Effects of Photoperiod Change on Melatonin Secretion, Immune Function and Antioxidant Status of Cashmere Goats


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Animal and Experiment Design
2.3. Preparation of Blood Sample and Analysis
2.4. Total mRNA Extraction and Quality Determination
2.5. Quantitative RT-PCR Analysis
2.6. Statistical Analysis
3. Results
3.1. MLT Secretion
3.2. Immune Function
3.3. Antioxidant Status Indicators
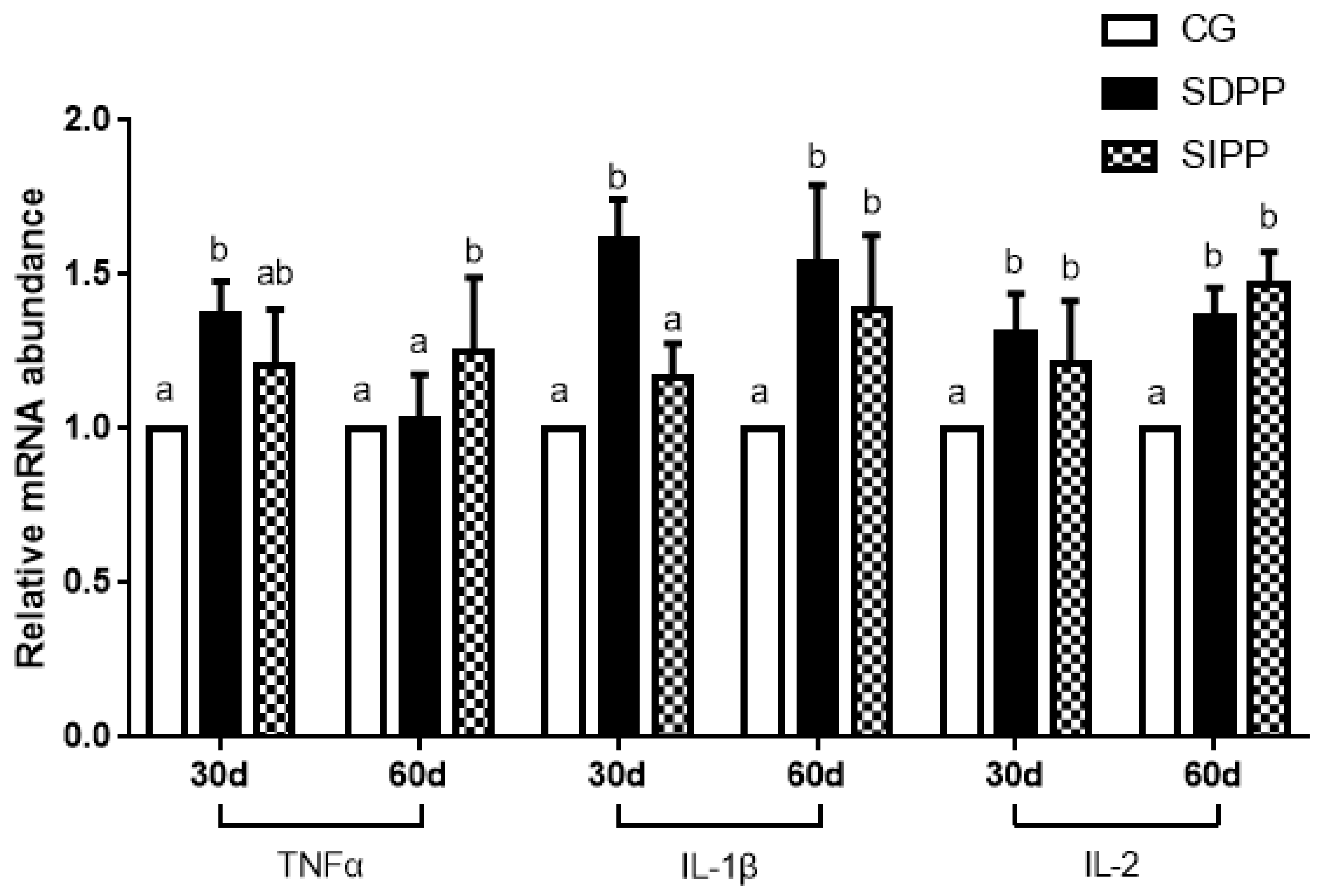
3.4. The Relative Expression of mRNA
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moore, R.Y. Neural control of pineal function in mammals and birds. J. Neural Transm. Suppl. 1978, 13, 47. [Google Scholar]
- Chakravarti, S.; Rizvi, S. Physiological effects of melatonin: Implication on human health. Biomedicine 2008, 28, 252–255. [Google Scholar]
- Li, C.; Zhou, X. Melatonin and male reproduction. Clin. Chim. Acta 2015, 446, 175–180. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Ma, T.; Shi, J.; Zhang, Z.; Wang, J.; Zhu, K.; Li, Y.; Yang, M.; Song, Y.; Liu, G. Melatonin and its receptor MT1 are involved in the downstream reaction to luteinizing hormone and participate in the regulation of luteinization in different species. J. Pineal Res. 2016, 61, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Barrett, P.; Bolborea, M. Molecular pathways involved in seasonal body weight and reproductive responses governed by melatonin. J. Pineal Res. 2012, 52, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.; Maestroni, G.; Cardinali, D.P.; Esquifino, A.I.; Perumal, S.P.; Miller, S.C. Melatonin, immune function and aging. Recent Pat. Endocr. Metab. Immune Drug Discov. 2005, 29, 2–27. [Google Scholar]
- Carmen, R.; Mayo, J.C.; Sainz, R.M.; Isaac, A.; Federico, H.; Vanesa, M.; Reiter, R.J. Regulation of antioxidant enzymes: A significant role for melatonin. J. Pineal Res. 2010, 36, 1–9. [Google Scholar]
- Amaresh Kumar, S.; Somenath, G.; Priyoneel, B.; Chandana, H. Daily variation in melatonin level, antioxidant activity and general immune response of peripheral blood mononuclear cells and lymphoid tissues of Indian goat Capra hircus during summer and winter. Indian J. Exp. Biol. 2014, 52, 467–477. [Google Scholar]
- Mayo, J.C.; Sainz, R.M.; Antolín, I.; Herrera, F.; Martin, V.; Rodriguez, C. Melatonin regulation of antioxidant enzyme gene expression. Cell Mol. Life Sci. 2002, 59, 1706–1713. [Google Scholar] [CrossRef]
- He, Y.M.; Deng, H.H.; Shi, M.H.; Bodinga, B.M.; Chen, H.L.; Han, Z.S.; Jiang, Z.L.; Li, Q.W. Melatonin modulates the functions of porcine granulosa cells via its membrane receptor MT2 in vitro. Anim. Reprod. Sci. 2016, 172, 164–172. [Google Scholar] [CrossRef]
- Dun-Xian, T.; Manchester, L.C.; Xiaoyan, L.; Rosales-Corral, S.A.; Dario, A.C.; Reiter, R.J. Mitochondria and chloroplasts as the original sites of melatonin synthesis: A hypothesis related to melatonin’s primary function and evolution in eukaryotes. J. Pineal Res. 2013, 54, 127–138. [Google Scholar]
