Cognitive Bias in Zoo Animals: An Optimistic Outlook for Welfare Assessment

Simple Summary
Abstract
1. Introduction
2. Cognitive Bias and Animal Welfare
3. Measuring Cognitive Bias
4. Cognitive Bias Testing on Exotic Species Housed in Zoos and Other Settings
4.1. Cognitive Bias Tests in Zoos
4.2. Cognitive Bias Tests on Zoo Species Kept in Other Settings
5. Why Should Zoos Conduct Cognitive Bias Research?
5.1. Cognitive Bias Tests Can Inform Us About the Animals’ Welfare
5.2. Cognitive Bias Tasks Themselves Can be Enriching for Animals
5.3. Findings Can Improve Management of Animals
5.4. Cognitive Bias Research in Zoos Has Intrinsic Value
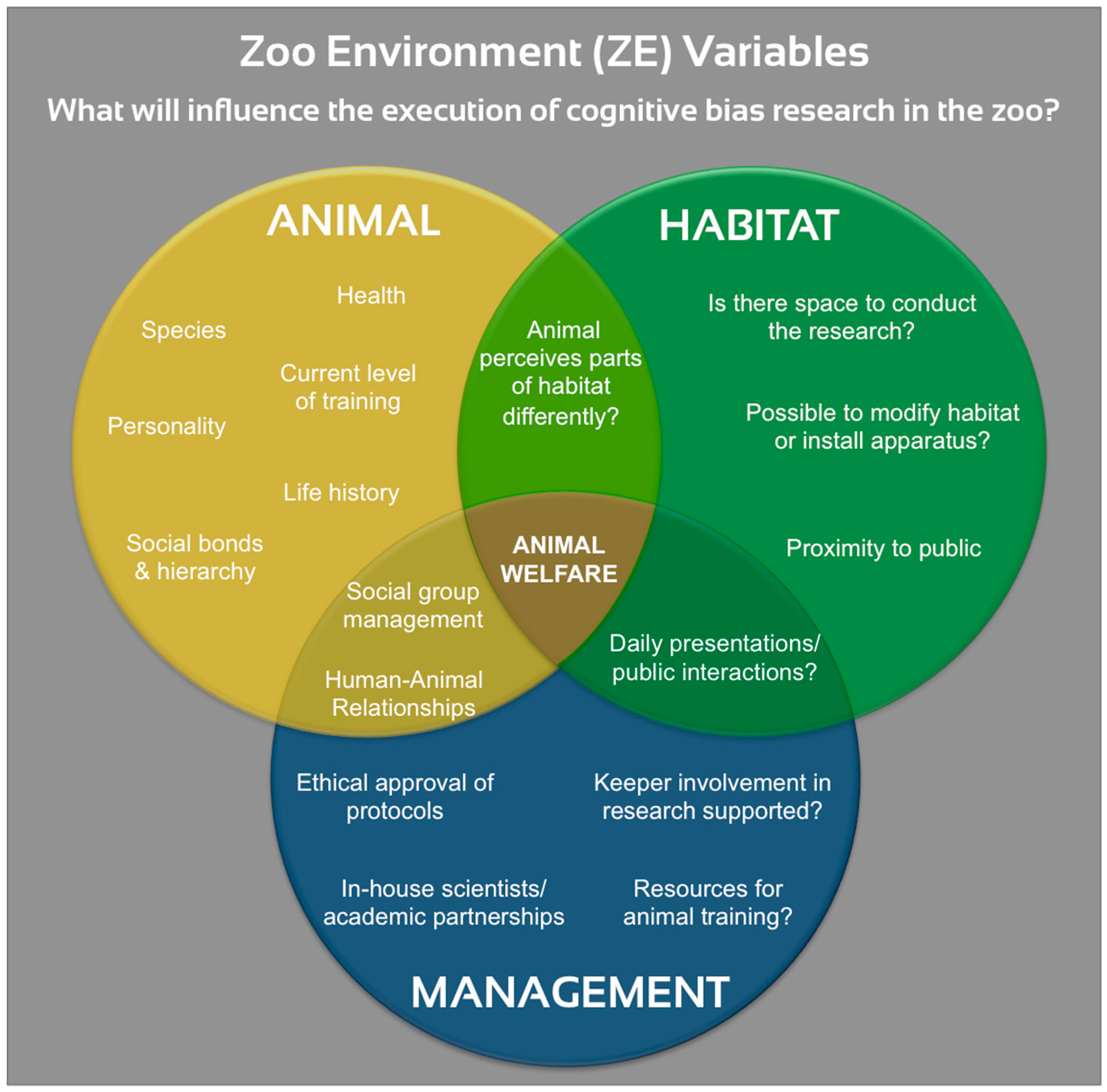
6. Recommendations for Cognitive Bias Tests in Zoo Settings
6.1. Which Animals, and What Type of Test?
6.2. Study Goals: Feasible for Animals, Keepers and Scientists
6.2.1. Ask a Research Question Relevant to Subject Animals
6.2.2. Thoroughly Involve Animal Caretakers at All Stages
6.2.3. Collaborate with Academic Institutions
6.3. Designing the Task to Measure Cognitive Bias
6.3.1. Appropriate Cue Choice
6.3.2. Test Procedure
6.3.3. Social Isolation: Not Always Necessary
6.3.4. Interpretations and Limitations of Cognitive Bias Data
7. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Mendl, M.; Burman, O.; Parker, R.M.A.; Paul, E.S. Cognitive bias as an indicator of animal emotion and welfare: Emerging evidence and underlying mechanisms. Appl. Anim. Behav. Sci. 2009, 118, 161–181. [Google Scholar] [CrossRef]
- Eysenck, M.W.; Mogg, K.; May, J.; Richards, A.; Mathews, A. Bias in interpretation of ambiguous sentences related to threat in anxiety. J. Abnorm. Psychol. 1991, 100, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Wright, W.F.; Bower, G.H. Mood effects on subjective probability assessment. Organ. Behav. Hum. Decis. Process. 1992, 52, 276–291. [Google Scholar] [CrossRef]
- Harding, E.J.; Paul, E.S.; Mendl, M. Cognitive bias and affective state. Nature 2004, 427, 312. [Google Scholar] [CrossRef] [PubMed]
- Dawkins, M.S. Animal welfare and the paradox of animal consciousness. Adv. Study Behav. 2015, 47, 5–38. [Google Scholar] [CrossRef]
- Mason, G.; Veasey, J.S. How should the psychological well-being of zoo elephants be objectively investigated? Zoo Biol. 2010, 29, 237–255. [Google Scholar] [CrossRef] [PubMed]
- Spruijt, B.M.; van den Bos, R.; Pijlman, F.T.A. A concept of welfare based on reward evaluating mechanisms in the brain: Anticipatory behaviour as an indicator for the state of reward systems. Appl. Anim. Behav. Sci. 2001, 72, 145–171. [Google Scholar] [CrossRef]
- Mendl, M.; Burman, O.H.P.; Paul, E.S. An integrative and functional framework for the study of animal emotion and mood. Proc. Biol. Sci. 2010, 277, 2895–2904. [Google Scholar] [CrossRef] [PubMed]
- Webster, J. Animal Welfare: Limping towards Eden; Blackwell Publishing Ltd.: Oxford, UK, 2005; ISBN 1-4051-1877-6. [Google Scholar]
- Boissy, A.; Manteuffel, G.; Jensen, M.B.; Moe, R.O.; Spruijt, B.; Keeling, L.J.; Winckler, C.; Forkman, B.; Dimitrov, I.; Langbein, J.; et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 2007, 92, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Titulaer, M.; Blackwell, E.J.; Mendl, M.; Casey, R.A. Cross sectional study comparing behavioural, cognitive and physiological indicators of welfare between short and long term kennelled domestic dogs. Appl. Anim. Behav. Sci. 2013, 147, 149–158. [Google Scholar] [CrossRef]
- Baciadonna, L.; McElligott, A.G. The use of judgement bias to assess welfare in farm livestock. Anim. Welf. 2015, 24, 81–91. [Google Scholar] [CrossRef]
- Roelofs, S.; Boleij, H.; Nordquist, R.; van der Staay, F.J. Making decisions under ambiguity: Judgment bias tasks for assessing emotional state in animals. Front. Behav. Neurosci. 2016, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Clegg, I.L.K.; Rödel, H.G.; Delfour, F. Bottlenose dolphins engaging in more social affiliative behaviour judge ambiguous cues more optimistically. Behav. Brain Res. 2017, 322, 115–122. [Google Scholar] [CrossRef] [PubMed]
- McGuire, M.C.; Vonk, J.; Fuller, G.; Allard, S. Using an ambiguous cue paradigm to assess cognitive bias in Gorillas (Gorilla gorilla gorilla) during a forage manipulation. Anim. Behav. Cogn. 2017, 4, 91–104. [Google Scholar] [CrossRef]
- McGuire, M.C.; Vonk, J.; Johnson-Ulrich, Z. Ambiguous results when using the ambiguous-cue paradigm to assess learning and cognitive bias in Gorillas and a Black Bear. Behav. Sci. 2017, 7, 51. [Google Scholar] [CrossRef] [PubMed]
- Whitham, J.C.; Wielebnowski, N. New directions for zoo animal welfare science. Appl. Anim. Behav. Sci. 2013, 147, 247–260. [Google Scholar] [CrossRef]
