterial genera between the groups. These bacterial genera exhibited correlations with physiological indices such as CHO, IgA, IFN-α, and T-AOC. In conclusion, L. acidophilus can modulate lipid metabolism, immunity, and antioxidant capacity by regulating the relative abundance of specific bacterial communities, which helps dogs to adapt to today’s lifestyle.
1. Introduction
Lactobacillus acidophilus (L. acidophilus) is a prevalent probiotic characterized by good resistance against acid and bile salts [1], which ensures its survival in hostile environment. And it also can colonize complex gastrointestinal ecosystem, with beneficial effects accumulating over a longer period of time compared to non-colonizing probiotics [2]. Beyond its survival mechanisms, L. acidophilus exhibits the ability to metabolize dietary phytoglycosides and externalize its bioactive phytochemicals [3]. Furthermore, it plays a pivotal role in the body’s immune system, with studies demonstrating its effectiveness in reducing cholesterol (CHO) levels and combating pathogenic bacteria [4,5]. Its potential extends to improving the progression of nonalcoholic steatosis by lowering CHO, establishing it as an effective strategy for treating nonalcoholic fatty liver disease (NAFLD) [6]. And L. acidophilus also has demonstrated promising potential in ameliorating type 2 diabetes and obesity by modulating CHO levels and the abundance of the intestinal bacterial community in murine models [7,8].
In the process of domestication and urbanization, dogs and humans exhibit remarkable consistency in terms of dietary structure and living environment, particularly in developed countries [9,10,11]. There are also similarities between dog and human gut microbiota in terms of gene content and response to diet [12]. Given that most dogs are subjected to high-carbohydrate diets and encounter similar lifestyle challenges as humans [13], the metabolic dysfunctions associated with obesity in dogs can result in a significant increase in serum total CHO and triglycerides (TG) [14,15]. With dogs assuming pivotal roles as companions in people’s lives, the management of their nutrition and health are worthy of investigation and consideration. Notably, L. acidophilus has been reported to enhance growth in dogs [16]. Specifically, the L. acidophilus strain DSM13241 can be successfully incorporated into a dry diet and survive in the canine or feline gastrointestinal tract. This incorporation is associated with an elevation in the concentration of IgG in dogs, accompanied by an increase in the relative abundance of fecal Lactobacilli and a decrease in the relative abundance of Clostridial organisms [17], and it has been observed to reduce fecal pH, elevate the relative abundance of beneficial Lactobacillus and L. acidophilus, while decreasing the relative abundance of Clostridium spp. and Enterococcus faecalis in cats [18]. Additionally, L acidophilus D2/CSL (CECT 4529) significantly improved the nutritional status and fecal parameters in dogs [19]. These findings suggest that this L. acidophilus strain holds the potential to enhance the balance of the bacterial community in dogs and cats, thereby contributing to improved intestinal health and immune function.
This study investigated the effects of L. acidophilus on digestibility, metabolism, and serum indices in dogs, with the main aim of developing a novel probiotic to provide valuable insights into dog nutrition and health research, offering guidance for the formulation of pet food.
2. Materials and Methods
2.1. L. acidophilus
L. acidophilus was isolated and preserved in our laboratory. The DeMan-Rogosa-Sharpe (MRS) medium was used to resuscitate and passage cultures for 36 h at 37 °C, and the concentration of bacterial solution was determined to be 1 × 108 CFU/mL.
2.2. Experimental Animals and Study Design
A total of 12 three-month-old Chinese rural dogs, with a mean body weight of 4.68 ± 0.93 kg, were recruited for this study, and the ratio of male to female dogs was equal. Each dog was housed in a cage, all animals were kept under controlled conditions of room temperature, humidity, and a 12/12 h light/dark cycle, and the dogs were provided with two daily feedings and unrestricted access to laboratory water. One month prior to the commencement of the experiment, all of the dogs underwent vaccination and deworming procedures following routine immunization protocols. Specifically, each animal received the canine quadruple vaccine and underwent deworming based on fecal examination and overall physical condition.
After a minimum acclimatization period of 3 days in laboratory conditions, the dogs were randomly assigned to either the control group (CON) or the L. acidophilus group (LAC) with 6 replicates per group. All of the animals were fed dry kibble food twice daily and had unrestricted access to laboratory water to meet the nutrient requirement of dogs at maintenance [20]. Dogs in the LAC group received 1 mL L. acidophilus culture solution, while those in the CON group were administered an equal volume (1 mL) of normal saline orally after feeding. After each feeding, the syringe containing the bacteria solution or normal saline was injected into the dog’s mouth to ensure full swallowing. The average daily feed intake per animal was approximately 200 g/d during the pretest, and the dose of probiotics was calculated to be 2 × 108 CFU/day (European Standard No. EN 15878:2021; Animal feeding stuffs: Methods of sampling and analysis -Detection and enumeration of lactobacillus spp. used as feed additive. UNE EN 15787:2022; Animal feeding stuffs: Methods of sampling and analysis - Detection and enumeration of Lactobacillus spp. used as feed additive. UNE EN 15787:2022. compound feeds, meal or pellets which contain about 109 CFU/kg Lactobacillus species) [19,21]. The entire experiment spanned 28 days, and the composition and nutritional content of the basal diet are detailed in Table 1.

Table 1.
Composition and nutrient levels of basal diets (air-dry basis).
2.3. Sample Collection
On the 20th day of the experiment, fecal samples from all dogs were collected over four consecutive days. The samples were fixed with 10% H2SO4 and stored at −20 °C for the subsequent digestibility trial. On the 28th day of the experiment, 10 mL of blood per dog was obtained from the dorsal subcutaneous cephalic vein of the forelimb of the animal using a disposable negative pressure sampling vessel with separation glue coagulant. The collected blood samples were then centrifuged at 3500× g for 10 min at 4 °C to extract the serum. The serum was utilized to determine the concentrations of serum indices immediately. Furthermore, approximately 5 g of feces from the animals were collected immediately after defecation at the conclusion of the experiment. These fecal samples were stored at −80 °C for subsequent bacterial community analysis.