- Reiter, R.J. Oxidative processes and antioxidative defense mechanisms in the aging brain. FASEB J. 1995, 9, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, G.; CosKun, S.; Erbas, D.; Hasanoglu, E. The effect of melatonin on liver superoxide dismutase activity, serum nitrate and thyroid hormone levels. Jap. J. Physiol. 2000, 50, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ng, T.B. Effect of pineal indoles on activities of the antioxidant defense enzymes superoxide dismutase, catalase, and glutathione reductase, and levels of reduced and oxidized glutathione in rat tissues. Biochem. Cell Biol. 2000, 78, 447. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, P.K.; Ishwar, A.K.; Kumar, R.; Niyogi, D.; Kumar, M. Effect of exogenous melatonin and different photoperiods on oxidative status and antioxidant enzyme activity in Chhotanagpuri ewe. Vet. World 2018, 11, 130–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baydas, G.; Gursu, M.F.; Yilmaz, S.; Canpolat, S.; Yasar, A.; Cikim, G.; Canatan, H. Daily rhythm of glutathione peroxidase activity, lipid peroxidation and glutathione levels in tissues of pinealectomized rats. Neurosci. Lett. 2002, 323, 195–198. [Google Scholar] [CrossRef]
- Lissoni, P.; Bolis, S.; Brivio, F.; Fumagalli, L. A phase II study of neuroimmunotherapy with subcutaneous low-dose IL-2 plus the pineal hormone melatonin in untreatable advanced hematologic malignancies. Anticancer Res. 2000, 20, 2103–2105. [Google Scholar] [PubMed]
- Ma, N.; Abaker, J.A.; Bilal, M.S.; Dai, H.; Shen, X. Sodium butyrate improves antioxidant stability in sub-acute ruminal acidosis in dairy goats. BMC Vet. Res. 2018, 14, 275. [Google Scholar] [CrossRef]
- Yao, X.; Zhang, G.; Guo, Y.; Eisamahy, M.; Wang, S.; Wan, Y.; Le, H.; Liu, Z.; Feng, W.; Zhang, Y. Vitamin D receptor expression and potential role of vitamin D on cell proliferation and steroidogenesis in goat ovarian granulosa cells. Theriogenology 2017, 102, 162. [Google Scholar] [CrossRef]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. NAR 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Zhang, Y.S. Effect of Artemisia Ordosica Krasch on Antioxidant and Immune Function in Inner Mongolia White Cashmere Goats. Master’s Thesis, Inner Mongolia Agricultural University, Inner Mongolia, China, 2018. [Google Scholar]
- Liu, J.H.; Xu, T.T.; Liu, Y.J.; Zhu, W.Y.; Mao, S.Y. A high-grain diet causes massive disruption of ruminal epithelial tight junctions in goats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R232–R241. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Martin, G.B.; Liu, S.; Shi, B.; Guo, X.; Zhao, Y.; Yan, S. The mechanism through which dietary supplementation with heated linseed grain increases n-3 long-chain polyunsaturated fatty acid concentration in subcutaneous adipose tissue of cashmere kids. J. Anim. Sci. 2019, 97, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034. [Google Scholar] [CrossRef] [PubMed]