- Clegg, I.L.K.; Borger-Turner, J.L.; Eskelinen, H.C. C-Well: The development of a welfare assessment index for captive bottlenose dolphins (Tursiops truncatus). Anim. Welf. 2015, 24, 267–282. [Google Scholar] [CrossRef]
- Maple, T.L. Toward a science of welfare for animals in the zoo. J. Appl. Anim. Welf. Sci. 2007, 10, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Maple, T.; Perdue, B.M. Zoo Animal Welfare; Springer: Berlin, Germany, 2013; Volume 14, ISBN 978-3-642-35954-5. [Google Scholar]
- Bethell, E.J. A “How-To” guide for designing judgment bias studies to assess captive animal welfare. J. Appl. Anim. Welf. Sci. 2015, 18, S18–S42. [Google Scholar] [CrossRef] [PubMed]
- Gygax, L. The A to Z of statistics for testing cognitive judgement bias. Anim. Behav. 2014, 95, 59–69. [Google Scholar] [CrossRef]
- Paul, E.S.; Harding, E.J.; Mendl, M. Measuring emotional processes in animals: The utility of a cognitive approach. Neurosci. Biobehav. Rev. 2005, 29, 469–491. [Google Scholar] [CrossRef] [PubMed]
- Brajon, S.; Laforest, J.P.; Schmitt, O.; Devillers, N. The way humans behave modulates the emotional state of piglets. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Zidar, J.; Campderrich, I.; Ansson, E.; Wichman, A.; Winberg, S.; Keeling, L.; Løvlie, H. Environmental complexity buffers against stress-induced negative judgement bias in female chickens. Sci. Rep. 2018, 8, 5404. [Google Scholar] [CrossRef] [PubMed]
- Désiré, L.; Boissy, A.; Veissier, I. Emotions in farm animals: A new approach to animal welfare in applied ethology. Behav. Process. 2002, 60, 165–180. [Google Scholar] [CrossRef]
- Barnett, J.L.; Hemsworth, P.H. The validity of physiological and behavioural measures of animal welfare. Appl. Anim. Behav. Sci. 1990, 25, 177–187. [Google Scholar] [CrossRef]
- Reefmann, N.; Bütikofer Kaszàs, F.; Wechsler, B.; Gygax, L. Physiological expression of emotional reactions in sheep. Physiol. Behav. 2009, 98, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Hemsworth, P.H.; Mellor, D.J.; Cronin, G.M.; Tilbrook, A.J. Scientific assessment of animal welfare. N. Z. Vet. J. 2015, 63, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Yeates, J.W.; Main, D.C.J. Assessment of positive welfare: A review. Vet. J. 2008, 175, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Menargues Marcilla, A.; Urios, V.; Limiñana, R. Seasonal rhythms of salivary cortisol secretion in captive Asian elephants (Elephas maximus). Gen. Comp. Endocrinol. 2012, 176, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Montanha, J.C.; Silva, S.L.; Boere, V. Comparison of salivary cortisol concentrations in Jaguars kept in captivity with differences in exposure to the public. Ciência Rural 2009, 39, 1745–1751. [Google Scholar] [CrossRef]
- Clegg, I.L.K.; Rödel, H.G.; Cellier, M.; Vink, D.; Michaud, I.; Mercera, B.; Böye, M.; Hausberger, M.; Lemasson, A.; Delfour, F. Schedule of human-controlled periods structures bottlenose dolphin (Tursiops truncatus) behavior in their free-time. J. Comp. Psychol. 2017, 131, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Horback, K.M.; Miller, L.J.; Andrews, J.R.M.; Kuczaj, S.A. Diurnal and nocturnal activity budgets of zoo elephants in an outdoor facility. Zoo Biol. 2014, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Salmeto, A.L.; Hymel, K.A.; Carpenter, E.C.; Brilot, B.O.; Bateson, M.; Sufka, K.J. Cognitive bias in the chick anxiety-depression model. Brain Res. 2011, 1373, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Bateson, M.; Nettle, D. Development of a cognitive bias methodology for measuring low mood in chimpanzees. PeerJ 2015, 3, e998. [Google Scholar] [CrossRef] [PubMed]
- Burman, O.; McGowan, R.; Mendl, M.; Norling, Y.; Paul, E.; Rehn, T.; Keeling, L. Using judgement bias to measure positive affective state in dogs. Appl. Anim. Behav. Sci. 2011, 132, 160–168. [Google Scholar] [CrossRef]
- Wichman, A.; Keeling, L.J.; Forkman, B. Cognitive bias and anticipatory behaviour of laying hens housed in basic and enriched pens. Appl. Anim. Behav. Sci. 2012, 140, 62–69. [Google Scholar] [CrossRef]
- Asher, L.; Friel, M.; Griffin, K.; Collins, L.M. Mood and personality interact to determine cognitive biases in pigs. Biol. Lett. 2016, 12. [Google Scholar] [CrossRef] [PubMed]
- Cussen, V.A.; Mench, J.A. Personality predicts cognitive bias in captive psittacines, Amazona amazonica. Anim. Behav. 2014, 89, 123–130. [Google Scholar] [CrossRef]