2.4. Measurement of Body Weight
On the morning of the 7th, 14th, 21st, and 28th day of the experiment, the body weight of the experimental animals was measured on an empty stomach. The total weight gain (TWG), average daily weight gain (ADG), and weight gain rate (WGR) were calculated using the following formulas:
TWG = final weight − initial weight
ADG = (final weight − initial weight)/28
WGR = (final weight − initial weight)/initial weight
2.5. Measurement of Nutrient Apparent Digestibility
The diet and fecal samples underwent thawing and drying at 65 °C for 48 h, followed by grinding through a 1 mm sieve before subsequent analysis. Duplicate analyses were conducted on both feed and fecal samples to determine dry matter (DM, method No. 934.01), crude protein (CP, method No. 954.01), and ether extract (EE, method No. 920.39), in accordance with AOAC methods [22]. Carbohydrate content was calculated using the formula: Carbohydrate (%) = DM − CP − EE − Ash (crude ash).
2.6. Measurement of Serum Indices
The levels of serum lipid indices were assessed using Biosino Bio-Technology and Science Inc. (Beijing, China) by an automatic biochemical analyzer (VITALIB-E). This included the measurement of triglycerides (TG), cholesterol (CHO), high-density lipoprotein cholesterol (HDL-C), and Low-Density Lipoprotein Cholesterol (LDL-C). The levels of immune indices (IgA, IgM, IFN-α, IL-2) were determined using Enzyme-Linked Immunosorbent Assay (ELISA) kits from Jianglai Biocompany, (Shanghai, China). Additionally, the levels of antioxidant indices (T-AOC, SOD, GSH-Px) were measured using commercial kits from Jiancheng Bioengineering Institute (Nanjing, China).
2.7. Bacterial Community Analysis
Total genomic DNA in the fecal samples was extracted using the SDS method, and the purity and concentration of the DNA were verified through 1% agarose gel electrophoresis. Specific primers with barcodes (341F: 5′-CCTAYGGGRBGCASCAG-3′, 806R: 5′-GGACTACNNGGGTATCTAAT-3′) were employed for amplification, depending on the selected region for sequencing [23]. The resulting amplicons were sequenced on the Illumina Novaseq 6000 (San Diego, CA, USA) to generate paired-end reads.
Species annotation was conducted using the QIIME2 software (Verision 1.9.1) for 16S rRNA gene sequences, with the Silva Database serving as the annotation database. GraphPad Prism 9.4.1 software was utilized to analyze differences in α diversity indices and bacterial composition between groups. To assess the complexity of bacterial community composition and compare differences among groups, β diversity was calculated based on Jaccard and unweighted UniFrac distances using QIIME2. Principal coordinate analysis (PCoA) was performed to generate principal coordinates and visualize sample differences in multidimensional data. The PCoA results were visualized using the ade4 and ggplot2 packages in R software (Version 2.15.3).
2.8. Statistical Analysis