- Hastings, M.H.; Maywood, E.S.; Reddy, A.B. Two decades of circadian time. J. Neuroendocrinol. 2008, 20, 812–819. [Google Scholar] [CrossRef]
- Maronde, E.; Stehle, J.H. The mammalian pineal gland: Known facts, unknown facets. Trends Endocrinol. Metab. 2007, 18, 142–149. [Google Scholar] [CrossRef]
- Sweeney, T.; Donovan, A.; Roche, J.F.; O’Callaghan, D. Variation in the ability of a long day followed by a short day photoperiod signal to initiate reproductive activity in ewes at different times of the year. J. Reprod. Fertil. 1997, 109, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Yellon, S.M.; Foster, D.L. Alternate photoperiods time puberty in the female lamb. Endocrinology 1985, 116, 2090. [Google Scholar] [CrossRef]
- Rao, G.N.; Berk, B.C. Active oxygen species stimulate vascular smooth muscle cell growth and proto-oncogene expression. Circul. Res. 1992, 70, 593–599. [Google Scholar] [CrossRef]
- Yun-Zhong, F.; Sheng, Y.; Guoyao, W. Free radicals, antioxidants, and nutrition. Nutrition 2002, 18, 872–879. [Google Scholar]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Michiels, C.; Raes, M.; Toussaint, O.; Remacle, J. Importance of SE-glutathione peroxidase, catalase, and CU/ZN-SOD for cell survival against oxidative stress. Free Radic. Biol. Med. 1994, 17, 235–248. [Google Scholar] [CrossRef]
- Albarrán, M.T.; López-Burillo, S.; Pablos, M.I.; Reiter, R.J.; Agapito, M.T. Endogenous rhythms of melatonin, total antioxidant status and superoxide dismutase activity in several tissues of chick and their inhibition by light. J. Pineal Res. 2010, 30, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Mauricio, D.-M.; Rolando, H.-M.; Jorge, S.; Vctoria, C.d.S. Day-night cycle of lipid peroxidation in rat cerebral cortex and their relationship to the glutathione cycle and superoxide dismutase activity. Neuroscience 1985, 16, 859–863. [Google Scholar]
- Khizhkin, E.A.; IIyukha, V.A.; Vinogradova, A.; Antonova, E.P.; Morozov, A.V. Circadian Rhythms of Antioxidant Enzyme’s Activity in Young and Adult Rats under Light Deprivation Conditions. Adv. Gerontol. 2018, 8, 328–338. [Google Scholar] [CrossRef]
- Mates, J. Effects of antioxidant enzymes in the molecular control of reactive oxygen species toxicology. Toxicology 2000, 153, 83–104. [Google Scholar] [CrossRef]
- Stana, L.G.; Muselin, F.; Cretescu, I.; Gravila, C.; Cimpoes, S.; Trif, A. The influence of Sempervivum tectorum and melatonin administration on erythrocyte catalase in rats exposed to aluminium sulphate. Lucr. Stiintifice Zooteh. Si Biotehnol. 2012, 45, 133–136. [Google Scholar]
