
Models and Methods to Investigate Acute Stress Responses in Cattle

Abstract
:1. Introduction
Defining Stress
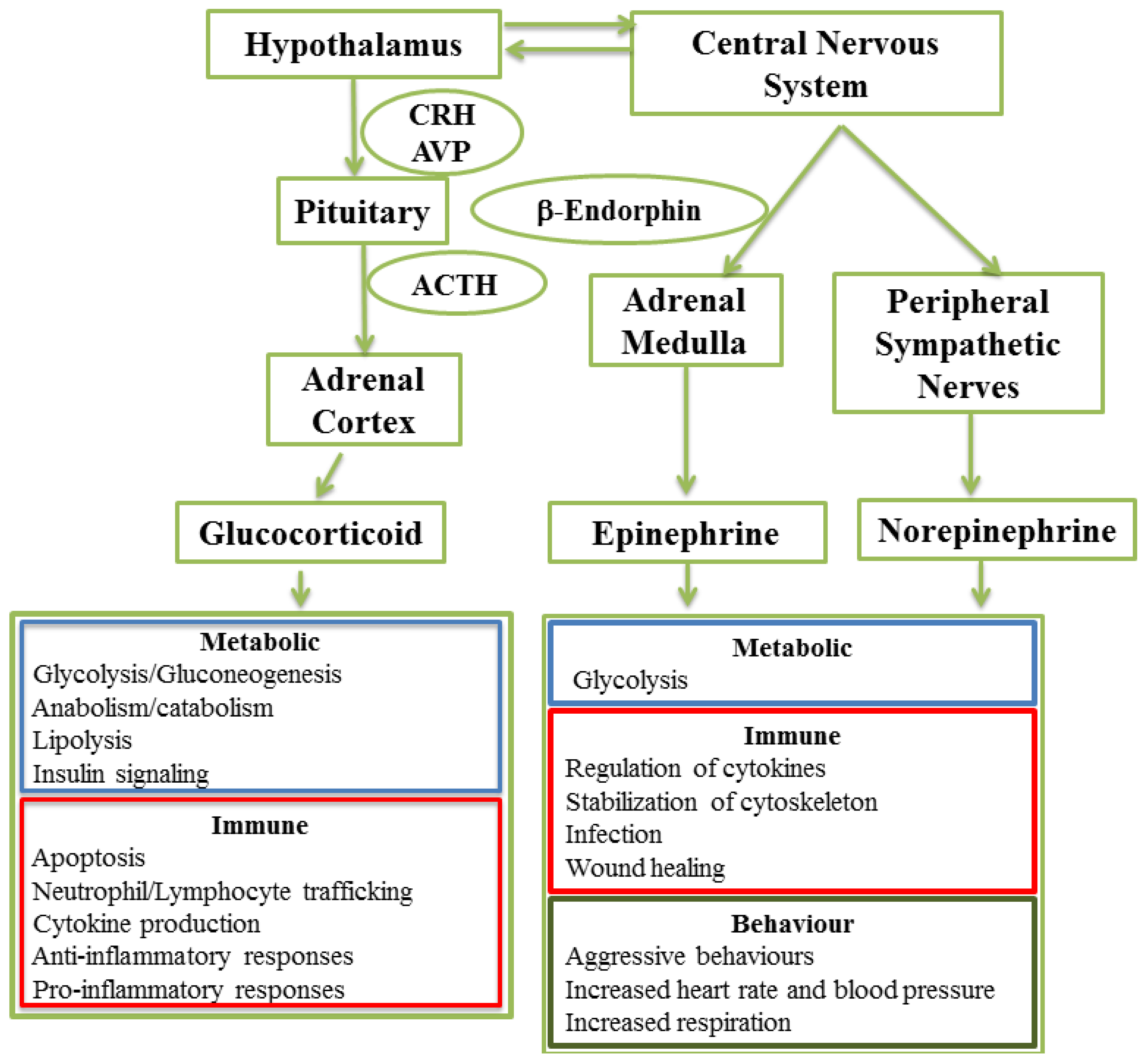
2. The Stress Response
2.1. Glucocorticoid-Induced Signaling
2.2. Metabolic Functions Activated by Glucocorticoid-Induced Signaling

2.3. Immune Responses Regulated by Glucocorticoid-Induced Signaling
2.4. Catecholamine Signaling
2.5. Metabolic Functions Activated by Catecholamine-Induced Signaling
2.6. Immune Responses Regulated by Catecholamine-Induced Signaling
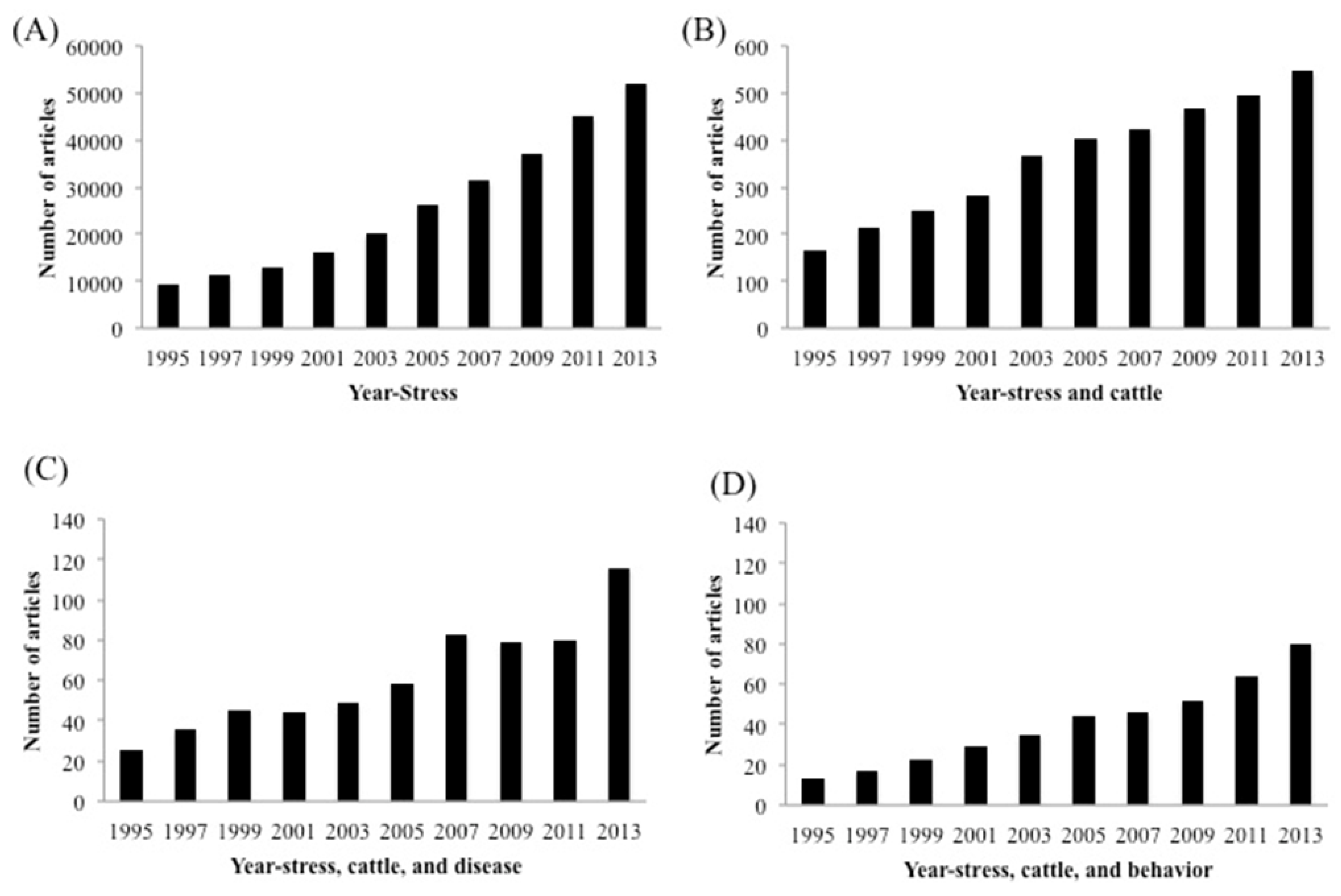
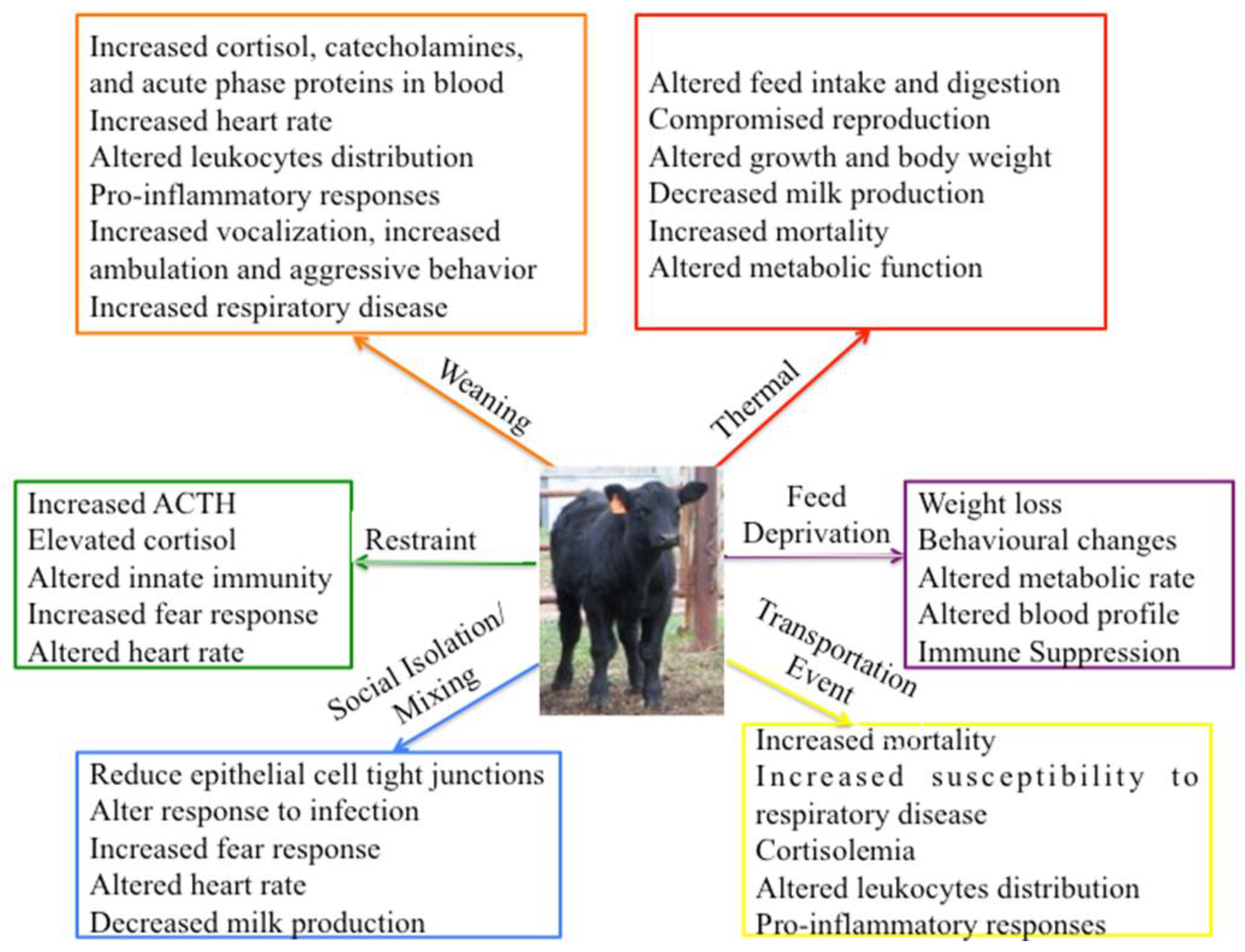
3. Stressors Studied in Cattle


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stressor | Cattle | Pigs | Hens | Sheep | Duck | Horse | Mice | Rats |
|---|---|---|---|---|---|---|---|---|
| Physical stressors | ||||||||
| Thermal Stressors | ||||||||
| Cold | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ |
| Heat | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ |
| Transportation | ✔ | ✔ | ✔ | ✔ | ✗ | ✔ | ✔ | ✔ |
| Feed Deprivation/restriction | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ |
| Noise | ✔ | ✔ | ✔ | ✗ | ✗ | ✗ | ✔ | ✔ |
| Psychological stressors | ||||||||
| Weaning | ✔ | ✔ | ✗ | ✔ | ✗ | ✔ | ✔ | ✔ |
| Social isolation/mixing | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ |
| Restraint | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ |

3.1. Physical Stressors
3.1.1. Thermal Stress
3.1.2. Transportation Stress
3.1.3. Feed Deprivation
3.2. Psychological Stressors
3.2.1. Maternal Separation and Weaning
3.2.2. Social Isolation and Mixing
3.2.3. Restraint Stress
3.2.4. Stress and Disease
4. Measuring Stress Responses
4.1. Measurement of Behavioral Responses to Stress
4.2. Measuring Physiological Responses to Stress
| Physiological Responses to Stress | Reference |
|---|---|
| Cortisol Measurement | |
| Serum/Plasma | [74] |
| Urine | [85] |
| Salivary | [86] |
| Hair | [80] |
| Milk | [87] |
| Epinephrine | [54] |
| Norepinephrine | [54] |
| Faecal metabolites | [88] |
| White Blood Cell Counts | |
| Complete Blood Count | [45] |
| Neutrophil/Lymphocyte | [45] |
| Heart rate | [66] |
| Respiratory rate | [66] |
| Body Temperature | [14] |
| Glucose | [89] |
| Pyruvate | [15] |
| Acute Phase Proteins | [77] |
| Innate Immune Responses | [90] |
| Body Weight | [77] |
| Serum proteome | [8] |
| Transcriptional Profiles | [14] |
5. Immunometabolism

6. Implications
6.1. Changing Approaches to Animal Agriculture
6.2. Translation to Human Health
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Goldstein, D.S.; Kopin, I.J. Evolution of concepts of stress. Stress 2007, 10, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Cannon, W.B. Bodily Changes in Pain, Hunger, Fear, and Rage: An Account of Recent Researches into the Function of Emotional Excitement; Appleton and Company: New York, NY, USA, 1929. [Google Scholar]
- Selye, H. The evolution of the stress concept. Am. Sci. 1973, 61, 692–699. [Google Scholar] [CrossRef]
- Baldwin, D.V. Primitive mechanisms of trauma response: An evolutionary perspective on trauma-related disorders. Neurosci. Biobehav. Rev. 2013, 37, 1549–1566. [Google Scholar] [CrossRef] [PubMed]
- Gallup, G.G. Tonic immobility: The role of fear and predation. Psychol. Rec. 1977, 27, 41–61. [Google Scholar]
- Minton, J.E. Function of the hypothalamic-pituitary-adrenal axis and the sympathetic nervous system in models of acute stress in domestic farm animals. J. Anim. Sci. 1994, 72, 1891–1898. [Google Scholar] [PubMed]