Statistical analysis was conducted using GraphPad Prism 9.4.1 software. The t test was employed to assess the statistical significance of body weight, nutrient apparent digestibility, serum indices, and α diversity indices between the CON and LAC groups. Anoism analysis was utilized to calculate β diversity indices. The differentiation of bacterial genera between the two groups was analyzed using MetaStat. Statistical significance was defined as p < 0.05 (*), p < 0.01 (**), and p < 0.001 (***). The correlations between bacterial community and physiological indices were evaluated using the Spearman rank correlation coefficient. Network visualization was performed using R 4.2.2 to illustrate the relationships.
3. Results
3.1. Effects of Lactobacillus acidophilus on Body Weight in Chinese Rural Dogs
As indicated in Table 2, the body weight on days 7, 14, 21, and 28 exhibited a slight increase in the LAC group compared to the CON group; however, the differences were not statistically significant (p > 0.05). Similarly, the ADG and WGR also displayed a marginal increase, but the differences were not statistically significant (p > 0.05).
Table 2.
Effects of Lactobacillus acidophilus on Body Weight in Chinese Rural Dogs.
3.2. Effect of Lactobacillus acidophilus on Apparent Digestibility of Nutrients in Chinese Rural Dogs
As illustrated in Table 3, the apparent digestibility of nutrients, including DM, CP, EE, and carbohydrates, were not statistically significant compared to the CON group (p > 0.05).
Table 3.
Effect of Lactobacillus acidophilus on Apparent Digestibility of Nutrients in Chinese Rural Dogs.
3.3. Effect of Lactobacillus acidophilus on Serum Indices in Chinese Rural Dogs
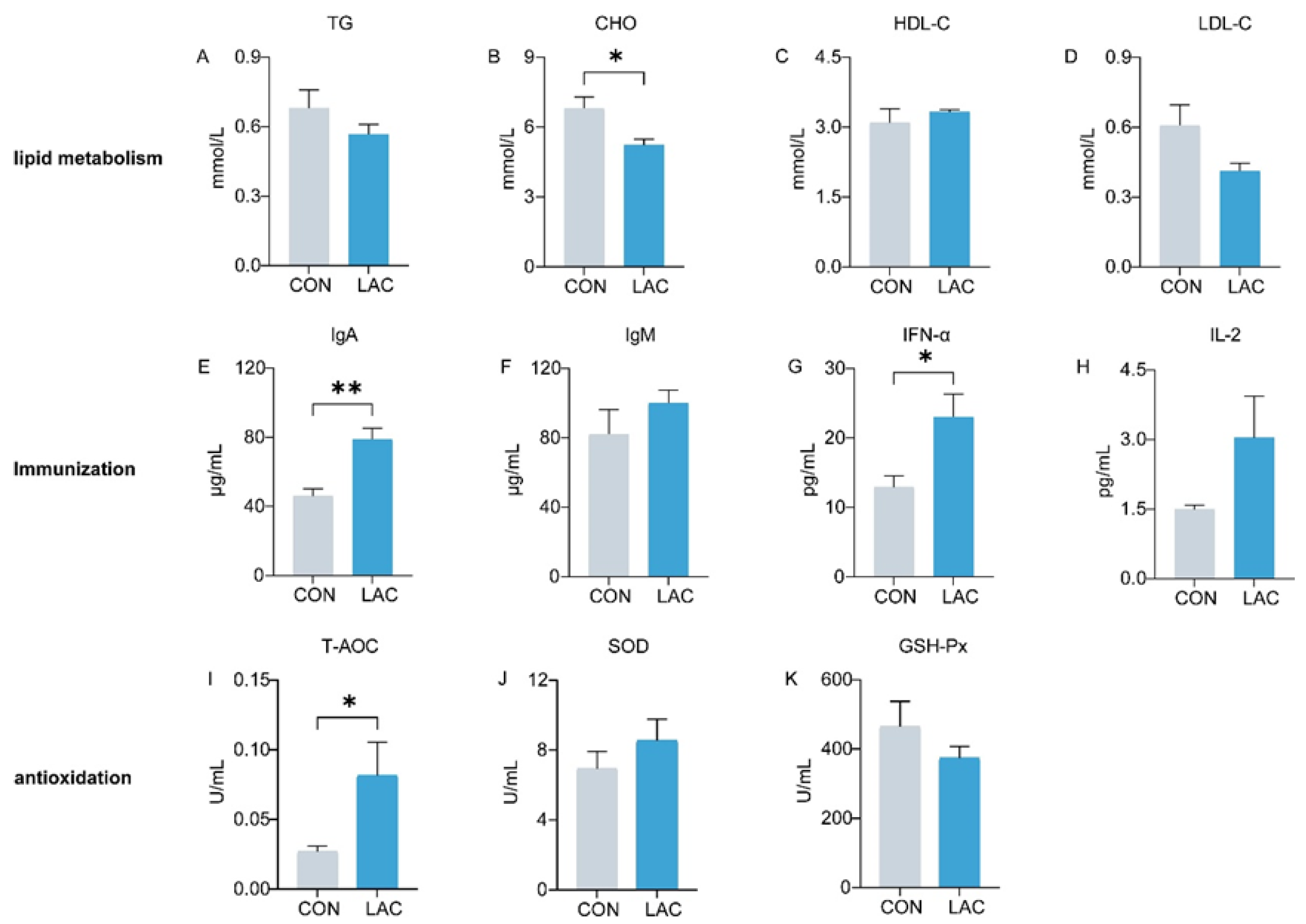
Figure 1 depicts the impact of L. acidophilus on serum lipid metabolism, immunity, and antioxidation in dogs. Comparative analysis with the CON group revealed a significant reduction in CHO levels in the LAC group (p < 0.05) (Figure 1B). TG and LDL exhibited a decrease in the LAC group, although the differences were not statistically significant (p > 0.05) (Figure 1A,D). In terms of immune indices, IgA in the LAC group demonstrated a remarkable increase compared to the CON group (p < 0.01) (Figure 1E), and IFN-α exhibited a significant elevation (p < 0.05) (Figure 1G). IgM and IL-2 also showed an increase, though the differences were not significant (p > 0.05) (Figure 1F,H). Regarding antioxidation, T-AOC showed a significant increase in the LAC group (p < 0.05) (Figure 1I), whereas the activity of SOD did not exhibit a significant difference between the two groups (p > 0.05) (Figure 1J).
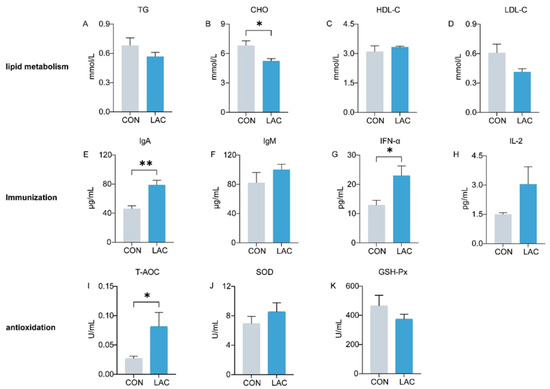
Figure 1.
