1. Introduction
The emergence of antibiotics played an important role in animal growth and reproduction, disease prevention and treatment, etc. However, with the excessive abuse of antibiotics, bacteria in animals have developed resistance, disrupting the balance of animal gastrointestinal microbiota and, most importantly, it can also cause serious harm to human health [
1,
2]. China is a major country of the world’s livestock and poultry farming, as well as a major producer and user of antibiotics. In order to produce safe and high-quality livestock and poultry products and achieve healthy breeding, researchers urgently need to find ideal substitutes for antibiotic feed additives. Probiotics have the advantages of enhancing animal immunity and disease resistance, improving animal production performance and feed utilization, and are an ideal alternative to antibiotics [
3,
4,
5]. The impact mechanism of probiotics on animals is mainly manifested through two aspects: regulating gastrointestinal health and improving the body’s immunity [
6]. Probiotic additives can accelerate the growth and development of immune organs in ruminants by regulating the structure, quantity, and fermentation level of the gastrointestinal microbiota [
7,
8,
9].
Animal probiotics, as one of the raw materials for microbial preparations, are currently mainly used in feed additives and silage [
10,
11]. Although probiotic products currently cannot completely replace antibiotic additives in diets, their application scope has gradually expanded and their reliability has improved since use began [
12,
13]. Probiotics have been shown to enhance the gut microbiota of livestock and poultry, and enhance immunity and growth performance [
14,
15,
16]. At present, research on probiotics is very limited, mainly focusing on monogastrics, and there is still less research on ruminants, which to some extent limits the effective application of probiotics [
17]. The development of multi-omics technology provides the possibility for a deeper understanding of the effects and mechanisms of probiotics.
The main probiotics used in ruminant feed include
Lactobacillus,
Aspergillus oryzae,
Streptococcus, and
Bifidobacterium [
6,
17,
18]. Probiotic preparations mainly adjust the microbiota balance in an animal’s gastrointestinal tract, serving as the dominant microbiota in the animal gut to compensate for the number of normal microbiota, promote the formation of intestinal dominant microbiota, inhibit the growth and reproduction of pathogenic microorganisms, and restore the balance of gastrointestinal microbiota [
19,
20]. According to existing research findings, probiotics are defined as live microbial feed additives that beneficially affect host animals by enhancing intestinal balance, thereby improving feed utilization, nutrient absorption, growth rate, and economic benefits of poultry [
21,
22,
23]. Preliminary experimental studies have found that the
Bacillus licheniformis group (
Bacillus licheniformis),
Bacillus subtilis binary group (
Bacillus subtilis and
Enterococcus faecalis),
Bifidobacterium triple group (
Bifidobacterium longum,
Lactobacillus bulgaricus, and
Streptococcus thermophilus) and
Bifidobacterium tetravalent group (
Bifidobacterium,
Lactobacillus acidophilus,
Enterococcus faecalis, and
Bacillus cereus) all improved the growth performance of fattening sheep, with the
Bifidobacterium tetravalent group having the most significant effect (
p < 0.05). We chose
Bifidobacterium tetravalent for this experiment (
Supplementary Table S1). Our research focuses on demonstrating the mechanism by which adding probiotics to the diet enhances the growth performance of fattening sheep. From multiple perspectives, including rumen microbiota, fecal microbiota, rumen metabolomics, serum metabolomics, urine metabolomics, and transcriptome sequencing of rumen epithelial cells, this will provide scientific guidance for the study of probiotics additives in ruminant feed.
2. Materials and Methods
2.1. Experimental Animals and Design
Twelve healthy Hu sheep (of similar age and weight) from Zhoushi Sheep Industry Co., Ltd. in Chuzhou, China, were selected for this experiment and randomly divided into two groups. The weight range of the fattening sheep in each group was 22.6 ± 1.9 kg. The control group (CON) was fed with a basic diet, and the formula and chemical composition are shown in
Table 1. The experimental group (PRB) added probiotics (bifidobacteria quadruplex live bacteria:
Bifidobacterium,
Lactobacillus acidophilus,
Enterococcus faecalis, and
Bacillus cereus) to the control group’s diet, with a prefeeding period of 2 weeks and a formal feeding period of 2 months. During the experiment, each group was fed at a fixed time in the morning and evening, and the fattening sheep in each group fed and drank freely. Other treatments were carried out according to the daily feeding and management methods of the farm. The experimental plan was approved by the Animal Protection Committee of Anhui Agricultural University (NO: SYDW-P20190600601), and the experimental design and workflow are shown in
Figure 1.