- Mormede, P.; Andanson, S.; Auperin, B.; Beerda, B.; Guemene, D.; Malmkvist, J.; Manteca, X.; Manteuffel, G.; Prunet, P.; van Reenen, C.G.; et al. Exploration of the hypothalamic-pituitary-adrenal function as a tool to evaluate animal welfare. Physiol. Behav. 2007, 92, 317–339. [Google Scholar] [CrossRef] [PubMed]
- Aich, P.; Jalal, S.; Czuba, C.; Schatte, G.; Herzog, K.; Olson, D.J.; Ross, A.R.; Potter, A.A.; Babiuk, L.A.; Griebel, P. Comparative approaches to the investigation of responses to stress and viral infection in cattle. OMICS 2007, 11, 413–434. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.L.; Tank, A.W. Stress-induced catecholaminergic function: Transcriptional and post-transcriptional control. Stress 2007, 10, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.J.; Webb, H.E.; Zourdos, M.C.; Acevedo, E.O. Cardiovascular reactivity, stress, and physical activity. Front. Physiol. 2013, 4, 314. [Google Scholar] [CrossRef] [PubMed]
- Prager, E.M.; Johnson, L.R. Stress at the synapse: Signal transduction mechanisms of adrenal steroids at neuronal membranes. Sci. Signal. 2009. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Topete, D.; Cidlowski, J.A. One hormone, two actions: Anti- and pro-inflammatory effects of glucocorticoids. Neuroimmunomodulation 2015, 22, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Liberman, A.C.; Antunica-Noguerol, M.; Arzt, E. Modulation of the glucocorticoid receptor activity by post-translational modifications. Nucl. Recept. Res. 2014, 1, 1–15. [Google Scholar] [CrossRef]
- Kolli, V.; Upadhyay, R.C.; Singh, D. Peripheral blood leukocytes transcriptomic signature highlights the altered metabolic pathways by heat stress in zebu cattle. Res. Vet. Sci. 2014, 96, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Mehla, K.; Magotra, A.; Choudhary, J.; Singh, A.K.; Mohanty, A.K.; Upadhyay, R.C.; Srinivasan, S.; Gupta, P.; Choudhary, N.; Antony, B.; et al. Genome-wide analysis of the heat stress response in Zebu (Sahiwal) cattle. Gene 2014, 533, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Revollo, J.R.; Cidlowski, J.A. Mechanisms generating diversity in glucocorticoid receptor signaling. Ann. N. Y. Acad. Sci. 2009, 1179, 167–178. [Google Scholar] [CrossRef] [PubMed]
- De Guia, R.M.; Rose, A.J.; Herzig, S. Glucocorticoid hormones and energy homeostasis. Horm. Mol. Biol. Clin. Investig. 2014, 19, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, P.D.; Aich, P.; Stookey, J.; Popowych, Y.; Potter, A.; Babiuk, L.; Griebel, P.J. Stress significantly increases mortality following a secondary bacterial respiratory infection. Vet. Res. 2012, 43, 21. [Google Scholar] [CrossRef] [PubMed]
- Hermoso, M.A.; Matsuguchi, T.; Smoak, K.; Cidlowski, J.A. Glucocorticoids and tumor necrosis factor alpha cooperatively regulate toll-like receptor 2 gene expression. Mol. Cell. Biol. 2004, 24, 4743–4756. [Google Scholar] [CrossRef] [PubMed]
- Griebel, P.; Hill, K.; Stookey, J. How stress alters immune responses during respiratory infection. Anim. Health Res. Rev. 2014, 15, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Chinenov, Y.; Rogatsky, I. Glucocorticoids and the innate immune system: Crosstalk with the toll-like receptor signaling network. Mol. Cell. Endocrinol. 2007, 275, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Ayroldi, E.; Cannarile, L.; Migliorati, G.; Nocentini, G.; Delfino, D.V.; Riccardi, C. Mechanisms of the anti-inflammatory effects of glucocorticoids: Genomic and nongenomic interference with MAPK signaling pathways. FASEB J. 2012, 26, 4805–4820. [Google Scholar] [CrossRef] [PubMed]