Effects of L. acidophilus on serum indices of Chinese rural dogs. (A–D) Comparison of lipid metabolism indices between the CON and LAC groups. (E–H) Comparison of immunological indices between the CON and LAC groups. (I–K) Comparison of antioxidant indices between the CON and LAC groups. * and ** indicate p < 0.05 and p < 0.01, respectively. TG: triglycerides, CHO: cholesterol, HDL-C: high-density lipoprotein cholesterol, LDL-C: low-density lipoprotein, IgA: immunoglobulin A, IgM: immunoglobulin M, IFN-α: interferon α, IL-2: interleukin 2, T-AOC: total antioxidant capacity, SOD: Superoxide dismutase, GSH-Px: Glutathione Peroxidase.
3.4. Effects of Lactobacillus acidophilus on the Bacterial Community in Chinese Rural Dogs
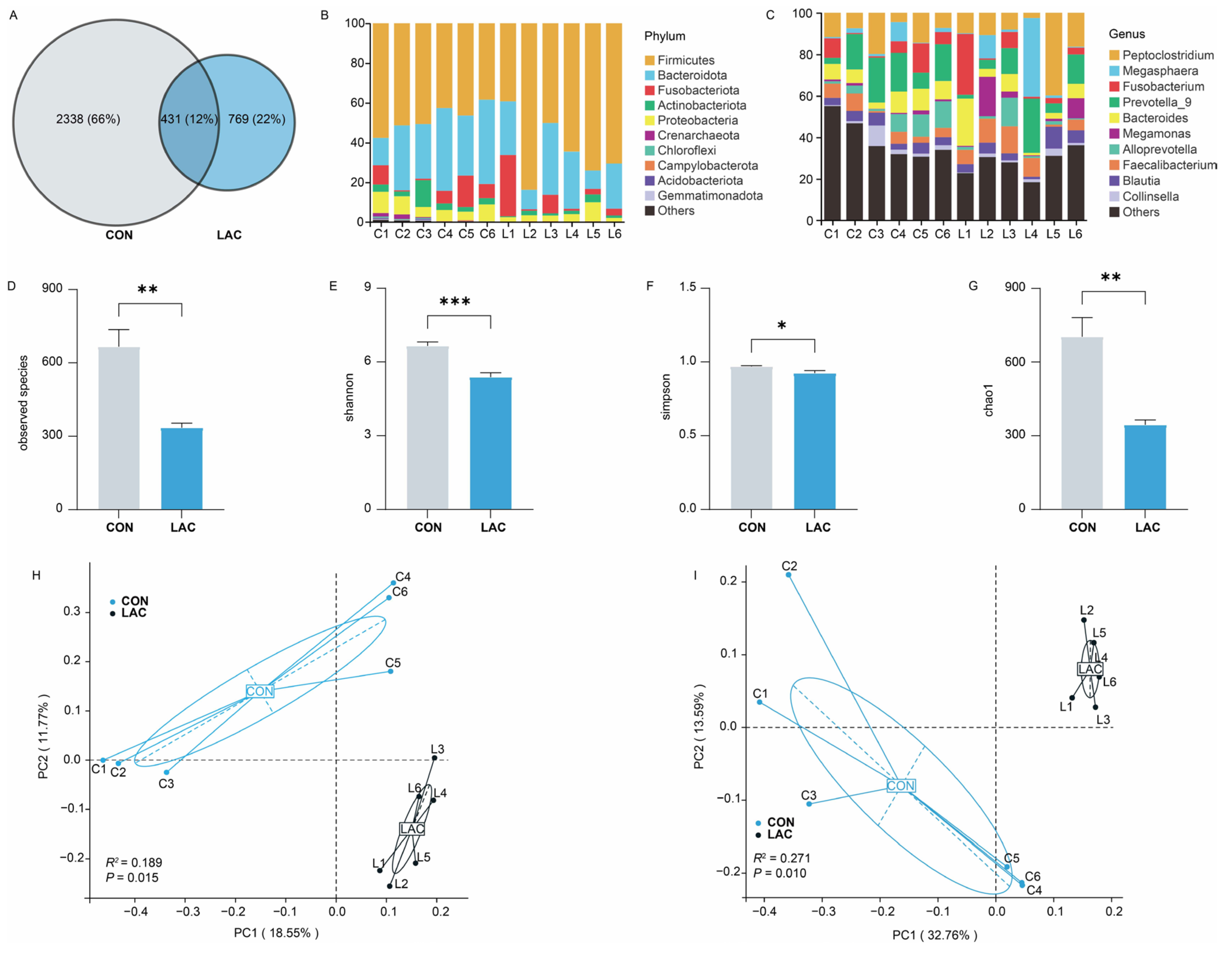
To explore the impact of L. acidophilus on the bacterial community of Chinese rural dogs, a total of 431 amplicon sequence variants (ASVs) were shared in the two groups, with 2338 and 769 unique ASVs in the CON and LAC groups, respectively (Figure 2A). At the phylum level, Firmicutes and Bacteroidota were the most abundant bacteria in both groups, followed by Proteobacteria in the CON group and Fusobacteriota in the LAC group, collectively accounting for 86.41% in the CON group and 93.99% in the LAC group of the bacterial composition (Figure 2B). Moreover, 476 bacterial genera were identified from the feces of dogs, with Prevotella_9 (CON = 14.30%, LAC = 10.59%) and Peptoclostridium (CON = 10.88%, LAC = 14.57%) being the predominant genera (Figure 2C).
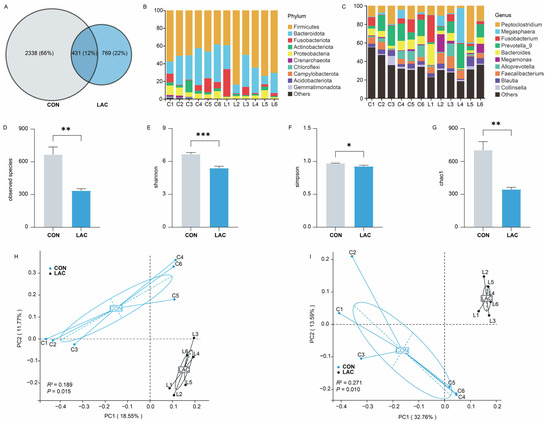
Figure 2.
Effects of L. acidophilus on the composition and diversity of bacterial communities in Chinese rural dogs. (A) Shared and unique bacterial ASVs between the two groups. (B) Bacterial community composition at the phylum level in feces. (C) Bacterial community composition at the genus level in feces. (D) Composition of observed species of bacterial diversity between the two groups. (E) Composition of Shannon indices of bacterial diversity between the two groups. (F) Composition of Simpson indices of bacterial diversity between the two groups. (G) Composition of the Chao 1 indices of bacterial diversity between the two groups. (H) Composition of bacterial community based on Jaccard distance between the two groups. (I) Composition of bacterial community based on unweighted UniFrac distance between the two groups. *, ** and *** indicate p < 0.05, p < 0.01 and p < 0.001, respectively.
Assessment of bacterial community diversity between the CON and LAC groups revealed that the number of observed species, the Shannon and Chao 1 indices, were significantly lower in the LAC group than in the CON group (p < 0.01) (Figure 2D,E,G), while the Simpson indices were significantly lower in the LAC group than in the CON group (p < 0.05) (Figure 2F). This suggests that L. acidophilus significantly reduces the species number and bacterial community richness in feces. The PCoA results demonstrated significant differences in fecal bacterial community and its composition between the two groups based on Jaccard (Anoism: R2 = 0.189, p = 0.015) and unweighted UniFrac (Anoism: R2 = 0.271, p = 0.010) distances, explaining at least 30.32% and 46.35% of the variation, respectively (Figure 2H,I).