2.2. Test Samples and Data Collection
On the 1st and 60th day of the formal experiment, each group of experimental sheep was weighed, with an empty stomach before feeding, to calculate the average daily gain. The feed consumption of each group of fattening sheep during the 2-month experimental period was calculated to calculate the average daily feed intake. The feed conversion rate was calculated based on the average daily gain/average daily feed intake.
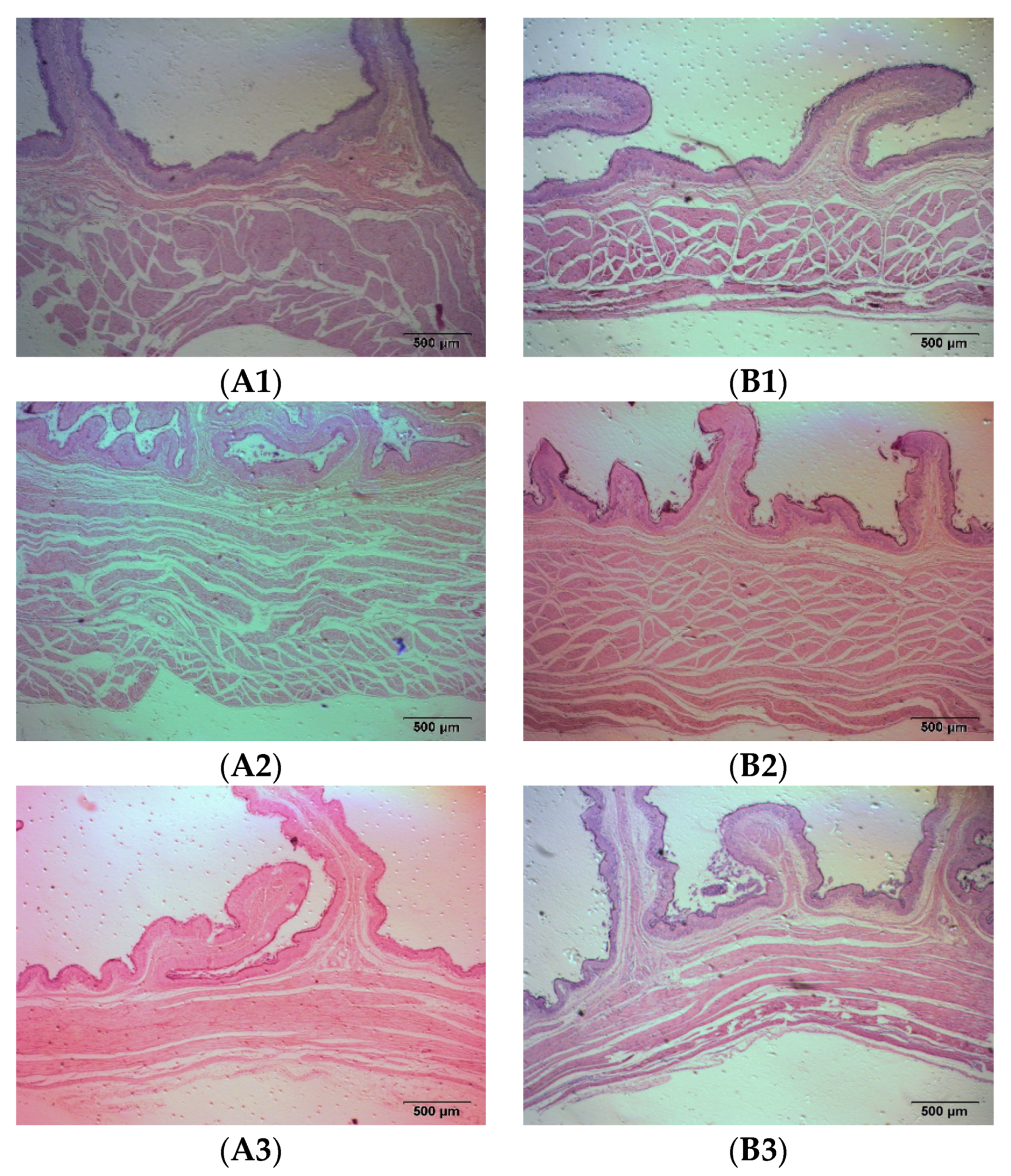
On the 60th day of the formal experiment, before feeding in the morning, the rumen fluid of each group was collected using a sheep stomach catheter. After filtering through 4 layers of gauze, it was quickly loaded into a 50 mL centrifuge tube, and centrifuged at 3000 r/min for 15 min. An amount of 1 mL was taken for analysis of the volatile fatty acids (VFAs), 2 mL was taken for a colorimetric analysis of the ammonia nitrogen content, and the remainder was stored at −80 °C for later use. An amount of 100 g of fresh fecal samples from each group of fattening sheep were collected, and store in liquid nitrogen for later use. Fasting blood was collected using a blood collection vessel without anticoagulants, and 10 mL of blood was collected from each Hu sheep and centrifuged at 3500 r/min for 15 min. The supernatant was used as the serum. The serum antioxidant indicators were analyzed using a reagent kit, including the total antioxidant capacity (T-AOC) (A015-1-2, Nanjing Institute of Biotechnology, Nanjing, China), superoxide dismutase (SOD) (A001-3-2, Nanjing Institute of Biotechnology, Nanjing, China), glutathione peroxidase (GSH-Px) (A005-1-2, Nanjing Jiancheng Institute of Biotechnology, Nanjing, China), and malondialdehyde (MDA) content (A003-1-2, Nanjing Jiancheng Institute of Biotechnology, Nanjing, China). The remaining was stored at −80 °C for future use. Fresh urine from the fattening sheep was collected and stored at −80 °C for future use. After the feeding experiment, all 12 fattening sheep in the CON and PRB groups