- Webster Marketon, J.I.; Glaser, R. Stress hormones and immune function. Cell. Immunol. 2008, 252, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Burns, V. Stress and antibody response to vaccination: Implications of animal studies for human clinical research. Expert Rev. Vaccines 2004, 3, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Padgett, D.A.; Glaser, R. How stress influences the immune response. Trends Immunol. 2003, 24, 444–448. [Google Scholar] [CrossRef]
- Ziegler, M.G.; Elayan, H.; Milic, M.; Sun, P.; Gharaibeh, M. Epinephrine and the metabolic syndrome. Curr. Hypertens. Rep. 2012, 14, 1–7. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, I.A.; Bennett, T.; Fellows, I.W. Catecholamines and the control of metabolism in man. Clin. Sci. 1985, 68, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Vara, E.; Tamarit-Rodriguez, J. Does cyclic guanosine monophosphate mediate noradrenaline-induced inhibition of islet insulin secretion stimulated by glucose and palmitate? Biochem. J. 1991, 278, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.D.; Qiu, Y.; Cui, X.; Goh, Y.P.; Mwangi, J.; David, T.; Mukundan, L.; Brombacher, F.; Locksley, R.M.; Chawla, A. Alternatively activated macrophages produce catecholamines to sustain adaptive thermogenesis. Nature 2011, 480, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Stojadinovic, O.; Gordon, K.A.; Lebrun, E.; Tomic-Canic, M. Stress-induced hormones cortisol and epinephrine impair wound epithelization. Adv. Wound Care 2012, 1, 29–35. [Google Scholar]
- Kim, M.H.; Gorouhi, F.; Ramirez, S.; Granick, J.L.; Byrne, B.A.; Soulika, A.M.; Simon, S.I.; Isseroff, R.R. Catecholamine stress alters neutrophil trafficking and impairs wound healing by beta2-adrenergic receptor-mediated upregulation of IL-6. J. Invest. Ermatol. 2014, 134, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Szelenyi, J.; Vizi, E.S. The catecholamine cytokine balance: Interaction between the brain and the immune system. Ann. N. Y. Acad. Sci. 2007, 1113, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Lyte, M. The effect of stress on microbial growth. Anim. Health Res. Rev. 2014, 15, 172–174. [Google Scholar] [CrossRef] [PubMed]
- West, J.W. Effects of heat-stress on production in dairy cattle. J. Dairy Sci. 2003, 86, 2131–2144. [Google Scholar] [CrossRef]
- O’Brien, M.D.; Rhoads, R.P.; Sanders, S.R.; Duff, G.C.; Baumgard, L.H. Metabolic adaptations to heat stress in growing cattle. Domest. Anim. Endocrinol. 2010, 38, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Monteiro, A.P.; Thompson, I.M.; Hayen, M.J.; Dahl, G.E. Effect of late-gestation maternal heat stress on growth and immune function of dairy calves. J. Dairy Sci. 2012, 95, 7128–7136. [Google Scholar] [CrossRef] [PubMed]
- Strong, R.A.; Silva, E.B.; Cheng, H.W.; Eicher, S.D. Acute brief heat stress in late gestation alters neonatal calf innate immune functions. J. Dairy Sci. 2015, 98, 7771–7783. [Google Scholar] [CrossRef] [PubMed]
- Crescio, M.I.; Forastiere, F.; Maurella, C.; Ingravalle, F.; Ru, G. Heat-related mortality in dairy cattle: A case crossover study. Pre-Vet. Med. 2010, 97, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J.A.; Burdick, N.C.; Chase, C.C., Jr.; Coleman, S.W.; Spiers, D.E. Influence of environmental temperature on the physiological, endocrine, and immune responses in livestock exposed to a provocative immune challenge. Domest. Anim. Endocrinol. 2012, 43, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Young, B.A. Ruminant cold stress-effect on production. J. Anim. Sci. 1983, 57, 1601–1607. [Google Scholar] [PubMed]
- Hoerlein, A.B.; Marsh, C.L. Studies on the epizootiology of shipping fever in calves. J. Am. Vet. Med. Assoc. 1957, 131, 123–127. [Google Scholar] [PubMed]