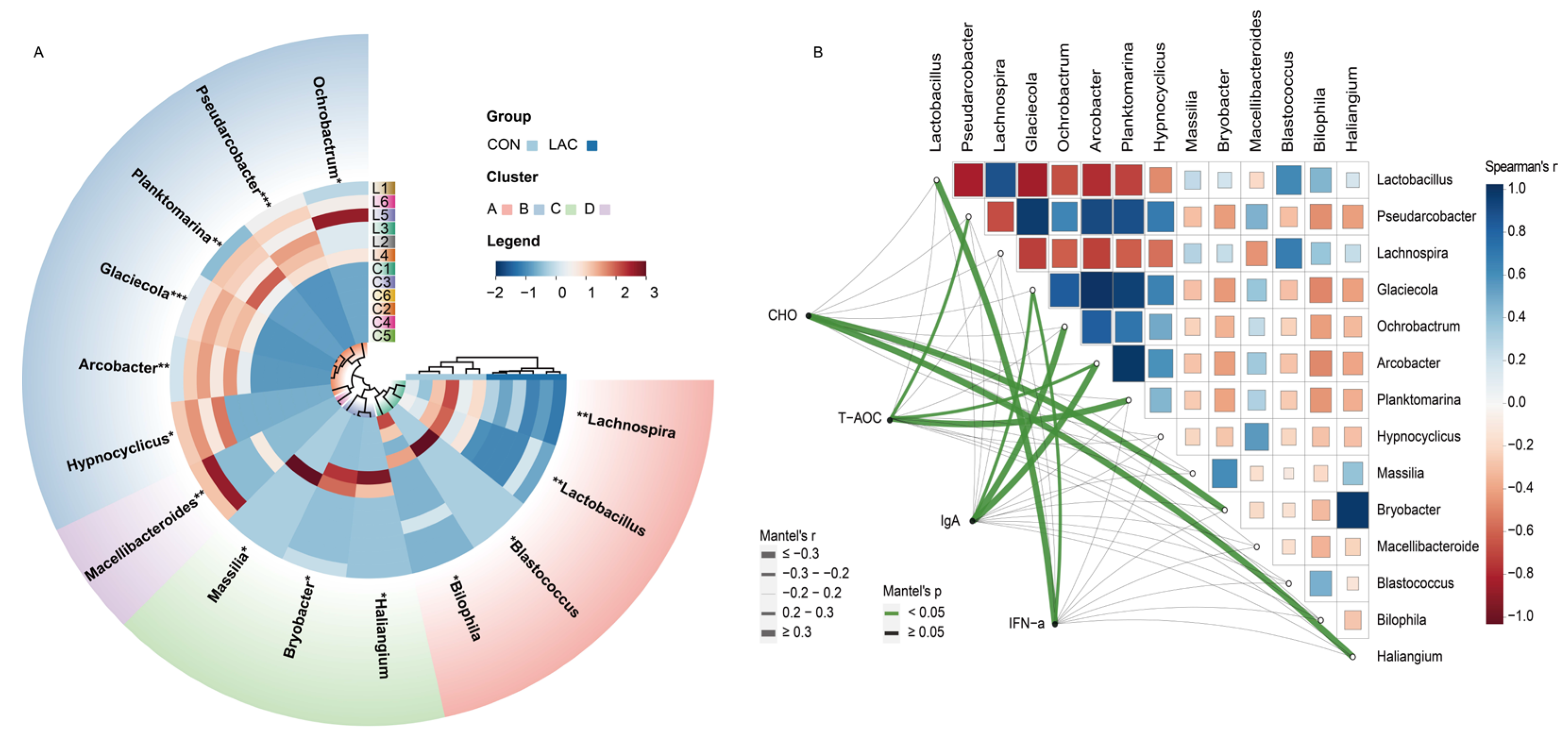
Furthermore, we identified 14 differential genera between the two groups through MetaStat analysis (Figure 3A). Specifically, the relative abundance of Pseudarcobacter and Glaciecola was extremely significantly increased in the LAC group compared to the CON group (p < 0.001). Planktomarina, Macellibacteroides, and Arcobacter were also extremely significantly increased in the LAC group compared to the CON group (p < 0.01). Ochrobactrum and Hypnocyclicus were significantly increased in the LAC group compared to the CON group (p < 0.05). However, Lactobacillus and Lachnospira were significantly decreased in the LAC group compared to the CON group (p < 0.01), while Massilia, Bryobacter, Haliangium, Bilophila, and Blastococcus were significantly decreased in the LAC group compared to the CON group (p < 0.05) (Figure 3A).
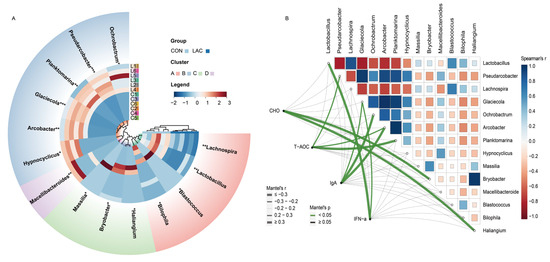
Figure 3.
Correlation analysis between bacterial community differences and physiological indices in Chinese rural dogs. (A) Comparison of different bacterial genera by MetaStat analysis. (B) Analysis of correlation between bacterial genera and physiological indices. *, ** and *** indicate p < 0.05, p < 0.01 and p < 0.001, respectively. Spearman’s correlation coefficients are denoted with a color gradient. Physiological indices were related to each bacterial genus community composition environmental factor by Mantel tests. Edge width corresponds to the Mantel’s r statistic for the corresponding distance correlations, and edge color denotes the statistical significance.
For a deeper exploration of potential connections between the different bacterial community and physiological indices, Spearman’s rank correlation analysis was conducted to ascertain the correlation coefficients among differentially abundant bacteria. The results revealed that Lactobacillus, Pseudarcobacter, Lachnospira, Glaciecola, Ochrobactrum, Arcobacter, Planktomarina, and Hypnocyclicus exhibited the most significant correlations (Figure 3B). Correlation coefficients between differentially abundant bacteria and key indicators such as CHO, T-AOC, IgA, and IFN-α were analyzed by the Mantel test. The results indicated that Bryobacter and Haliangium were correlated with CHO, Pseudarcobacter, Arcobacter, and Planktomarina were correlated with T-AOC, Glaciecola, Ochrobactrum, and Arcobacter were correlated with IgA, while Lactobacillus and Glaciecola were correlated with IFN-α (Figure 3B).
4. Discussion
Probiotics, defined as living microorganisms conferring health benefits when administered in sufficient quantities, play a vital role in promoting host well-being [24]. Among these, L. acidophilus stands out as the most commonly utilized probiotic in various applications. Research has demonstrated its efficacy, such as reducing yolk cholesterol in laying hens and positively impacting their health and performance when incorporated into their diet [25]. L. acidophilus NCFM has also been found to influence intestinal carbohydrate, bile acid, and vitamin E metabolism [26]. The impact of probiotics on animal body weight is known to be contingent upon factors such as specific strains, probiotic dosage, and intervention duration [27]. However, our study revealed that L. acidophilus did not exert significant effects on body weight and the apparent digestibility of nutrients in Chinese rural dogs. This outcome could be attributed to the relatively short intervention duration of L. acidophilus and the heterogeneity of Chinese rural dogs, potentially obscuring any significant changes in animal body weight within the test results.