were slaughtered (fasted for 12 h before slaughter), and the rumen epithelial tissue was collected and stored in liquid nitrogen for transcriptome sequencing. In addition, the intestinal tissue of the fattened sheep was collected, fixed with 4% paraformaldehyde solution, embedded in paraffin, and sliced. After HE staining, the morphological changes in each segment of intestinal tissue were observed.
2.3. Microbial Analysis of Rumen Fluid and Feces
Using a DNA extraction kit to extract microbial DNA from the rumen fluid and feces, 16S rRNA gene V3-V4 region universal primers were used to amplify the target sequence using a two-step PCR method. A Qubit 4.0 was used to measure the concentration of the extracted DNA and amplified sequence. Bioengineering (Shanghai, China) Co., Ltd. We used an Illumina MiSeq system for high-throughput sequencing, and analyzed the differences in microbial diversity and microbial community structure in the rumen and feces of fattening sheep between the CON and PRB groups.
2.4. Metabolomics Analysis of Rumen, Serum, and Urine
An amount of 10 μL of the previously preserved rumen fluid, serum, and urine samples were taken, and ultra-high-performance liquid chromatography–tandem quadrupole–time-of-flight mass spectrometry (UPLC-Q-TOF-MS) was used to perform a metabolomics analysis of the rumen fluid, serum, and urine of the fattening sheep. After processing and quality control of the obtained raw data, a multivariate statistical analysis was conducted; the conditions were that the p-value of Student’s t-test was less than 0.05 and the Variable Importance in the Projection (VIP) of the first principal component of the OPLS-DA model was greater than 1. Differential metabolites in the rumen fluid, serum, and urine were screened, and a metabolic pathway analysis was conducted to identify the key pathways with the highest correlation with the differential metabolites, and we analyzed the differences in the metabolites and metabolic pathways between the CON group and the PRB group of fattening sheep.