- Jensen, R.; Pierson, R.E.; Braddy, P.M.; Saari, D.A.; Lauerman, L.H.; England, J.J.; Keyvanfar, H.; Kauppinen, A.; Suuronen, T.; Ojala, J.; et al. Antagonistic crosstalk between NF-kappaB and SIRT1 in the regulation of inflammation and metabolic disorders. Cell Signal. 2013, 25, 1939–1948. [Google Scholar]
- Burdick, N.C.; Carroll, J.A.; Randel, R.D.; Willard, S.T.; Vann, R.C.; Chase, S.D.L., Jr.; Hulbert, L.E.; Welsh, T.H., Jr. Influence of temperament and transportation on physiological and endocrinological parameters in bulls. Livestock Sci. 2011, 139, 213–221. [Google Scholar] [CrossRef]
- Blecha, F.; Boyles, S.L.; Riley, J.G. Shipping suppresses lymphocyte blastogenic responses in Angus and Brahman X Angus feeder calves. Anim. Sci. 1984, 59, 576–583. [Google Scholar]
- Ishizaki, H.; Kariya, Y. Road Transportation Stress Promptly Increases Bovine Peripheral Blood Absolute NK Cell Counts and Cortisol Levels. J. Vet. Med. Sci. 2010, 72, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.J.; Santurtun, E. The welfare of livestock transported by ship. Vet. J. 2013, 196, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Earley, B.; Murray, M.; Prendiville, D.J.; Pintado, B.; Borque, C.; Canali, E. The effect of transport by road and sea on physiology, immunity and behaviour of beef cattle. Res. Vet. Sci. 2012, 92, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Griebel, P.; Schoonderwoerd, M.; Babiuk, L.A. Ontogeny of the immune response: Effect of protein-energy malnutrition in neonatal calves. Can. J. Vet. Res. 1987, 51, 429–435. [Google Scholar]
- Bourguet, C.; Deiss, V.; Boissy, A.; Andanson, S.; Terlouw, E.M. Effects of feed deprivation on behavioral reactivity and physiological status in Holstein cattle. J. Anim. Sci. 2011, 89, 3272–3285. [Google Scholar] [CrossRef] [PubMed]
- Galyean, M.L.; Lee, R.W.; Hubbert, M.E. Influence of fasting and transit on ruminal and blood metabolites in beef steers. J. Anim. Sci. 1981, 53, 7–18. [Google Scholar] [PubMed]
- Clarke, G.; Stilling, R.M.; Kennedy, P.J.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Minireview: Gut microbiota: The neglected endocrine organ. Mol. Cell. Endocrinol. 2014, 28, 1221–1238. [Google Scholar] [CrossRef] [PubMed]
- Faturi, C.B.; Tiba, P.A.; Kawakami, S.E.; Catallani, B.; Kerstens, M.; Suchecki, D. Disruptions of the mother-infant relationship and stress-related behaviours: Altered corticosterone secretion does not explain everything. Neurosci. Biobehav. Rev. 2010, 34, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Haley, D.B.; Bailey, D.W.; Stookey, J.M. The effects of weaning beef calves in two stages on their behavior and growth rate. J. Anim. Sci. 2005, 83, 2205–2214. [Google Scholar] [PubMed]
- Lefcourt, A.M.; Elsasser, T.H. Adrenal responses of Angus x Hereford cattle to the stress of weaning. J. Anim. Sci. 1995, 73, 2669–2676. [Google Scholar] [PubMed]
- O’Mahony, S.M.; Hyland, N.P.; Dinan, T.G.; Cryan, J.F. Maternal separation as a model of brain-gut axis dysfunction. Psychopharmacology (Berlin) 2011, 214, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J.A.; Arthington, J.D.; Chase, C.C., Jr. Early weaning alters the acute-phase reaction to an endotoxin challenge in beef calves. J. Anim. Sci. 2009, 87, 4167–4172. [Google Scholar] [CrossRef] [PubMed]
- Craig, J.V. Measuring social behavior: Social dominance. J. Anim. Sci. 1986, 62, 1120–1129. [Google Scholar] [PubMed]