Hypercholesterolemia can result from various factors, including gain-of-function mutations in proprotein convertase subtilisin kexin 9 (PCSK9), leading to decreased low-density lipoprotein (LDL) receptor levels, and loss-of-function variants associated with reduced LDL-cholesterol (LDL-C) levels and decreased coronary heart disease (CHD) risk [28,29,30,31]. Studies indicate that L. acidophilus can contribute to lowering cholesterol (CHO) levels through mechanisms such as inhibiting 3-hydroxy-3-methylglutaryl CoA reductase and promoting the excretion of dietary cholesterol in feces through co-precipitation of deconjugated bile acids in the intestine and/or adsorption by organisms [2]. Furthermore, L. acidophilus NS1 has been shown to increase the expression of sterol regulatory element-binding protein 2 (SREBP2) and low-density lipoprotein receptor (LDLR) in the liver, resulting in decreased total cholesterol and LDL cholesterol levels in plasma, with no significant change in high-density lipoprotein (HDL) [32]. Additionally, it can enhance lipid metabolism and insulin sensitivity through the SREBP-1c/PPARα signaling pathway, thereby preventing dietary obesity and related metabolic disorders [33]. The observed decrease in CHO and LDL levels in our study suggests that L. acidophilus may be a potential intervention for mitigating dietary obesity and related metabolic disorders.
The variable and stable regions of immunoglobulin interact synergistically to target the destruction and elimination of pathogenic microorganisms and toxins in the body [34]. IgA is widely recognized as a crucial antibody isotype responsible for safeguarding mucosal surfaces, its primary function involves immune exclusion, effectively preventing the entry of foreign substances [35]. Research has demonstrated that the unique C-terminal tail of IgA can impede the attachment of influenza A virus and other enveloped viruses using sialic acid as receptors on the cell surface [36]. Additionally, IgA can neutralize or clear pathogens by triggering mechanisms such as the IgA Fc receptor (FC-RI or CD89) on phagocytes, playing a crucial role in preventing the invasion of microorganisms on mucosal surfaces [37,38,39,40]. In our study, L. acidophilus significantly increased IgA serum content. This suggests that L. acidophilus can enhance the mucosal defense barrier in young dogs, effectively preventing the infection of specific pathogens.
Interferon regulatory factors (IRFs) constitute a family of transcription factors crucial in host defense, activating the transcription of IFN-α and other immune response genes upon activation. IFN-α occupies a pivotal role at the interface of innate and adaptive immunity in viral defense, making it instrumental in setting thresholds for autoimmunity. IFN-α enhances cytokine secretion, polyfunctionality, degranulation, and the cytotoxic potential of NK cells, while also augmenting viral inhibition by NK cells [41]. IL-2 serves as the principal growth factor for antigen-activated T lymphocytes, exerting control over autoimmunity through the production of CD4+ and CD25+ to regulate T cells [42]. In our study, both IFN-α and IL-2 serum levels increased to some extent, suggesting that L. acidophilus can enhance the body’s immunity. Regarding antioxidant effects, L. acidophilus ATCC 4356 has been shown to mitigate the development of atherosclerotic lesions in ApoE(−/−) mice by reducing oxidative stress and inflammatory responses [43]. The determination of antioxidant capacity helps to evaluate the physiological, environmental, and nutritional factors of the redox status of the body, which can provide information about the absorption and bioavailability of nutritional compounds [44]. Our results also support the notion that L. acidophilus possesses certain antioxidant capabilities.
The gastrointestinal tract serves as a pivotal physiological interface that integrates nutrient metabolism and microbiota-host interactions. During gut digestion, the collab-oration between hosts and microbes yields various enzymes, hormones, vitamins, and chemicals like short-chain fatty acids (SCFAs), bile acids, and conjugated linoleic acid (CLAs). These compounds play a critical role in regulating numerous host microbiome pathways, including those governing lipid levels. The antagonistic effect of L. acidophilus against pathogens and other organisms may stem from mechanisms such as nutrient and adhesion site competition, the production of metabolites like organic acids and hydrogen peroxide, as well as the synthesis of antibiotic-like compounds and bacteriocins [2]. Our study demonstrated that L. acidophilus significantly reduced the diversity of the fecal bacterial community and resulted in a more concentrated sample distribution in dogs, it may be that L. acidophilus inhibits the growth and reproduction of some bacterial communities. Dominant bacterial phyla in the LAC group included Firmicutes, Bacteroidetes, and Fusobacteriota, aligning with previous research findings [45,46]. Furthermore, the domestication of dogs was an important milestone of human civilization, a study of whole-genome resequencing in dogs and wolves showed that dogs had increased starch digestion function compared to wolves [47]. Our results showed that Prevotella_9 and Peptoclostridium were dominant genera, and some specific bacteria were correlated with physiological indices (such as CHO, T-AOC, IgA, and IFN-α), suggesting that L. acidophilus might enhance lipid metabolism, immunity, and antioxidant performance by modulating the relative abundance of the bacterial community, which makes it easier to adapt to the urban lifestyle of low physical activity.
5. Conclusions
In our study, L. acidophilus plays an important role in improving lipid metabolism, immunity, and anti-oxidation by regulating specific bacterial communities, but the specific correlation and mechanism need to be verified by further research.
Author Contributions
Conceptualization, C.X. and A.M.; methodology, A.M. and W.N.; software, A.M. and W.N.; validation, X.C. and W.Z.; formal analysis, W.Z. and W.N.; investigation, X.C. and W.Z.; resources, C.X.; data curation, Y.H. and Y.S.; writing—original draft preparation, A.M.; writing—review and editing, C.X.; visualization, H.Z.; supervision, C.X. and H.Z.; project administration, C.X.; funding acquisition, C.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Program, grant number 2023YFD1300053.