- Lecorps, B.; Weary, D.M.; Von Keyserlingk, M.A.G. Pessimism and fearfulness in dairy calves. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Clegg, I.L.K.; Delfour, F. Cognitive judgement bias is associated with frequency of anticipatory behavior in bottlenose dolphins. Zoo Biol. 2018, 37, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Mendl, M.; Brooks, J.; Basse, C.; Burman, O.; Paul, E.; Blackwell, E.; Casey, R. Dogs showing separation-related behaviour exhibit a “pessimistic” cognitive bias. Curr. Biol. 2010, 20, R839–R840. [Google Scholar] [CrossRef] [PubMed]
- Raoult, C.M.C.; Moser, J.; Gygax, L. Mood as cumulative expectation mismatch: A test of theory based on data from non-verbal cognitive bias tests. Front. Psychol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Bethell, E.J.; Holmes, A.; MacLarnon, A.; Semple, S. Evidence that emotion mediates social attention in rhesus macaques. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Rodd, Z.A.; Rosellini, R.A.; Stock, H.S.; Gallup, G.G., Jr. Learned helplessness in chickens (Gallus gallus): Evidence for attentional bias. Learn. Motiv. 1997, 28, 43–55. [Google Scholar] [CrossRef]
- Paré, W.P. Enhanced retrieval of unpleasant memories influenced by shock controllability, shock sequence, and rat strain. Biol. Psychiatry 1996, 39, 808–813. [Google Scholar] [CrossRef]
- Kumar, K.B.; Karanth, K.S. Enhanced retrieval of unpleasant memory in helpless rats. Biol. Psychiatry 1991, 30, 493–501. [Google Scholar] [CrossRef]
- Monk, J.E.; Doyle, R.E.; Colditz, I.G.; Belson, S.; Cronin, G.M.; Lee, C. Towards a more practical attention bias test to assess affective state in sheep. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Ash, H.; Buchanan-Smith, H.M. The long-term impact of infant rearing background on the affective state of adult common marmosets (Callithrix jacchus). Appl. Anim. Behav. Sci. 2016, 174, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Bethell, E.J.; Holmes, A.; MacLarnon, A.; Semple, S. Cognitive bias in a non-human primate: Husbandry procedures influence cognitive indicators of psychological well-being in captive rhesus macaques. Anim. Welf. 2012, 21, 185–195. [Google Scholar] [CrossRef]
- Bethell, E.; Holmes, A.; MacLarnon, A.; Semple, S. Emotion evaluation and response slowing in a non-human primate: New directions for cognitive bias measures of animal emotion? Behav. Sci. (Basel) 2016, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Cronin, K.A.; Bethell, E.J.; Jacobson, S.L.; Egelkamp, C.; Hopper, L.M.; Ross, S.R. Evaluating mood changes in response to anthropogenic noise with a response-slowing task in three species of zoo-housed primates. Anim. Behav. Cogn. 2018, 5, 209–221. [Google Scholar] [CrossRef]
- Gordon, D.J.; Rogers, L.J. Cognitive bias, hand preference and welfare of common marmosets. Behav. Brain Res. 2015, 287, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Keen, H.A.; Nelson, O.L.; Robbins, C.T.; Evans, M.; Shepherdson, D.J.; Newberry, R.C. Validation of a novel cognitive bias task based on difference in quantity of reinforcement for assessing environmental enrichment. Anim. Cogn. 2014, 17, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Lalot, M.; Ung, D.; Péron, F.; D’Ettorre, P.; Bovet, D. You know what? I’m happy. Cognitive bias is not related to personality but is induced by pair-housing in canaries (Serinus canaria). Behav. Process. 2017, 134, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Marzouki, Y.; Gullstrand, J.; Goujon, A.; Fagot, J. Baboons’ response speed is biased by their moods. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, S.S.D.C.; Fernandes, I.K.; Costa, T.S.O.; Nogueira-Filho, S.L.G.; Mendl, M. Does trapping influence decision-making under ambiguity in white-lipped peccary (Tayassu pecari)? PLoS ONE 2015, 10, e0127868. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.R.M.; Nogueira-Filho, S.L.G.; Sousa, M.B.C.; Dias, C.T.S.; Mendl, M.; Nogueira, S.S.C. Measurement of cognitive bias and cortisol levels to evaluate the effects of space restriction on captive collared peccary (Mammalia, Tayassuidae). Appl. Anim. Behav. Sci. 2016, 181, 76–82. [Google Scholar] [CrossRef]
- Pomerantz, O.; Terkel, J.; Suomi, S.J.; Paukner, A. Stereotypic head twirls, but not pacing, are related to a ’pessimistic’-like judgment bias among captive tufted capuchins (Cebus apella). Anim. Cogn. 2012, 15, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Schino, G.; Massimei, R.; Pinzaglia, M.; Addessi, E. Grooming, social rank and ‘optimism’ in tufted capuchin monkeys: A study of judgement bias. Anim. Behav. 2016, 119, 11–16. [Google Scholar] [CrossRef]
- Zeder, M.A. Core questions in domestication research. Proc. Natl. Acad. Sci. USA 2015, 112, 3191–3198. [Google Scholar] [CrossRef] [PubMed]