2.5. Transcriptomics Analysis
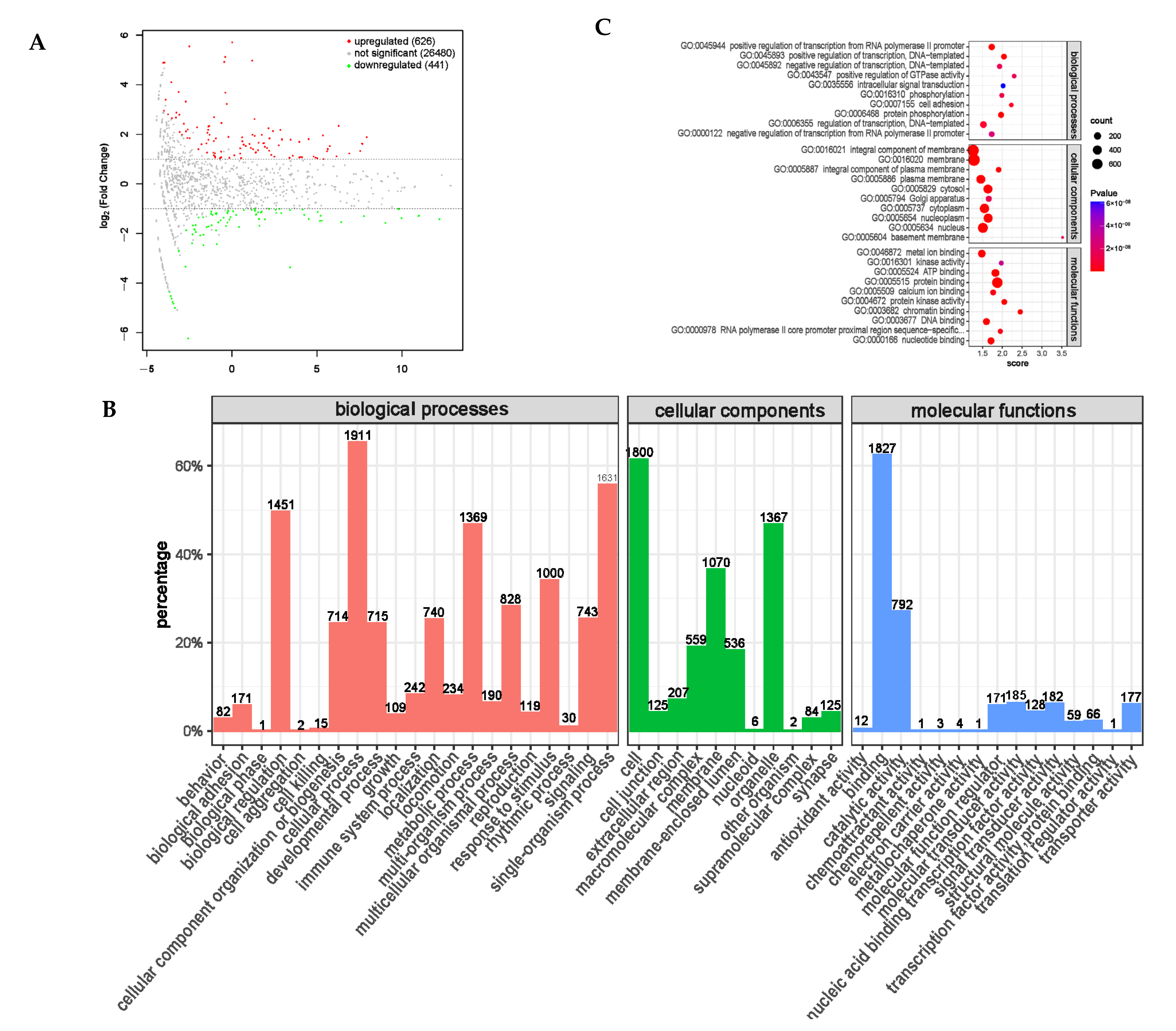
The RNA extraction and transcriptome sequencing of the rumen epithelial tissue samples were entrusted to Bioengineering (Shanghai, China) Co., Ltd. After RNA extraction, the samples were sequenced on the Illumina platform. After sequencing, data processing, and quality control, a differential expression analysis was performed using the expression level difference analysis software DESeq 2. Using a p-value < 0.05 and fold of difference |Fold Change | > 2 as the screening criteria, the differential expression of genes (DEGs) between the groups were screened. A comparative analysis was conducted between the DEGs and GO (Gene Ontology) databases to obtain the functional annotation information corresponding to the genes. Cluster Profiler was used for the functional enrichment analysis. When the p-value < 0.05, it was considered that the function was enriched, and the function of the differentially expressed genes was analyzed.