Institutional Review Board Statement
The animal study protocol was approved by the Animal Ethics Committee of the Laboratory Animal Administration at the Institute of Special Animal and Plant Sciences, Chinese Academy of Agricultural Sciences (Approval NO. ISAPSAEC-2021-60D) for studies involving animals in 24 February 2021.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author, [C.X.], upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gao, H.; Li, X.; Chen, X.; Hai, D.; Wei, C.; Zhang, L.; Li, P. The Functional Roles of Lactobacillus acidophilus in Different Physiological and Pathological Processes. J. Microbiol. Biotechnol. 2022, 32, 1226–1233. [Google Scholar] [CrossRef]
- Mital, B.K.; Garg, S.K. Anticarcinogenic, hypocholesterolemic, and antagonistic activities of Lactobacillus acidophilus. Crit. Rev. Microbiol. 1995, 21, 175–214. [Google Scholar] [CrossRef]
- Theilmann, M.C.; Goh, Y.J.; Nielsen, K.F.; Klaenhammer, T.R.; Barrangou, R.; Abou Hachem, M. Lactobacillus acidophilus Metabolizes Dietary Plant Glucosides and Externalizes Their Bioactive Phytochemicals. mBio 2017, 8, 10–1128. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, B.; Zhou, X.; Wang, Y.; Wang, H.; Jia, S.; Zhang, Z.; Chu, C.; Mu, J. Combined Lowering Effects of Rosuvastatin and L. acidophilus on Cholesterol Levels in Rat. J. Microbiol. Biotechnol. 2019, 29, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Alonso, L.; Fontecha, J.; Cuesta, P. Combined effect of Lactobacillus acidophilus and β-cyclodextrin on serum cholesterol in pigs. Br. J. Nutr. 2016, 115, 1–5. [Google Scholar] [CrossRef]
- Lee, N.Y.; Shin, M.J.; Youn, G.S.; Yoon, S.J.; Choi, Y.R.; Kim, H.S.; Gupta, H.; Han, S.H.; Kim, B.K.; Lee, D.Y.; et al. Lactobacillus attenuates progression of nonalcoholic fatty liver disease by lowering cholesterol and steatosis. Clin. Mol. Hepatol. 2021, 27, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Kang, X.; Yang, H.; Liu, H.; Yang, X.; Liu, Q.; Tian, H.; Xue, Y.; Ren, P.; Kuang, X.; et al. Lactobacillus acidophilus ameliorates obesity in mice through modulation of gut microbiota dysbiosis and intestinal permeability. Pharmacol. Res. 2022, 175, 106020. [Google Scholar] [CrossRef]
- Yan, F.; Li, N.; Shi, J.; Li, H.; Yue, Y.; Jiao, W.; Wang, N.; Song, Y.; Huo, G.; Li, B. Lactobacillus acidophilus alleviates type 2 diabetes by regulating hepatic glucose, lipid metabolism and gut microbiota in mice. Food Funct. 2019, 10, 5804–5815. [Google Scholar] [CrossRef]
- Sandøe, P.; Palmer, C.; Corr, S.; Astrup, A.; Bjørnvad, C.R. Canine and feline obesity: A One Health perspective. Vet. Rec. 2014, 175, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Hamper, B. Current Topics in Canine and Feline Obesity. Vet. Clin. N. Am. Small Anim. Pract. 2016, 46, 785–795. [Google Scholar] [CrossRef]
- Clark, M.; Hoenig, M. Metabolic Effects of Obesity and Its Interaction with Endocrine Diseases. Vet. Clin. N. Am. Small Anim. Pract. 2016, 46, 797–815. [Google Scholar] [CrossRef]
- Coelho, L.P.; Kultima, J.R.; Costea, P.I.; Fournier, C.; Pan, Y.; Czarnecki-Maulden, G.; Hayward, M.R.; Forslund, S.K.; Schmidt, T.S.B.; Descombes, P.; et al. Similarity of the dog and human gut microbiomes in gene content and response to diet. Microbiome 2018, 19, 72. [Google Scholar] [CrossRef]
- Grześkowiak, Ł.; Endo, A.; Beasley, S.; Salminen, S. Microbiota and probiotics in canine and feline welfare. Anaerobe 2015, 34, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Tvarijonaviciute, A.; Barić-Rafaj, R.; Horvatic, A.; Muñoz-Prieto, A.; Guillemin, N.; Lamy, E.; Tumpa, A.; Ceron, J.J.; Martinez-Subiela, S.; Mrljak, V. Identification of changes in serum analytes and possible metabolic pathways associated with canine obesity-related metabolic dysfunction. Vet. J. 2019, 244, 51–59. [Google Scholar] [CrossRef]
- Chikamune, T.; Katamoto, H.; Ohashi, F.; Shimada, Y. Serum lipid and lipoprotein concentrations in obese dogs. J. Vet. Med. Sci. 1995, 57, 595–598. [Google Scholar] [CrossRef]
- Pasupathy, K.; Sahoo, A.; Pathak, N.N. Effect of lactobacillus supplementation on growth and nutrient utilization in mongrel pups. Arch. Tierernahr. 2001, 55, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Baillon, M.-L.A.; Marshall-Jones, Z.V.; Butterwick, R.F. Effects of probiotic Lactobacillus acidophilus strain DSM13241 in healthy adult dogs. Am. J. Vet. Res. 2004, 65, 338–343. [Google Scholar] [CrossRef]
- Marshall-Jones, Z.V.; Baillon, M.L.; Croft, J.M.; Butterwick, R.F. Effects of Lactobacillus acidophilus DSM13241 as a probiotic in healthy adult cats. Am. J. Vet. Res. 2006, 67, 1005–1012. [Google Scholar] [CrossRef]
- Marelli, S.P.; Fusi, E.; Giardini, A.; Martino, P.A.; Polli, M.; Bruni, N.; Rizzi, R. Effects of probiotic Lactobacillus acidophilus D2/CSL (CECT 4529) on the nutritional and health status of boxer dogs. Vet. Rec. 2020, 187, e28. [Google Scholar] [CrossRef]
- National Research Council, Division on Earth, Life Studies, Committee on Animal Nutrition, Subcommittee on Dog, & Cat Nutrition. Research Council. Nutrient Requirements of Dogs and Cats; The National Academies Press: Washington, DC, USA, 2006. [Google Scholar]
- Animal Feeding Stuffs: Methods of Sampling and Analysis—Detection and Enumeration of Lactobacillus spp. Used as Feed Additive. UNE EN 15787:2022. Available online: https://standards.iteh.ai/catalog/standards/cen/cd3bf18a-7ac3-47b2-bc7e-c9571525c94f/en-15787-2021 (accessed on 13 March 2024).