- Matheson, S.M.; Asher, L.; Bateson, M. Larger, enriched cages are associated with “optimistic” response biases in captive European starlings (Sturnus vulgaris). Appl. Anim. Behav. Sci. 2008, 109, 374–383. [Google Scholar] [CrossRef]
- Bateson, M.; Matheson, S.M. Performance on a categorisation task suggests that removal of environmental enrichment induces “pessimism” in captive European starlings (Sturnus vulgaris). Anim. Welf. 2007, 16, 33–36. [Google Scholar]
- Brilot, B.O.; Asher, L.; Bateson, M. Stereotyping starlings are more “pessimistic”. Anim. Cogn. 2010, 13, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Brilot, B.O.; Normandale, C.L.; Parkin, A.; Bateson, M. Can we use starlings’ aversion to eyespots as the basis for a novel “cognitive bias” task? Appl. Anim. Behav. Sci. 2009, 118, 182–190. [Google Scholar] [CrossRef]
- Mason, G.; Mendl, M. Why is there no simple way of measuring animal welfare? Anim. Welf. 2007, 2, 301–319. [Google Scholar]
- Botreau, R.; Bonde, M.; Butterworth, A.; Perny, P.; Bracke, M.B.M.; Capdeville, J.; Veissier, I. Aggregation of measures to produce an overall assessment of animal welfare. Part 1: A review of existing methods. Animal 2007, 1, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Rose, P. Ensuring a Good Quality of Life in the Zoo: Underpinning Welfare-positive animal management with ecological evidence. In Zoo Animals: Husbandry, Welfare and Public Interactions; Berger, M., Corbett, S., Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2018; pp. 141–198. [Google Scholar]
- Berger, M.; Corbett, S. Zoo Animals: Husbandry, Welfare and Public Interactions; Nova Science Publishers Inc.: New York, NY, USA, 2018; ISBN 978-1-53613-536-7. [Google Scholar]
- Blackett, T.A.; McKenna, C.; Kavanagh, L.; Morgan, D.R. The welfare of wild animals in zoological institutions: Are we meeting our duty of care? Int. Zoo Yearb. 2017, 51, 187–202. [Google Scholar] [CrossRef]
- Marchant-Forde, J.N. The science of animal behavior and welfare: Challenges, opportunities, and global perspective. Front. Vet. Sci. 2015, 2, 16. [Google Scholar] [CrossRef] [PubMed]
- Clegg, I.L.K. Developing Welfare Parameters for Bottlenose Dolphins (Tursiops truncatus) under Human Care. Ph.D. Thesis, Université Paris, Paris, France, 2017. [Google Scholar]
- Von Fersen, L.; Encke, D.; Huttner, T.; Baumgartner, K. Establishment and implementation of an animal welfare decision tree to evaluate the welfare of zoo animals. Aquat. Mamm. 2018, 44, 211–220. [Google Scholar] [CrossRef]
- Clayton, S.; Fraser, J.; Saunders, C.D. Zoo experiences: Conversations, connections, and concern for animals. Zoo Biol. 2009, 28, 377–397. [Google Scholar] [CrossRef] [PubMed]
- Reade, L.S.; Waran, N.K. The modern zoo: How do people perceive zoo animals? Appl. Anim. Behav. Sci. 1996, 47, 109–118. [Google Scholar] [CrossRef]
- Price, A.; Boeving, E.R.; Shender, M.A.; Ross, S.R. Understanding the effectiveness of demonstration programs. J. Mus. Educ. 2015, 40, 46–54. [Google Scholar] [CrossRef]
- Alba, A.C.; Leighty, K.A.; Pittman Courte, V.L.; Grand, A.P.; Bettinger, T.L. A turtle cognition research demonstration enhances visitor engagement and keeper-animal relationships. Zoo Biol. 2017, 36, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, S.T.; Maxwell, A.; Hansen, K.A. Research as an enrichment tool to improve welfare in captive animals. Anim. Husb. Dairy Vet. Sci. 2017, 2, 1–6. [Google Scholar] [CrossRef]
- Riley, L.M. Conserving behaviour with cognitive enrichment: A new frontier for zoo animal management? In Zoo Animals: Husbandry, Welfare and Public Interactions; Berger, M., Corbett, S., Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2018; pp. 142–199. [Google Scholar]
- Hoy, J.M.; Murray, P.J.; Tribe, A. Thirty years later: Enrichment practices for captive mammals. Zoo Biol. 2010, 29, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Newberry, R.C. Environmental enrichment: Increasing the biological relevance of captive environments. Appl. Anim. Behav. Sci. 1995, 44, 229–243. [Google Scholar] [CrossRef]
- Laule, G. The use of behavioral management techniques to reduce or eliminate abnormal behavior. Anim. Welf. Inf. Cent. Newsl. 1993, 4, 8–11. [Google Scholar]
- Colahan, H.; Breder, C. Primate training at Disney’s Animal Kingdom. J. Appl. Anim. Welf. Sci. 2003, 6, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Melfi, V. Is training zoo animals enriching? Appl. Anim. Behav. Sci. 2013, 147, 299–305. [Google Scholar] [CrossRef]