- Ninomiya, S.; Sato, S. Assessment of the effect of presenting a companion’s face picture on social isolation stress using saliva sampling in cows. Anim. Sci. J. 2011, 82, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Munksgaard, L.; Simonsen, H.B. Behavioral and pituitary adrenal-axis responses of dairy cows to social isolation and deprivation of lying down. J. Anim. Sci. 1996, 74, 769–778. [Google Scholar] [PubMed]
- Van Reenen, C.G.; Mars, M.H.; Leushuis, I.E.; Rijsewijk, F.A.M.; van Oirschot, J.T.; Blokhuis, H.J. Social isolation may influence responsiveness to infection with bovine herpesvirus 1 in veal calves. Vet. Microbiol. 2000, 75, 135–143. [Google Scholar] [CrossRef]
- Stelwagen, K.; Hopster, H.; van der Werf, J.T.; Blokhuis, H.J. Short communication: Effects of isolation stress on mammary tight junctions in lactating dairy cows. J. Dairy Sci. 2000, 83, 48–51. [Google Scholar] [CrossRef]
- Von Keyserlingk, M.A.G.; Olenick, D.; Weary, D.M. Acute behavioral effects of regrouping dairy cows. J. Dairy Sci. 2008, 91, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- O’Driscou, K.; von Keyserlingk, M.A.G.; Weary, D.M. Effects of mixing on drinking and competitive behavior of dairy calves. J. Dairy Sci. 2006, 89, 229–233. [Google Scholar] [CrossRef]
- Philips, C.J.C.; Rind, M.I. The effects on production an behavior of mixing uniparous and multiparous cows. J. Dairy Sci. 2001, 84, 2424–2429. [Google Scholar] [CrossRef]
- Veissier, I.; Boissy, A.; dePassille, A.M.; Rushen, J.; van Reenen, C.G.; Roussel, S.; Andanson, S.; Pradel, P. Calves’ responses to repeated social regrouping and relocation. J. Anim. Sci. 2001, 79, 2580–2593. [Google Scholar] [PubMed]
- Andrade, O.; Orihuela, A.; Solano, J.; Galina, C.S. Some effects of repeated handling and the use of a mask on stress responses in Zebu cattle during restraint. Appl. Anim. Behav. Sci. 2001, 71, 175–181. [Google Scholar] [CrossRef]
- Herskin, M.S.; Munksgaard, L.; Andersen, J.B. Effects of social isolation and restraint on adrenocortical responses and hypoalgesia in loose-housed dairy cows. J. Anim. Sci. 2007, 85, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Szenci, O.; Karen, A.; Bajcsy, A.C.; Gaspardy, A.; de Sousa, N.M.; Beckers, J.F. Effect of restraint stress on plasma concentrations of cortisol, progesterone and pregnancy associated-glycoprotein-1 in pregnant heifers during late embryonic development. Theriogenology 2011, 76, 1380–1385. [Google Scholar] [CrossRef] [PubMed]
- Buynitsky, T.; Mostofsky, D.I. Restraint stress in biobehavioral research: Recent developments. Neurosci. Biobehav. Rev. 2009, 33, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.D.; Fulton, R.W.; Lehenbauer, T.W.; Step, D.L.; Confer, A.W. The epidemiology of bovine respiratory disease: What is the evidence for preventive measures? Can. Vet. J. 2010, 51, 1351–1359. [Google Scholar] [PubMed]
- Duff, G.C.; Galyean, M.L. Board-invited review: Recent advances in management of highly stressed, newly received feedlot cattle. J. Anim. Sci. 2007, 85, 823–840. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, L.M.; Raphael, W. Significance of metabolic stress, lipid mobilization, and inflammation on transition cow disorders. Vet. Clin. N. Am. Food Anim. Pract. 2013, 29, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Rostagno, M.H. Can stress in farm animals increase food safety risk? Foodborne Pathog. Dis. 2009, 6, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Bristow, D.J.; Holmes, D.S. Cortisol levels and anxiety-related behaviors in cattle. Physiol. Behav. 2007, 90, 626–628. [Google Scholar] [CrossRef] [PubMed]
- Curley, K.O., Jr.; Paschal, J.C.; Welsh, T.H., Jr.; Randel, R.D. Technical note: Exit velocity as a measure of cattle temperament is repeatable and associated with serum concentration of cortisol in Brahman bulls. J. Anim. Sci. 2006, 84, 3100–3103. [Google Scholar] [CrossRef] [PubMed]