- Mirunalini, S.; Karthishwaran, K.; Dhamodharan, G.; Mohan, S. Melatonin attenuates lipid peroxidation and enhances circulatory antioxidants during mammary carcinogenesis in rats. J. Biochem. Technol. 2010, 2, 171–174. [Google Scholar]
- Mukherjee, A.; Haldar, C.; Vishwas, D.K. Melatonin prevents dexamethasone-induced testicular oxidative stress and germ cell apoptosis in golden hamster, Mesocricetus auratus. Andrologia 2015, 47, 920–931. [Google Scholar] [CrossRef]
- Mustafa, O.; Mehmet, O.; Yasemin Gulcan, K.; Bekir, O.; Bulent, U.; Nail, E.; Mehmet, Y.; Seref, D.; Bulent, K.; Cengizhan, A. Efficacy of melatonin, mercaptoethylguanidine and 1400W in doxorubicin- and trastuzumab-induced cardiotoxicity. J. Pineal Res. 2015, 50, 89–96. [Google Scholar]
- Pablos, M.I.; Reiter, R.J.; Ortiz, G.G.; Guerrero, J.M.; Agapito, M.T.; Chuang, J.-I.; Sewerynek, E. Rhythms of glutathione peroxidase and glutathione reductase in brain of chick and their inhibition by light. Neurochem. Int. 1998, 32, 69–75. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, Z.; Qin, Z.; Yan, S.; Shi, B. Effects of chitosan addition on growth performance, diarrhoea, anti-oxidative function and serum immune parameters of weaned piglets. S. Afr. J. Anim. Sci. 2018, 48, 142–150. [Google Scholar] [CrossRef]
- Subramanian, P.; Mirunalini, S.; Pandi-Perumal, S.R.; Trakht, I.; Cardinali, D.P. Melatonin treatment improves the antioxidant status and decreases lipid content in brain and liver of rats. Eur. J. Pharmacol. 2007, 571, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Livrea, M.A.; Tesoriere, L.; D’Arpa, D.; Morreale, M. Reaction of Melatonin With Lipoperoxyl Radicals in Phospholipid Bilayers. Free Radic. Biol. Med. 1997, 23, 706–711. [Google Scholar] [CrossRef]
- Shilpa Chakravarty, S.I.R. Day and Night GSH and MDA Levels in Healthy Adults and Effects of Different Doses of Melatonin on These Parameters. Int. J. Cell Biol. 2011, 2011, 404591. [Google Scholar] [CrossRef] [PubMed]
- Bayir, A.; Kara, H.; Koylu, O.; Kocabaş, R.; Ak, A. The effects of melatonin on erythrocyte and brain cholinesterase, nitric oxide and MDA levels in acute organophosphate toxicity. Toxicol. Lett. 2011, 205, S227. [Google Scholar] [CrossRef]
- Verma, A.K.; Singh, S.; Rizvi, S.I. Redox homeostasis in a rodent model of circadian disruption: Effect of melatonin supplementation. Gen. Comp. Endocr. 2019, 280, 97–103. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Eduardo, E.Z.; Zhou, Z.; Russel, J.R. Melatonin as a Potent and Inducible Endogenous Antioxidant: Synthesis and Metabolism. Molecules 2015, 20, 18886–18906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Li, S.; Zhou, Y.; Meng, X.; Zhang, J.-J.; Xu, D.-P.; Li, H.-B. Melatonin for the prevention and treatment of cancer. Oncotarget 2017, 8, 39896. [Google Scholar] [CrossRef]