- Brando, S.I.C.A. Animal Learning and Training. Implications for animal welfare. Vet. Clin. N. Am. Exot. Anim. Pract. 2012, 15, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Laule, G.E.; Bloomsmith, M.A.; Schapiro, S.J. The use of positive reinforcement training techniques to enhance the care, management, and welfare of primates in the laboratory. J. Appl. Anim. Welf. Sci. 2003, 6, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Bloomsmith, M.A.; Jones, M.L.; Snyder, R.J.; Singer, R.A.; Gardner, W.A.; Liu, S.C.; Maple, T.L. Positive reinforcement training to elicit voluntary movement of two giant pandas throughout their enclosure. Zoo Biol. 2003, 22, 323–334. [Google Scholar] [CrossRef]
- Pomerantz, O.; Terkel, J. Effects of positive reinforcement training techniques on the psychological welfare of zoo-housed chimpanzees (Pan troglodytes). Am. J. Primatol. 2009, 71, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Clark, F. Cognitive enrichment and welfare: Current approaches and future directions. Anim. Behav. Cogn. 2017, 4, 52–71. [Google Scholar] [CrossRef]
- Svendsen, P.M. The effect of handling and training on measures of the affective state of farmed mink (Neovison vison). In Proceedings of the Xth International Scientific Congress in fur Animal Production; Larsen, P.F., Møller, S.H., Clausen, T., Hammer, A.S., Lássen, T.M., Nielsen, V.H., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2012; pp. 360–367. [Google Scholar]
- Clegg, I.L.K.; Rödel, H.G.; Boivin, X.; Delfour, F. Looking forward to interacting with their caretakers: Dolphins’ anticipatory behaviour indicates motivation to participate in specific events. Appl. Anim. Behav. Sci. 2018, 22, 85–93. [Google Scholar] [CrossRef]
- Hosey, G.; Melfi, V. Human-animal interactions, relationships and bonds: A review and analysis of the literature. Int. J. Comp. Psychol. 2014, 27, 117–142. [Google Scholar] [CrossRef]
- Hintze, S.; Melotti, L.; Colosio, S.; Bailoo, J.D.; Boada-Saña, M.; Würbel, H.; Murphy, E. A cross-species judgement bias task: Integrating active trial initiation into a spatial Go/No-go task. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.H.; Munafò, M.R.; Robinson, E.S.J. Investigating the psychopharmacology of cognitive affective bias in rats using an affective tone discrimination task. Psychopharmacology (Berlin) 2013, 226, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Melfi, V.A. There are big gaps in our knowledge, and thus approach, to zoo animal welfare: A case for evidence-based zoo animal management. Zoo Biol. 2009, 28, 574–588. [Google Scholar] [CrossRef] [PubMed]
- Lemos de Figueiredo, R. Remembering the forgotten: Highlighting the need for more research and education on understudied zoo-housed taxa. In Zoo Animals: Husbandry, Welfare and Public Interactions; Berger, M., Corbett, S., Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2018; pp. 293–298. [Google Scholar]
- Williams, E.; Bremner-Harrison, S.; Ward, S.J. Can we meet the needs of social species in zoos? An overview of the impact of group housing on welfare in socially housed zoo mammals. In Zoo Animals: Husbandry, Welfare and Public Interactions; Nova Science Publishers: Hauppauge, NY, USA, 2018; pp. 1–88. [Google Scholar]
- Montaudouin, S.; Le Pape, G. Comparison between 28 zoological parks: Stereotypic and social behaviours of captive brown bears (Ursus arctos). Appl. Anim. Behav. Sci. 2005, 92, 129–141. [Google Scholar] [CrossRef]
- Waples, K.A.; Gales, N.J. Evaluating and minimising social stress in the care of captive bottlenose dolphins (Tursiops aduncus). Zoo Biol. 2002, 21, 5–26. [Google Scholar] [CrossRef]
- Barker, T.H.; George, R.P.; Howarth, G.S.; Whittaker, A.L. Assessment of housing density, space allocation and social hierarchy of laboratory rats on behavioural measures of welfare. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Behringer, V.; Stevens, J.M.G.; Deschner, T.; Hohmann, G. Getting closer: Contributions of zoo studies to research on the physiology and development of Bonobos Pan paniscus, Chimpanzees Pan troglodytes and other primates. Int. Zoo Yearb. 2018, 52, 1–14. [Google Scholar] [CrossRef]
- Janson, C.H.; Byrne, R. What wild primates know about resources: Opening up the black box. Anim. Cogn. 2007, 10, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Brando, S.I.C.A. Advances in husbandry training in marine mammal care programs. Int. J. Comp. Psychol. 2010, 23, 777–791. [Google Scholar] [CrossRef]
- Douglas, C.; Bateson, M.; Walsh, C.; Bédué, A.; Edwards, S.A. Environmental enrichment induces optimistic cognitive biases in pigs. Appl. Anim. Behav. Sci. 2012, 139, 65–73. [Google Scholar] [CrossRef]