- Grandin, T. Assessment of stress during handling and transport. J. Anim. Sci. 1997, 75, 249–257. [Google Scholar] [PubMed]
- Earley, B.; Drennan, M.; O’Riordan, E.G. The effect of road transport in comparison to a novel environment on the physiological, metabolic and behavioural responses of bulls. Res. Vet. Sci. 2013, 95, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Herskin, M.S.; Munksgaard, L.; Ladwig, J. Effects of acute stressors on nociception, adrenocortical responses and behavior of dairy cows. Physiol. Behav. 2004, 83, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Macbeth, B.J.; Cattet, M.R.L.; Stenhouse, G.B.; Gibeau, M.L.; Janz, D.M. Hair cortisol concentration as a noninvasive measure of long-term stress in free-ranging grizzly bears (Ursus arctos): Considerations with implications for other wildlife. Can. J. Zool. 2010, 88, 935–949. [Google Scholar] [CrossRef]
- Gonzalez-de-la-Vara, M.R.; Valdez, R.A.; Lemus-Ramirez, V.; Vazquez-Chagoyan, J.C.; Villa-Godoy, A.; Romano, M.C. Effects of adrenocorticotropic hormone challenge and age on hair cortisol concentrations in dairy cattle. Can. J. Vet. Res. 2011, 75, 216–221. [Google Scholar]
- Russell, E.; Koren, G.; Rieder, M.; van Uum, S. Hair cortisol as a biological marker of chronic stress: Current status, future directions and unanswered questions. Psychoneuroendocrinology 2012, 37, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Stalder, T.; Steudte, S.; Miller, R.; Skoluda, N.; Dettenborn, L.; Kirschbaum, C. Intraindividual stability of hair cortisol concentrations. Psychoneuroendocrinology 2012, 37, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Lynch, E.M.; Earley, B.; McGee, M.; Doyle, S. Effect of abrupt weaning at housing on leukocyte distribution, functional activity of neutrophils, and acute phase protein response of beef calves. BMC Vet. Res. 2010, 6, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Loughlin, A.; McGee, M.; Waters, S.M.; Doyle, S.; Earley, B. Examination of the bovine leukocyte environment using immunogenetic biomarkers to assess immunocompetence following exposure to weaning stress. BMC Vet. Res. 2011, 7, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Clercq, N.; Julie, V.B.; Croubels, S.; Delahaut, P.; Vanhaecke, L. A validated analytical method to study the long-term stability of natural and synthetic glucocorticoids in livestock urine using ultra-high performance liquid chromatography coupled to Orbitrap-high resolution mass spectrometry. J. Chromatogr. A 2013, 1301, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Loberg, J.M.; Hernandez, C.E.; Thierfelder, T.; Jensen, M.B.; Berg, C.; Lidfors, L. Weaning and separation in two steps-A way to decrease stress in dairy calves suckled by foster cows. Appl. Anim. Behav. Sci. 2008, 111, 222–234. [Google Scholar] [CrossRef]
- Fukasawa, M.; Tsukada, H. Relationship between milk cortisol concentration and the behavioral characteristics of postpartum cows introduced to a new group. Anim. Sci. J. 2010, 81, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Mostl, E.; Maggs, J.L.; Schrotter, G.; Besenfelder, V.; Palme, R. Measurement of cortisol metabolites in Faeces of Ruminants. Vet. Res. Commun. 2002, 26, 127–139. [Google Scholar] [CrossRef] [PubMed]
- McCorkell, R.; Wynne-Edwards, K.; Galbraith, J.; Schaefer, A.; Caulkett, N.; Boysen, S.; Pajor, E.; the class of UCVM class of 2012. Transport versus on-farm slaughter of bison: Physiological stress, animal welfare, and avoidable trim losses. Can. Vet. J. 2013, 54, 769–774. [Google Scholar] [PubMed]