2.6. Data Analysis
The normal distribution of the data was tested using the SPSSAU Data Science Analysis Platform (
https://spssau.com/, accessed on 6 February 2024). The Shapiro–Wilk test was used for normality testing. The experimental sample consisted of 6 replicates. In order to organize the data, Excel 2016 was used, and SPSS software (version 20.0, Chicago, IL, USA) was used to conduct independent sample t-tests on the data.
p < 0.05 is the significant difference, and
p < 0.01 is the extremely significant difference standard. The difference between the two levels is statistically significant, with
p < 0.05 and
p < 0.01, respectively.
4. Discussion
Research has shown that probiotics can play an important role as the best alternative to antibiotics, mainly through the interaction of bacteria–bacteria and host–bacteria [
10]. They have many beneficial aspects, such as anti-oxidation, anti-inflammatory, anti-allergy, anti-cancer, anti-mutation, anti-diabetes, and anti-virus potential, and they can also be used as a growth promoter for animals [
31,
32,
33,
34]. Probiotics are defined as live microbial feed additives that beneficially affect host animals by enhancing intestinal balance, improving feed efficiency, nutrient absorption, and growth rate [
35]. Research has found that probiotics can be used as useful bacteria with which to eliminate pathogenic bacteria, and their potential applications can help improve the growth performance of livestock and poultry, improve the digestion and absorption capacity of nutrients, promote production, and ensure the health of livestock [
36,
37,
38]. In this study, compared with the CON group, the PRB group was fed 1.5 × 10
8 CFU/g probiotics daily, and the average daily weight gain of the PRB group was significantly higher than that of the CON group (
p < 0.05), indicating that the addition of probiotics significantly improved the growth performance of the fattening sheep, which is consistent with previous research results showing that probiotics improve livestock and poultry growth performance.
The main characteristic of ruminants is the fermentation of microorganisms in the rumen, and the rumen volume accounts for 80% of the entire stomach volume [
39]. There are a large number of ciliates and bacteria in the rumen, which play an important role in the rumen fermentation process. Research has found that probiotics can regulate gastrointestinal microbiota and promote digestion [
40,
41]. Feeding probiotics improves the composition of the rumen microbiota in fattening sheep, enhances microbial fermentation, and facilitates the absorption and utilization of cellulose and hemicellulose into volatile fatty acids, promoting protein decomposition and synthesis [
42]. The rumen muscle layer thickness of the PRB fattening sheep was significantly lower than that of the CON (
p < 0.01), due to the enhanced digestion of microorganisms in the rumen, which does not require strong peristalsis of food in the rumen, resulting in a thinning of the rumen muscle layer thickness. At the same time, it reduces the energy consumption of rumen peristalsis, and this experiment confirms that supplementing with probiotics improves the composition of rumen microorganisms and improves the digestion efficiency of fattening sheep feed.
The intestine is an important site for the digestion and absorption of nutrients, and the efficient digestion and absorption of feed nutrients plays an important role in the healthy growth of livestock [
43]. The intestinal villi on the surface of the duodenal intima increase the surface area of the intestine, allowing for more nutrients to be absorbed. In addition, the cells on the surface of the intestinal villi secrete more digestive enzymes, enabling efficient digestion and the absorption of feed nutrients [
44]. In this experiment, the length of the duodenal villi in the PRB group was significantly higher than that in the CON group (
p < 0.05), indicating that adding probiotics helps improve the digestion and absorption of nutrients in fattening sheep. The peristalsis of the intestine mainly relies on the contraction ability of the muscular layer. The developed muscular layer can promote intestinal peristalsis and enhance the absorption and utilization of nutrients. The PRB group had no significant difference except for the mucosal muscular layer of the colon (
p > 0.05), while the mucosal layer and muscular layer of the cecum, colon, and rectum significantly increased, confirming that the PRB group were more efficiency at nutrient absorption and utilization (
p < 0.05).