- Enkel, T.; Gholizadeh, D.; von Bohlen Und Halbach, O.; Sanchis-Segura, C.; Hurlemann, R.; Spanagel, R.; Gass, P.; Vollmayr, B. Ambiguous-cue interpretation is biased under stress- and depression-like states in rats. Neuropsychopharmacology 2010, 35, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Clegg, I.L.K.; Delfour, F. Can we assess marine mammal welfare in captivity and in the wild? Considering the example of bottlenose dolphins. Aquat. Mamm. 2018, 44, 181–200. [Google Scholar] [CrossRef]
- Pack, A.A. The synergy of laboratory and field studies of dolphin behavior and cognition. Int. J. Comp. Psychol. 2010, 538–565. [Google Scholar] [CrossRef]
- Bateson, M.; Desire, S.; Gartside, S.E.; Wright, G.A. Agitated honeybees exhibit pessimistic cognitive biases. Curr. Biol. 2011, 21, 1070–1073. [Google Scholar] [CrossRef] [PubMed]
- Shepherdson, D.; Lewis, K.D.; Carlstead, K.; Bauman, J.; Perrin, N. Individual and environmental factors associated with stereotypic behavior and fecal glucocorticoid metabolite levels in zoo housed polar bears. Appl. Anim. Behav. Sci. 2013, 147, 268–277. [Google Scholar] [CrossRef]
- Ventre, J.; Jett, J. Killer whales, theme parks and controversy: An exploration of the evidence. In Animals and Tourism: Understanding Diverse Relationships; Markwell, K., Ed.; UK Channel View Productions: Bristol, UK, 2015; p. 328. [Google Scholar]
- Schroeder, P.; Jones, S.; Young, I.S.; Sneddon, L.U. What do zebrafish want? Impact of social grouping dominance and gender on preference for enrichment. Lab. Anim. 2014, 48, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Delfour, F.; Beyer, H. Assessing the effectiveness of environmental enrichment in bottlenose dolphins (Tursiops truncatus). Zoo Biol. 2012, 31, 137–150. [Google Scholar] [CrossRef]
- Nettle, D.; Bateson, M. The evolutionary origins of mood and its disorders. Curr. Biol. 2012, 22, R712–R721. [Google Scholar] [CrossRef] [PubMed]
- Whitham, J.C.; Wielebnowski, N. Animal-Based welfare monitoring: Using keeper ratings as an assessment tool. Zoo Biol. 2009, 28, 545–560. [Google Scholar] [CrossRef] [PubMed]
- Carlstead, K.; Mellen, J.; Kleiman, D.G. Black rhinoceros (Diceros bicornis) in U.S. zoos: I. Individual behavior profiles and their relationship to breeding success. Zoo Biol. 1999, 18, 17–34. [Google Scholar] [CrossRef]
- Pederson, A.K.; King, J.E.; Landau, V.I. Chimpanzee (Pan troglodytes) personality predicts behavior. J. Res. Pers. 2005, 39, 534–549. [Google Scholar] [CrossRef]
- Weiss, A.; King, J.E.; Perkins, L. Personality and subjective well-being in orangutans (Pongo pygmaeus and Pongo abelii). J. Pers. Soc. Psychol. 2006, 90, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Robinson, L.M.; Altschul, D.M.; Wallace, E.K.; Úbeda, Y.; Llorente, M.; Machanda, Z.; Slocombe, K.E.; Leach, M.C.; Waran, N.K.; Weiss, A. Chimpanzees with positive welfare are happier, extraverted, and emotionally stable. Appl. Anim. Behav. Sci. 2017, 90–97. [Google Scholar] [CrossRef]
- Phillips, C.J.C.; Tribe, A.; Lisle, A.; Galloway, T.K.; Hansen, K. Keepers’ rating of emotions in captive big cats, and their use in determining responses to different types of enrichment. J. Vet. Behav. Clin. Appl. Res. 2017, 20, 22–30. [Google Scholar] [CrossRef]
- Meagher, R.K. Observer ratings: Validity and value as a tool for animal welfare research. Appl. Anim. Behav. Sci. 2009, 119, 1–14. [Google Scholar] [CrossRef]
- Szokalski, M.S.; Litchfield, C.A.; Foster, W.K. What can zookeepers tell us about interacting with big cats in captivity? Zoo Biol. 2013, 32, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Claxton, A.M. The potential of the human-animal relationship as an environmental enrichment for the welfare of zoo-housed animals. Appl. Anim. Behav. Sci. 2011, 133, 1–10. [Google Scholar] [CrossRef]
- Chelluri, G.I.; Ross, S.R.; Wagner, K.E. Behavioral correlates and welfare implications of informal interactions between caretakers and zoo-housed chimpanzees and gorillas. Appl. Anim. Behav. Sci. 2013, 147, 306–315. [Google Scholar] [CrossRef]
- Vonk, J. Advances in Animal Cognition. Behav. Sci. (Basel) 2016, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Barber, J.C.E. Programmatic approaches to assessing and improving animal welfare in zoos and aquariums. Zoo Biol. 2009, 28, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Würbel, H.; Garner, J.P. Refinement of Rodent Research though Environmental Enrichment and Systematic Randomization. Available online: https://www.semanticscholar.org/paper/Refinement-of-rodent-research-through-environmental-Garner/5307822a61c90fdf354e38e697cdc1143086baab?tab=abstract (accessed on 25 June 2018).