- Xie, Z.; Chen, F.; Li, W.A.; Geng, X.; Li, C.; Meng, X.; Feng, Y.; Liu, W.; Yu, F. A review of sleep disorders and melatonin. Neurol. Res. 2017, 39, 1–7. [Google Scholar] [CrossRef]
- Chen, L.Y.; Renn, T.Y.; Liao, W.C.; Mai, F.D.; Ho, Y.J.; Hsiao, G.; Lee, A.W.; Chang, H.M. Melatonin successfully rescues hippocampal bioenergetics and improves cognitive function following drug intoxication by promoting Nrf2-ARE signaling activity. J. Pineal Res. 2017, 63, e12417. [Google Scholar] [CrossRef] [PubMed]
- Janjetovic, Z.; Jarrett, S.G.; Lee, E.F.; Duprey, C.; Reiter, R.J.; Slominski, A.T. Melatonin and its metabolites protect human melanocytes against UVB-induced damage: Involvement of NRF2-mediated pathways. Sci. Rep. 2017, 7, 1274. [Google Scholar] [CrossRef] [PubMed]
- Niture, S.K.; Kaspar, J.W.; Shen, J.; Jaiswal, A.K. Nrf2 signaling and cell survival. Toxicol. Appl. Pharmacol. 2010, 244, 37–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carolyn, S.; Telen, M.J.; Jen-Tsan, C. microRNA miR-144 modulates oxidative stress tolerance and associates with anemia severity in sickle cell disease. Blood 2010, 116, 4338–4348. [Google Scholar] [Green Version]
- Ding, K.; Wang, H.; Xu, J.; Li, T.; Zhang, L.; Ding, Y.; Zhu, L.; He, J.; Zhou, M. Melatonin stimulates antioxidant enzymes and reduces oxidative stress in experimental traumatic brain injury: The Nrf2–ARE signaling pathway as a potential mechanism. Free Radic. Biol. Med. 2014, 73, 1–11. [Google Scholar] [CrossRef]
- Negrette, B.; Bonilla, E.; Valero, N.; Pons, H.; Tamayo, J.G.; Chacín-Bonilla, L.; Medina-Leendertz, S.; Añez, F. Melatonin Treatment Enhances the Efficiency of Mice Immunization with Venezuelan Equine Encephalomyelitis Virus TC-83. Neurochem. Res. 2001, 26, 767–770. [Google Scholar] [CrossRef]
- Akbulut, K.; Gonul, B.; Akbulut, H. The effects of melatonin on humoral immune responses of young and aged rats. Immunol. Investig. 2001, 30, 17–20. [Google Scholar] [CrossRef]
- Demas, G.E.; Nelson, R.J. Exogenous melatonin enhances cell-mediated, but not humoral, immune function in adult male deer mice (Peromyscus maniculatus). J. Biol. Rhythms 1998, 13, 245–252. [Google Scholar] [CrossRef]
- Yin, Y.L.; Tang, Z.R.; Sun, Z.H.; Liu, Z.Q.; Li, T.J.; Huang, R.L.; Ruan, Z.; Deng, Z.Y.; Gao, B.; Chen, L.X. Effect of Galacto-mannan-oligosaccharides or Chitosan Supplementation on Cytoimmunity and Humoral Immunity in Early-weaned Piglets. Asian Australas. J. Anim. Sci. 2008, 21, 723–731. [Google Scholar] [CrossRef]
- Pioli, C.; Caroleo, M.C.; Nistico, G.; Doriac, G. Melatonin increases antigen presentation and amplifies specific and non specific signals for T-cell proliferation. Int. J. Immunopharmacol. 1993, 15, 463. [Google Scholar] [CrossRef]