In this study, supplementing with probiotics significantly increased the total volatile fatty acid concentration (
p < 0.01), indicating that probiotics have an improved effect on rumen microbial fermentation. The increase in total volatile fatty acid level indicates a high energy supply level for fattening sheep, and the average daily weight gain of the PRB group of fattening sheep significantly increased. The gastrointestinal microbiota is a signaling hub that links the host’s metabolism and immune system. Adding probiotics to the diet helps to establish and maintain suitable microbiota in the gastrointestinal tract [
45].
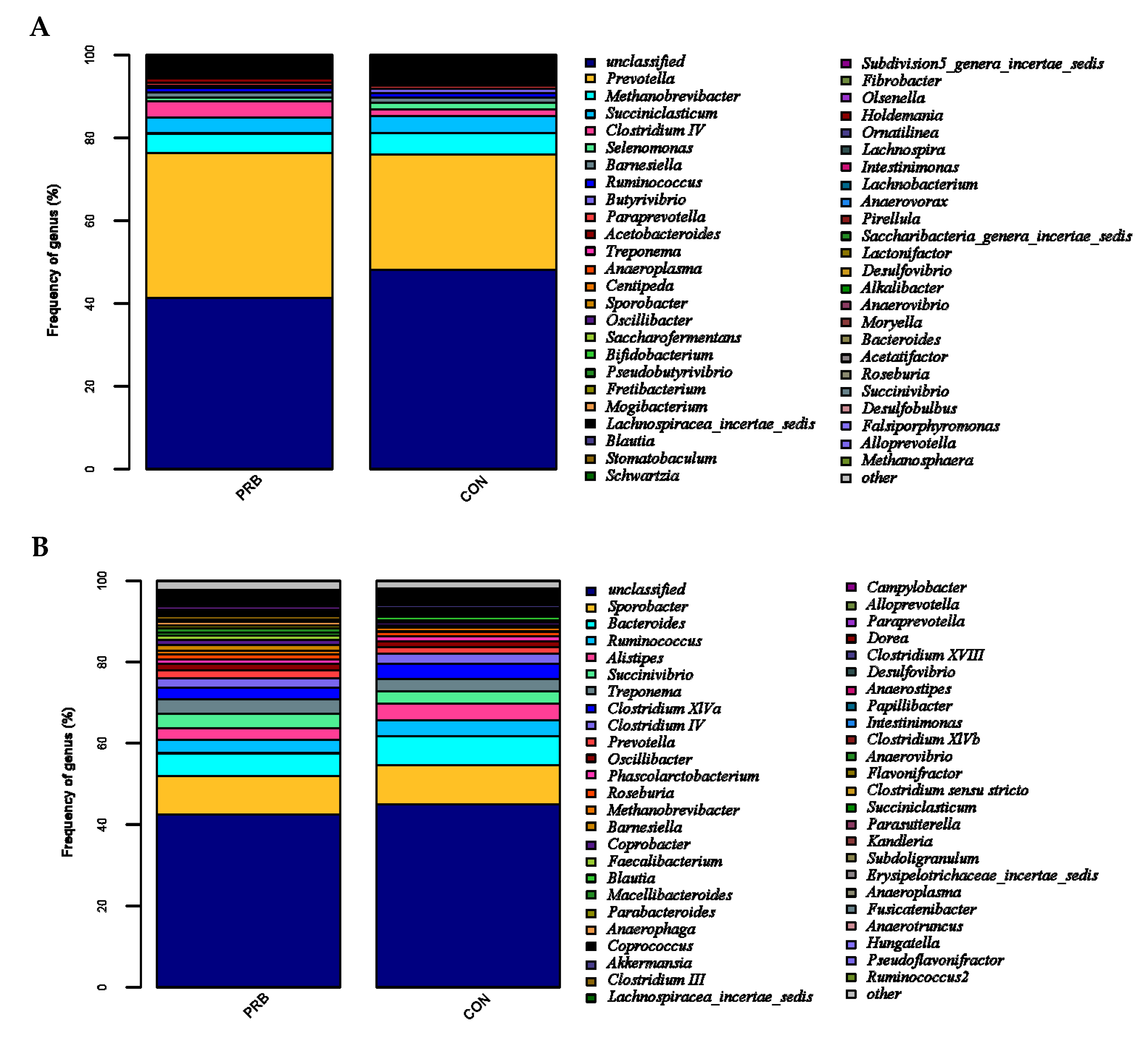
A rumen microbiota analysis revealed a significant difference between the PRB group and the CON group in their total of five bacterial genera (
p < 0.05). Compared to the CON group, the PRB group significantly upregulated Subdivision5_genera_incertae_sedis and Acinetobacter, and significantly downregulated
Butyrivibrio,
Saccharofermentans, and
Fibrobacter. The rumen, as a unique digestive organ of ruminants, plays an important role. Improving the composition of rumen microbiota by supplementing with probiotics is of great significance for improving the growth performance of fattening sheep [
46]. Probiotics have been shown to have various beneficial physiological effects on animal intestines. An analysis of fecal microorganisms revealed that the significantly upregulated genus in the PRB group was
Faecalicoccus, and the significantly downregulated genera included
Coprococcus, Porphyromonas, and
Anaerobacterium. Supplementing with probiotics in the diet is beneficial for promoting the reproduction of intestinal microbiota, maintaining the balance of intestinal microbiota, increasing intestinal patency, enhancing intestinal protective function, regulating the body’s metabolism, promoting the growth and development of fattening sheep, and enhancing immunity.