- Baumans, V.; Van Loo, P.L.P. How to improve housing conditions of laboratory animals: The possibilities of environmental refinement. Vet. J. 2013, 195, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Poole, T. Happy animals make good science. Lab. Anim. 1997, 31, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Cronin, K.A.; Jacobson, S.L.; Bonnie, K.E.; Hopper, L.M. Studying primate cognition in a social setting to improve validity and welfare: A literature review highlighting successful approaches. PeerJ 2017, 5, e3649. [Google Scholar] [CrossRef] [PubMed]
- Koolhaas, J.M.; Korte, S.M.; De Boer, S.F.; Van Der Vegt, B.J.; Van Reenen, C.G.; Hopster, H.; De Jong, I.C.; Ruis, M.A.W.; Blokhuis, H.J. Coping styles in animals: Current status in behavior and stress- physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef]
- Hennessy, M.B.; Kaiser, S.; Sachser, N. Social buffering of the stress response: Diversity, mechanisms, and functions. Front. Neuroendocrinol. 2009, 30, 470–482. [Google Scholar] [CrossRef] [PubMed]
- Hogg, S. A review of the validity and variability of the elevated plus-maze as an animal model of anxiety. Pharmacol. Biochem. Behav. 1996, 54, 21–30. [Google Scholar] [CrossRef]
- Rodgers, R.J.; Cole, J.C. Influence of social isolation, gender, strain, and prior novelty on plus-maze behaviour in mice. Physiol. Behav. 1993, 54, 729–736. [Google Scholar] [CrossRef]
- Bateson, M.; Emmerson, M.; Ergün, G.; Monaghan, P.; Nettle, D. Opposite effects of early-life competition and developmental telomere attrition on cognitive biases in juvenile European starlings. PLoS One 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Briefer, E.F.; McElligott, A.G. Rescued goats at a sanctuary display positive mood after former neglect. Appl. Anim. Behav. Sci. 2013, 146, 45–55. [Google Scholar] [CrossRef]
- Faustino, A.I.; Oliveira, G.A.; Oliveira, R.F. Linking appraisal to behavioral flexibility in animals: Implications for stress research. Front. Behav. Neurosci. 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Rygula, R.; Papciak, J.; Popik, P. Trait pessimism predicts vulnerability to stress-induced anhedonia in rats. Neuropsychopharmacology 2013, 38, 2188–2196. [Google Scholar] [CrossRef] [PubMed]
- Clegg, I.L.K.; van Elk, C.E.; Delfour, F. Applying welfare science to bottlenose dolphins (Tursiops truncatus). Anim. Welf. 2017, 26, 165–176. [Google Scholar] [CrossRef]
- Wemelsfelder, F.; Mullan, S. Applying ethological and health indicators to practical animal welfare assessment. Rev. Sci. Tech. Off. Int. Epiz 2014, 33, 111–120. [Google Scholar] [CrossRef]


{kind=link}
| A | B | C | D | E | F | G | H | I |
|---|---|---|---|---|---|---|---|---|
| Species | Setting | Type of Cognitive Bias | Experimental Manipulation? | Task and Cues | S+/S− or S++/S+? | Tested in Isolation or Group; in Home Environment? | Link between Cognitive Bias and Affective State? | Reference |
| Common marmosets, Callithrix jacchus | Laboratory | Judgement bias | Yes: rearing context. Either: family-reared twins, family-reared animals from triplet litters where only two remain, or supplementary fed triplets. | Go/No-Go; visual cues: height of wooden tubes | S+/S− | Isolated; home | No: overall no consistent effects of rearing context | Ash and Buchanan-Smith 2016 [50] |
| Chimpanzees, Pan troglodytes | Rescue center | Judgement bias | No: individual differences | Go/No-Go; visual cues: color of paper cone | S+/S− | Isolated; home | Yes, correlation with rank, higher rank = less pessimistic | Bateson and Nettle 2015 [36] |
| Rhesus macaques. Macaca mulatta | Laboratory | Judgement bias | Yes: environmental enrichment vs. veterinary examination | Go/No-Go; visual cues: length of line on screen | S+/S− | Isolated; home | Yes: enrichment and no veterinary examination decreased pessimism | Bethell et al. 2012 [51] |
| Rhesus macaques. Macaca mulatta | Laboratory | Attention bias | Yes: environmental enrichment vs. veterinary examination | Vigilance towards aggressive images of conspecific faces measured | n/a | Isolated; not in home | Yes: veterinary examination caused avoidance of images, enrichment caused sustained vigilance | Bethell et al. 2012 [45] |
| Rhesus macaques. Macaca mulatta | Laboratory | Response- slowing | Yes: some animals underwent a veterinary examination | Speed to touch 2D shape on touchscreen next to different images of conspecific faces | n/a | Isolated; home | Yes: undergoing a veterinary examination caused slower responses when negative emotional content (images of staring conspecific faces) present | Bethell et al. 2016 [52] |
| Bottlenose dolphins, Tursiops truncatus | Zoo | Judgement bias | No: measured spontaneous social and anticipatory behaviors | Go/Go; spatial cues: position of a target | S++/S+ | Group; home | Yes: increased synchronous swimming and decreased anticipatory behavior correlated with more optimistic responses | Clegg et al. 2017 [14]; Clegg and Delfour 2018 [42] |