- Maestroni, G.J. The immunoneuroendocrine role of melatonin. J. Pineal Res. 2010, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Habbal, O.A.; Al-Jabri, A.A. Circadian rhythm and the immune response: A review. Int. Rev. Immunol. 2009, 28, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Naidu, K.S.; Morgan, L.W.; Bailey, M.J. Inflammation in the avian spleen: Timing is everything. BMC Mol. Biol. 2010, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, M.K.; Vaughan, G.M.; Reiter, R.J. Effect of ovariectomy and constant dark on the weight of reproductive and certain other organs in the female vole, Microtus montanus. J. Reprod. Fertil. 1973, 32, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Haldar, C.; Singh, R.; Guchhait, P. Relationship between the annual rhythms in melatonin and immune system status in the tropical palm squirrel, Funambulus pennanti. Chronobiol. Int. 2001, 18, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Depres-Brummer, P.; Bourin, P.; Pages, N.; Metzger, G.; Lévi, F. Persistent T lymphocyte rhythms despite suppressed circadian clock outputs in rats. Am. J. Physiol. 1997, 273, 1891–1899. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, M.K.; Reiter, R.J. Transient hypertrophy of the ventral prostate and coagulating glands and accelerated thymic involution following pinealectomy in the mouse. Tex. Rep. Biol. Med. 1971, 29, 579–586. [Google Scholar]
- Mckinney, T.D.; Vaughan, M.K.; Reiter, R.J. Pineal influence on intermale aggression in adult house mice. Physiol. Behav. 1975, 15, 213–216. [Google Scholar] [CrossRef]
- Liu, F.; Ng, T.B.; Fung, M.C. Pineal indoles stimulate the gene expression of immunomodulating cytokines. J. Neural Transm. 2001, 108, 397–405. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Sequence of Nucleotide (5′-3′) | Genebank No. | Size (bp) | |
|---|---|---|---|---|
| SOD1 | F | ATCCACTTCGAGGCAAAGGG | NM_001285550.1 | 104 |
| R | GCACTGGTACAGCCTTGTGTA | |||
| SOD2 | F | TCAATAAGGAGCAGGGACGC | XM_005684984.1 | 85 |
| R | AGCAGGGGGATAAGACCTGT | |||
| GPx1 | F | ACATTGAAACCCTGCTGTCC | XM_005695962.2 | 216 |
| R | TCATGAGGAGCTGTGGTCTG | |||
| GPx4 | F | TTCCCTTGCAACCAGTTTGG | NC_030814.1 | 105 |
| R | TCATCCATTTCCACAGAGGGT | |||
| Nrf2 | F | AGCCAGGTGAGATGGAACTG | XM_005679848.2 | 120 |
| R | CCAGACTCCCTGTTTCGCTG | |||
| CAT | F | CACTCAGGTGCGGGATTTCT | GQ_204786.1 | 159 |
| R | ATGCGGGAGCCATATTCAGG | |||
| IL-1β | F | CATGTGTGCTGAAGGCTCTC | D63351.1 | 173 |
| R | AGTGTCGGCGTATCACCTTT | |||
| IL-2 | F | TGTCTTGCATTGCACTAACTCTTGC | AF_307018.1 | 116 |
| R | CCCAAAAGCAACTGTAAATCCAGC | |||
| IL-6 | F | GGGCTGCTCCTGGTGATGACTT | HM_565937.1 | 133 |
| R | CGATGTGCTTAATGAGAGCTTCGG | |||
| TNF-α | F | CAACAGGCCTCTGGTTCAGAC | NC_030830.1 | 209 |
| R | GGACCTGCGAGTAGATGAGG | |||
| β-actin | F | ACTGGGACGACATGGAGAAGA | U39357 | 199 |
| R | GCGTACAGGGACAGCACAG | |||
| B2M | F | GGTGCTGCTTAGAGGTCTCG | NM_001009284 | 109 |
| R | ACGCTGAGTTCACTCCCAAC | |||
| YWHAZ | F | TGTAGGAGCCCGTAGGTCATCT | AY970970 | 102 |
| R | TTCTCTCTGTATTCTCGAGCCATCT |