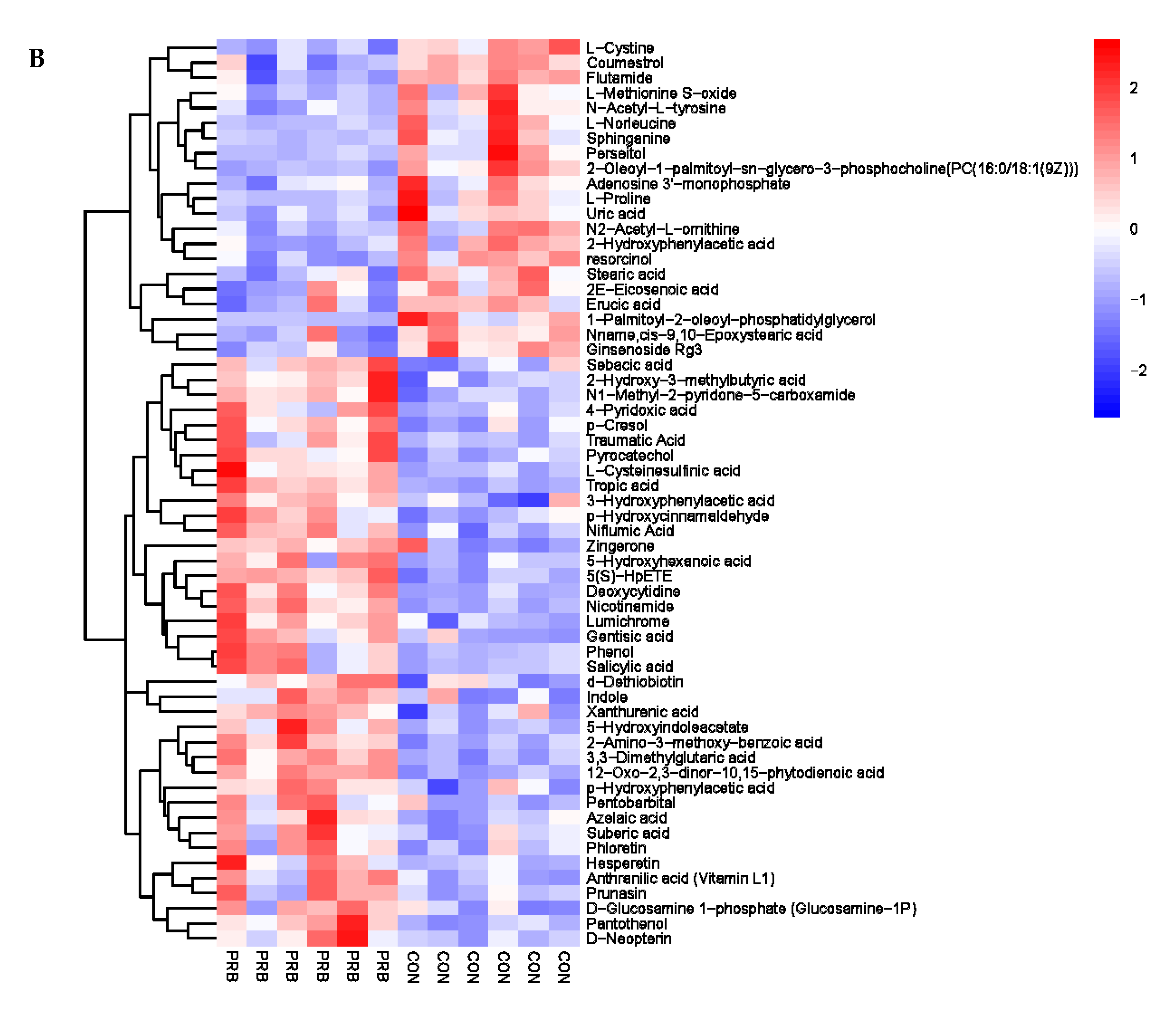
To further explore the mechanism of adding probiotics to the diet to enhance the growth performance of fattening sheep, we conducted a metabolomics analysis on the rumen fluid, serum, and urine. Significant changes in metabolites were observed in the rumen fluid, serum, and urine of fattening sheep supplemented with probiotics. The pyruvate in serum differential metabolites is mainly involved in the entire basic metabolic process through the acetyl-CoA and tricarboxylic acid cycle pathways, including the conversion between sugars, fats, and amino acids [
47]. These differential metabolites play an important role in energy metabolism and amino acid metabolism.
An analysis of the key metabolic pathways found that supplementing with probiotics had an impact on taurine and taurine metabolic pathways, vitamin B6 and vitamin B2, and a series of amino acid metabolism pathways. Taurine is a widely present free amino acid that participates in many different physiological processes, such as bile acid binding, osmotic regulation, antioxidant activity, calcium signaling regulation, and liver detoxification. It is the main biosynthetic compound of the liver and can improve gastric injury through its antioxidant properties [
48,
49]. Vitamin B6 plays a role in the various metabolic functions of proteins, carbohydrates, and lipids. At the same time, it can also participate in the conversion of tryptophan into niacin or 5-hydroxytryptamine, which can enhance immunity [