- AOAC. Official Methods of Analysis of the Association of Official’s Analytical Chemists; AOAC: Arlington, VI, USA, 2003. [Google Scholar]
- Berg, J.; Brandt, K.K.; Al-Soud, W.A.; Holm, P.E.; Hansen, L.H.; Sørensen, S.J.; Nybroe, O. Selection for Cu-tolerant bacterial communities with altered composition, but unaltered richness, via long-term Cu exposure. Appl. Environ. Microbiol. 2012, 78, 7438–7446. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Alaqil, A.A.; Abbas, A.O.; El-Beltagi, H.S.; El-Atty, H.K.A.; Mehaisen, G.M.K.; Moustafa, E.S. Dietary Supplementation of Probiotic Lactobacillus acidophilus Modulates Cholesterol Levels, Immune Response, and Productive Performance of Laying Hens. Animals 2020, 10, 1588. [Google Scholar] [CrossRef] [PubMed]
- Roager, H.M.; Sulek, K.; Skov, K.; Frandsen, H.L.; Smedsgaard, J.; Wilcks, A.; Skov, T.H.; Villas-Boas, S.G.; Licht, T.R. Lactobacillus acidophilus NCFM affects vitamin E acetate metabolism and intestinal bile acid signature in monocolonized mice. Gut Microbes 2014, 5, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Crovesy, L.; Ostrowski, M.; Ferreira, D.M.T.P.; Rosado, E.L.; Soares-Mota, M. Effect of Lactobacillus on body weight and body fat in overweight subjects: A systematic review of randomized controlled clinical trials. Int. J. Obes. 2017, 41, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- Liyanage, K.E.; Burnett, J.R.; Hooper, A.J.; van Bockxmeer, F.M. Familial hypercholesterolemia: Epidemiology, Neolithic origins and modern geographic distribution. Crit. Rev. Clin. Lab. Sci. 2011, 48, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Abifadel, M.; Rabès, J.P.; Devillers, M.; Munnich, A.; Erlich, D.; Junien, C.; Varret, M.; Boileau, C. Mutations and polymorphisms in the proprotein convertase subtilisin kexin 9 (PCSK9) gene in cholesterol metabolism and disease. Hum. Mutat. 2009, 30, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Davignon, J.; Dubuc, G.; Seidah, N.G. The influence of PCSK9 polymorphisms on serum low-density lipoprotein cholesterol and risk of atherosclerosis. Curr. Atheroscler. Rep. 2010, 12, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Seidah, N.G.; Awan, Z.; Chrétien, M.; Mbikay, M. PCSK9: A key modulator of cardiovascular health. Circ. Res. 2014, 114, 1022–1036. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Park, S.; Lee, H.; Min, B.; Jung, S.; Park, S.; Kim, E.; Oh, S. Effect of Lactobacillus acidophilus NS1 on plasma cholesterol levels in diet-induced obese mice. J. Dairy Sci. 2015, 98, 1492–1501. [Google Scholar] [CrossRef]
- Park, S.S.; Lee, Y.J.; Song, S.; Kim, B.; Kang, H.; Oh, S.; Kim, E. Lactobacillus acidophilus NS1 attenuates diet-induced obesity and fatty liver. J. Endocrinol. 2018, 237, 87–100. [Google Scholar] [CrossRef]
- Janda, A.; Bowen, A.; Greenspan, N.S.; Casadevall, A. Ig Constant Region Effects on Variable Region Structure and Function. Front. Microbiol. 2016, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.A. Canine IgA and IgA deficiency: Implications for immunization against respiratory pathogens. Can. Vet. J. 2019, 60, 1305–1311. [Google Scholar] [PubMed]
- Maurer, M.A.; Meyer, L.; Bianchi, M.; Turner, H.L.; Le, N.P.L.; Steck, M.; Wyrzucki, A.; Orlowski, V.; Ward, A.B.; Crispin, M.; et al. Glycosylation of Human IgA Directly Inhibits Influenza A and Other Sialic-Acid-Binding Viruses. Cell Rep. 2018, 23, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Herr, A.B.; Ballister, E.R.; Bjorkman, P.J. Insights into IgA-mediated immune responses from the crystal structures of human FcalphaRI and its complex with IgA1-Fc. Nature 2003, 423, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Breedveld, A.; van Egmond, M. IgA and FcαRI: Pathological Roles and Therapeutic Opportunities. Front. Immunol. 2019, 10, 553. [Google Scholar] [CrossRef]
- De Sousa-Pereira, P.; Woof, J.M. IgA: Structure, Function, and Developability. Antibodies 2019, 8, 57. [Google Scholar] [CrossRef]
- Mantis, N.J.; Rol, N.; Corthésy, B. Secretory IgA’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef]
- Kwaa, A.K.R.; Talana, C.A.G.; Blankson, J.N. Interferon Alpha Enhances NK Cell Function and the Suppressive Capacity of HIV-Specific CD8+ T Cells. J. Virol. 2019, 93, e01541-18. [Google Scholar] [CrossRef]
- Malek, T.R. The main function of IL-2 is to promote the development of T regulatory cells. J. Leukoc. Biol. 2003, 74, 961–965. [Google Scholar] [CrossRef]
- Chen, L.; Liu, W.; Li, Y.; Luo, S.; Liu, Q.; Zhong, Y.; Jian, Z.; Bao, M. Lactobacillus acidophilus ATCC 4356 attenuates the atherosclerotic progression through modulation of oxidative stress and inflammatory process. Int. Immunopharmacol. 2013, 17, 108–115. [Google Scholar] [CrossRef]
- Ghiselli, A.; Serafini, M.; Natella, F.; Scaccini, C. Total antioxidant capacity as a tool to assess redox status: Critical view and experimental data. Free Radic. Biol. Med. 2000, 29, 1106–1114. [Google Scholar] [CrossRef] [PubMed]
- Handl, S.; Dowd, S.E.; Garcia-Mazcorro, J.F.; Steiner, J.M.; Suchodolski, J.S. Massive parallel 16S rRNA gene pyrosequencing reveals highly diverse fecal bacterial and fungal communities in healthy dogs and cats. FEMS Microbiol. Ecol. 2011, 76, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Suchodolski, J.S. Companion animals symposium: Microbes and gastrointestinal health of dogs and cats. J. Anim. Sci. 2011, 89, 1520–1530. [Google Scholar] [CrossRef] [PubMed]
- Axelsson, E.; Ratnakumar, A.; Arendt, M.L.; Maqbool, K.; Webster, M.T.; Perloski, M.; Liberg, O.; Arnemo, J.M.; Hedhammar, A.; Lindblad-Toh, K. The genomic signature of dog domestication reveals adaptation to a starch-rich diet. Nature 2013, 495, 360–364. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).