| Chimpanzees, Pan troglodytes | Zoo | Response- slowing | No: used spontaneous anthropogenic overhead noise from annual aircraft show | Speed to touch 2D shape on touchscreen next to different images of conspecific faces | n/a | Group; home | No: loud sound event did not seem to impact affective state | Cronin et al. 2018 [53] |
| Gorillas, Gorilla gorilla gorilla | Zoo | Response- slowing | No: used spontaneous anthropogenic overhead noise from annual aircraft show | Speed to touch 2D shape on touchscreen next to different images of conspecific faces | n/a | Group; home | No: loud sound event did not seem to impact affective state | Cronin et al. 2018 [53] |
| Japanese macaques, Macaca fuscata | Zoo | Response- slowing | No: used spontaneous anthropogenic overhead noise from annual aircraft show | Speed to touch 2D shape on touchscreen next to different images of conspecific faces | n/a | Group; home | Yes: anthropogenic noise caused slower responses when negative emotional content (images of staring conspecific faces) present | Cronin et al. 2018 [53] |
| Orange-winged parrot, Amazona amazonica | Laboratory | Attention bias | No: personality assessment using subjective ratings | Performance on a foraging task with and without an unfamiliar observer | n/a | Isolated; home | Yes: more neurotic parrots showed greater attention bias, i.e., performed worse in task when unfamiliar human present | Cussen and Mench 2014 [40] |
| Common marmosets, Callithrix jacchus | Laboratory | Judgement bias | No: handedness of animals, data taken from retrospective records | Go/No-Go; visual cues: color of lid | S+/S− | Isolated; home | Yes: left-handed marmosets were more pessimistic | Gordon and Rogers 2015 [54] |
| Grizzly bears, Ursus arctos horribilis | Research, Education and Conservation center | Judgement bias | Yes: environmental enrichment given | Go/Go; visual cues: color of boards | S++/S+ | Isolated; home | No: environmental enrichment did not seem to alter affective state | Keen et al. 2014 [55] |
| Domestic canaries, Serinus canaria | Laboratory | Judgement bias | Yes: housed singly or in pairs, and personality measured through behavior coding, | Go/No-Go; spatial cues, position of food dishes | S+/S− | Isolated; not in home | Yes: pair-housed canaries judged more optimistically (but personality did not have an effect) | Lalot et al. 2017 [56] |
| Baboons, Papio papio | Laboratory | Response-slowing | No: measured spontaneous positive, neutral, and negative valence social and solitary behaviors | Computerized visual search task | n/a | Group; home | Yes: negatively valenced behaviors slowed following performance in task | Marzouki et al. 2014 [57] |
| Gorillas, Gorilla gorilla gorilla | Zoo | Ambiguous-cue paradigm | Yes: forage given as enrichment | Visual cues: 2D shapes on a touchscreen | S+/S− | Isolated; home | No: environmental enrichment did not seem to alter affective state | McGuire et al. 2017 [15] |
| American black bear, Ursus americanus | Zoo | Ambiguous-cue paradigm | No: measured spontaneous visitor density | Visual cues: 2D shapes on a touchscreen | S+/S− | Isolated; home | No: visitor density did not seem to alter affective state | McGuire et al. 2017 [16] |
| White-lipped peccaries, Tayassu pecari | Laboratory | Judgement bias | Yes: net-trapping or a control | Go/No-Go; auditory cues: different (multi-dimensional) tones | S+/S− | Group training, isolated testing; home | Yes: net-trapping made animals more pessimistic | Nogueira et al. 2015 [58] |
| Collared peccaries, Pecari tajacu | Laboratory | Judgement bias | Yes: space restriction in interaction with environmental enrichment | Go/No-Go; auditory cues: different (multi-dimensional) tones | S+/S− | Isolated; not in home | Yes: space restriction caused more pessimistic judgements, and effects were mitigated by enrichment | Oliveira et al. 2016 [59] |
| Tufted capuchins, Cebus apella | Laboratory | Judgement bias | No: measured stereotypic behaviors | Go/Go: visual cues: length of rectangles on board | S++/S+ | Isolated; home | Yes: monkeys performing more stereotypic head twirls judged more pessimistically | Pomerantz et al. 2012 [60] |
| Tufted capuchins, Cebus apella | Laboratory | Judgement bias | No: measured rates of conspecific grooming and hierarchical rank | Go/Go: spatial cues: position of rectangular object | S++/S+ | Isolated; home | Yes: more dominant monkeys and those who received more conspecific grooming were more optimistic | Schino et al. 2016 [61] |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clegg, I.L.K. Cognitive Bias in Zoo Animals: An Optimistic Outlook for Welfare Assessment. Animals 2018, 8, 104. https://doi.org/10.3390/ani8070104
Clegg ILK. Cognitive Bias in Zoo Animals: An Optimistic Outlook for Welfare Assessment. Animals. 2018; 8(7):104. https://doi.org/10.3390/ani8070104
Chicago/Turabian StyleClegg, Isabella L. K. 2018. "Cognitive Bias in Zoo Animals: An Optimistic Outlook for Welfare Assessment" Animals 8, no. 7: 104. https://doi.org/10.3390/ani8070104
APA StyleClegg, I. L. K. (2018). Cognitive Bias in Zoo Animals: An Optimistic Outlook for Welfare Assessment. Animals, 8(7), 104. https://doi.org/10.3390/ani8070104