50]. Vitamin B2 is involved in the tricarboxylic acid cycle and the catalytic action of multiple enzymes in the respiratory chain, promoting glucose, fat, and protein metabolism, thereby generating energy. Its metabolites FAD and FMN can serve as coenzymes that participate in electron transfer and ATP synthesis in the mitochondrial respiratory chain, maintaining the normal respiratory function of cells. It can also serve as an antioxidant, protecting cells from free radical damage and slowing down the aging process [
51]. The results of the metabolomics analysis suggest that probiotics can play a role in promoting growth and development by regulating energy and amino acid metabolism pathways.
In addition, we screened differential genes using a GO functional enrichment analysis. In the PRB and CON groups, we screened target genes with a significant differential expression of circRNA, lncRNA, mRNA, and miRNA, which are mainly related to cellular processes, biological regulation, and metabolic processes. They are mainly enriched in protein modification regulation (especially histone modification), immune function regulation, and energy metabolism pathways, which play important roles in animal growth and development.
Previous studies have found that adding probiotics to livestock and poultry feed can improve the digestive system’s beneficial microbiota, increase the feed digestion and absorption rate, and regulate the body’s immune system [
52,
53,
54]. These studies are consistent with the mechanism of adding probiotics to improve the growth performance of fattening sheep in this experiment, supporting the finding that supplementing with probiotics can improve the growth performance of fattening sheep. In the future, we will conduct in-depth research on the mechanisms of probiotics in healthy aquaculture, laying a solid theoretical foundation for the widespread application of probiotics in the field of animal husbandry.

 ,
, 


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}