- Hulbert, L.E.; Carroll, J.A.; Balloum, A.; Burdick, N.C.; Dailey, J.W.; Caldwell, L.C.; Loyd, A.N.; Vann, R.C.; Welsh, T.H.; Randel, R.D.; et al. Sexually dimorphic stress and pro-inflammatory cytokine responses to an intravenous corticotrophin-releasing hormone challenge of Brahman cattle following transportation. Innate Immun. 2013, 19, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Van Westerloo, D.J.; Choi, G.; Lowenberg, E.C.; Truijen, J.; de Vos, A.F.; Endert, E.; Meijers, J.C.; Zhou, L.; Pereira, M.P.; Queiroz, K.C.; et al. Acute stress elicited by bungee jumping suppresses human innate immunity. Mol. Med. 2011, 17, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Arsenault, R.J.; Li, Y.; Bell, K.; Doig, K.; Potter, A.; Griebel, P.J.; Kusalik, A.; Napper, S. Mycobacterium avium subsp. paratuberculosis inhibits gamma interferon-induced signaling in bovine monocytes: Insights into the cellular mechanisms of Johne’s disease. Infect. Immun. 2012, 80, 3039–3048. [Google Scholar] [CrossRef] [PubMed]
- Arsenault, R.J.; Li, Y.; Maattanen, P.; Scruten, E.; Doig, K.; Potter, A.; Griebel, P.; Kusalik, A.; Napper, S. Altered Toll-like receptor 9 signaling in Mycobacterium avium subsp. paratuberculosis-infected bovine monocytes reveals potential therapeutic targets. Infect Immun. 2013, 81, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Maattanen, P.; Trost, B.; Scruten, E.; Potter, A.; Kusalik, A.; Griebel, P.; Napper, S. Divergent immune responses to mycobacterium avium subsp paratuberculosis infection correlate with kinome responses at the site of intestinal infection. Infect. Immun. 2013, 81, 2861–2872. [Google Scholar] [CrossRef] [PubMed]
- Trost, B.; Kindrachuk, J.; Scruten, E.; Griebel, P.; Kusalik, A.; Napper, S. Kinotypes: Stable species- and individual-specific profiles of cellular kinase activity. BMC Genom. 2013, 14, 854. [Google Scholar] [CrossRef] [PubMed]
- Napper, S.; Dadgar, S.; Arsenault, R.J.; Trost, B.; Scruten, E.; Kusalik, A.; Shand, P. Induction of tissue- and stressor-specific kinomic responses in chickens exposed to hot and cold stresses. Poult. Sci. 2015, 94, 1333–1345. [Google Scholar] [CrossRef] [PubMed]
- Arsenault, R.J.; Napper, S.; Kogut, M.H. Salmonella enterica Typhimurium infection causes metabolic changes in chicken muscle involving AMPK, fatty acid and insulin/mTOR signaling. Vet. Res. 2013, 44, 35. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.J.; Trost, B.; Scruten, E.; Robertson, T.; Mostajeran, M.; Connor, W.; Kusalik, A.; Griebel, P.; Napper, S. Identification of developmentally-specific kinotypes and mechanisms of Varroa mite resistance through whole-organism, kinome analysis of honeybee. Front. Genet. 2014, 5, 139. [Google Scholar] [CrossRef] [PubMed]
- Lyles, J.L.; Calvo-Lorenzo, M.S.; Bill, E. Kunkle Interdisciplinary Beef Symposium: Practical developments in managing animal welfare in beef cattle: What does the future hold? J. Anim. Sci. 2014, 92, 5334–5344. [Google Scholar] [CrossRef] [PubMed]
- Marti, S.; Devant, M.; Amatayakul-Chantler, S.; Jackson, J.A.; Lopez, E.; Janzen, E.D.; Schwartzkopf-Genswein, K.S. Effect of anti-gonadotropin-releasing factor vaccine and band castration on indicators of welfare in beef cattle. J. Anim. Sci. 2015, 93, 1581–1591. [Google Scholar] [CrossRef] [PubMed]
- DuPont, R.L.; Rice, D.P.; Miller, L.S.; Shiraki, S.S.; Rowland, C.R.; Harwood, H.J. Economic costs of anxiety disorders. Anxiety 1996, 2, 167–172. [Google Scholar] [CrossRef]
- Lepine, J.P. The epidemiology of anxiety disorders: Prevalence and societal costs. J. Clin. Psychiatry 2002, 63, 4–8. [Google Scholar] [PubMed]