| Day | Item | Groups | p-Value | ||
|---|---|---|---|---|---|
| CG | SDPP | SIPP | |||
| 30 d | IgG (mg/mL) | 211.8 ± 18.4 a | 264.9 ± 10.8 b | 220.7 ± 20.8 a | 0.043 |
| IgM (μg/mL) | 160.8 ± 19.6 | 158.8 ± 22.9 | 165.8 ± 28.7 | 0.631 | |
| IgA (μg/mL) | 86.97 ± 12.84 | 94.83 ± 15.86 | 88.57 ± 19.21 | 0.427 | |
| IL-1β (ng/mL) | 148.3 ± 10.9 a | 190.5 ± 22.6 b | 160.3 ± 16.3 a | 0.037 | |
| IL-2 (pg/mL) | 197.8 ± 15.4 a | 247.1 ± 13.1 b | 221.4 ± 15.5 a | 0.05 | |
| TNF-α (ng/mL) | 50.11 ± 8.43 | 53.85 ± 9.10 | 52.28 ± 7.36 | 0.533 | |
| 60 d | IgG (mg/mL) | 202.6 ± 20.9 a | 270.8 ± 30.5 b | 257.3 ± 15.4 b | 0.049 |
| IgM (μg/mL) | 158.9 ± 17.3 | 163.6 ± 21.8 | 162.6 ± 19.4 | 0.738 | |
| IgA (μg/mL) | 90.33 ± 5.34 | 86.31 ± 12.72 | 83.92 ± 20.95 | 0.421 | |
| IL-1β (ng/mL) | 155.4 ± 12.6 a | 186.4 ± 12.8 b | 179.3 ± 10.1 b | 0.05 | |
| IL-2 (pg/mL) | 220.7 ± 19.8 a | 269.3 ± 13.9 b | 261.3 ± 24.6 ab | 0.044 | |
| TNF-α (ng/mL) | 44.24 ± 9.21 | 50.34 ± 7.39 | 49.34 ± 6.35 | 0.342 | |
| Day | Item | Groups | p-Value | ||
|---|---|---|---|---|---|
| CG | SDPP | SIPP | |||
| 30 d | T-SOD (U/mL) | 125.0 ± 1.9 a | 139.1 ± 8.4 b | 126.8 ± 2.3 a | 0.05 |
| GPx (U/mL) | 142.7 ± 4.6 a | 160.9 ± 5.3 b | 137.1 ± 9.4 a | <0.01 | |
| CAT (U/mL) | 2.22 ± 0.29 a | 3.00 ± 0.23 b | 2.37 ± 0.20 a | <0.01 | |
| MDA (nmol/mL) | 2.14 ± 0.11 a | 1.46 ± 0.21 b | 2.38 ± 0.12 a | 0.047 | |
| T-AOC (U/mL) | 0.243 ± 0.014 a | 0.272 ± 0.019 b | 0.211 ± 0.023 a | <0.01 | |
| 60 d | T-SOD (U/mL) | 123.7 ± 2. 7 a | 143.8 ± 10.4 b | 136.3 ± 6.4 b | <0.01 |
| GPx (U/mL) | 146.2 ± 2.0 a | 160.1 ± 5.1 b | 168.4 ± 5.1 b | 0.05 | |
| CAT (U/mL) | 2.08 ± 0.16 a | 3.68 ± 0.14 b | 3.33 ± 0.15 b | <0.01 | |
| MDA (nmol/mL) | 2.47 ± 0.33 a | 1.45 ± 0.15 b | 1.17 ± 0.12 b | 0.023 | |
| T-AOC (U/mL) | 0.231 ± 0.037 | 0.259 ± 0.013 | 0.226 ± 0.011 | 0.772 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, C.; Xu, Y.; Shi, L.; Guo, S.; Jin, X.; Yan, S.; Shi, B. Effects of Photoperiod Change on Melatonin Secretion, Immune Function and Antioxidant Status of Cashmere Goats. Animals 2019, 9, 766. https://doi.org/10.3390/ani9100766
Mao C, Xu Y, Shi L, Guo S, Jin X, Yan S, Shi B. Effects of Photoperiod Change on Melatonin Secretion, Immune Function and Antioxidant Status of Cashmere Goats. Animals. 2019; 9(10):766. https://doi.org/10.3390/ani9100766
Chicago/Turabian StyleMao, Chenyu, Yuanqing Xu, Lulu Shi, Shiwei Guo, Xiao Jin, Sumei Yan, and Binlin Shi. 2019. "Effects of Photoperiod Change on Melatonin Secretion, Immune Function and Antioxidant Status of Cashmere Goats" Animals 9, no. 10: 766. https://doi.org/10.3390/ani9100766
APA StyleMao, C., Xu, Y., Shi, L., Guo, S., Jin, X., Yan, S., & Shi, B. (2019). Effects of Photoperiod Change on Melatonin Secretion, Immune Function and Antioxidant Status of Cashmere Goats. Animals, 9(10), 766. https://doi.org/10.3390/ani9100766