- Greenberg, P.E.; Sisitsky, T.; Kessler, R.C.; Finkelstein, S.N.; Berndt, E.R.; Davidson, J.R.; Ballenger, J.C.; Fyer, A.J. The economic burden of anxiety disorders in the 1990s. J. Clin. Psychiatry 1999, 60, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Schniering, C.A.; Hudson, J.L.; Rapee, R.M. Issues in the diagnosis and assessment of anxiety disorders in children and adolescents. Clin. Psychol. Rev. 2000, 20, 453–478. [Google Scholar] [CrossRef]
- Hardeveld, F.; Spijker, J.; Vreeburg, S.A.; Graaf, R.D.; Hendriks, S.M.; Licht, C.M.; Nolen, W.A.; Penninx, B.W.; Beekman, A.T. Increased cortisol awakening response was associated with time to recurrence of major depressive disorder. Psychoneuroendocrinology 2014, 50, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Seok, J.; Warren, H.S.; Cuenca, A.G.; Mindrinos, M.N.; Baker, H.V.; Xu, W.; Richards, D.R.; McDonald-Smith, G.P.; Gao, H.; Hennessy, L.; et al. Inflammation and Host Response to Injury Large Scale Collaborative Research Program. Genomic responses in mouse models poorly mimic human inflammatory diseases. PNAS 2013, 110, 3507–3512. [Google Scholar] [CrossRef] [PubMed]
- Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The pig: A model for human infectious diseases. Trends Microbiol. 2012, 20, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Basel, M.T.; Balivada, S.; Beck, A.P.; Kerrigan, M.A.; Pyle, M.M.; Dekkers, J.C.; Wyatt, C.R.; Rowland, R.R.; Anderson, D.E.; Bossmann, S.H.; et al. Human xenografts are not rejected in a naturally occurring immunodeficient porcine line: A human tumor model in pigs. Biores Open Access. 2012, 1, 63–68. [Google Scholar] [CrossRef] [PubMed]
- White, F.C.; Roth, D.M.; Bloor, C.M. The pig as a model for myocardial ischemia and exercise. J. Am. Assoc. Lab. Anim. Sci. 1986, 36, 351–356. [Google Scholar]
- Miller, T.L.; Touch, S.M.; Shaffer, T.H. Matrix metalloproteinase and tissue inhibitor of matrix metalloproteinase expression profiles in tracheal aspirates do not adequately reflect tracheal or lung tissue profiles in neonatal respiratory distress: Observations from an animal model. Pediatr. Crit. Care Med. 2006, 7, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Lalwani, A.K.; Walsh, B.J.; Reilly, P.G.; Muzyczka, N.; Mhatre, A.N. Development of in vivo gene therapy for hearing disorders: Introduction of adeno-associated virus into the cochlea of the guinea pig. Gene Ther. 1996, 3, 588–592. [Google Scholar] [PubMed]
- Bendele, A.; McComb, J.; Gould, T.; McAbee, T.; Sennello, G.; Chlipala, E.; Guy, M. Animal models of arthritis: Relevance to human disease. Toxicol. Pathol. 1999, 27, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, T.P.; Eaglstein, W.H.; Davis, S.C.; Mertz, P. The pig as a model for human wound healing. Wound Repair Regen. 2001, 9, 66–76. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Arsenault, R.; Napper, S.; Griebel, P. Models and Methods to Investigate Acute Stress Responses in Cattle. Animals 2015, 5, 1268-1295. https://doi.org/10.3390/ani5040411
Chen Y, Arsenault R, Napper S, Griebel P. Models and Methods to Investigate Acute Stress Responses in Cattle. Animals. 2015; 5(4):1268-1295. https://doi.org/10.3390/ani5040411
Chicago/Turabian StyleChen, Yi, Ryan Arsenault, Scott Napper, and Philip Griebel. 2015. "Models and Methods to Investigate Acute Stress Responses in Cattle" Animals 5, no. 4: 1268-1295. https://doi.org/10.3390/ani5040411
APA StyleChen, Y., Arsenault, R., Napper, S., & Griebel, P. (2015). Models and Methods to Investigate Acute Stress Responses in Cattle. Animals, 5(4), 1268-1295. https://doi.org/10.3390/ani5040411