Effects of Nutritional Deprivation and Re-Alimentation on the Feed Efficiency, Blood Biochemistry, and Rumen Microflora in Yaks (Bos grunniens)

 ,
,  , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design, and Samples
2.1.1. Experiment 1—Effects of Starvation and Refeeding on Yak Growth Performance
2.1.2. Experiment 2—Effects of Starvation and Refeeding on Microbial Composition in the Rumen of Yaks
2.2. Experimental Diet
2.3. Nutrients Digestibility
2.4. Blood Biochemical Indicators
2.5. Rumen Fermentation Parameters
2.6. Polymerase Chain Reaction Amplification and Sequencing
2.7. Bioinformatics and Statistical Analysis
3. Results
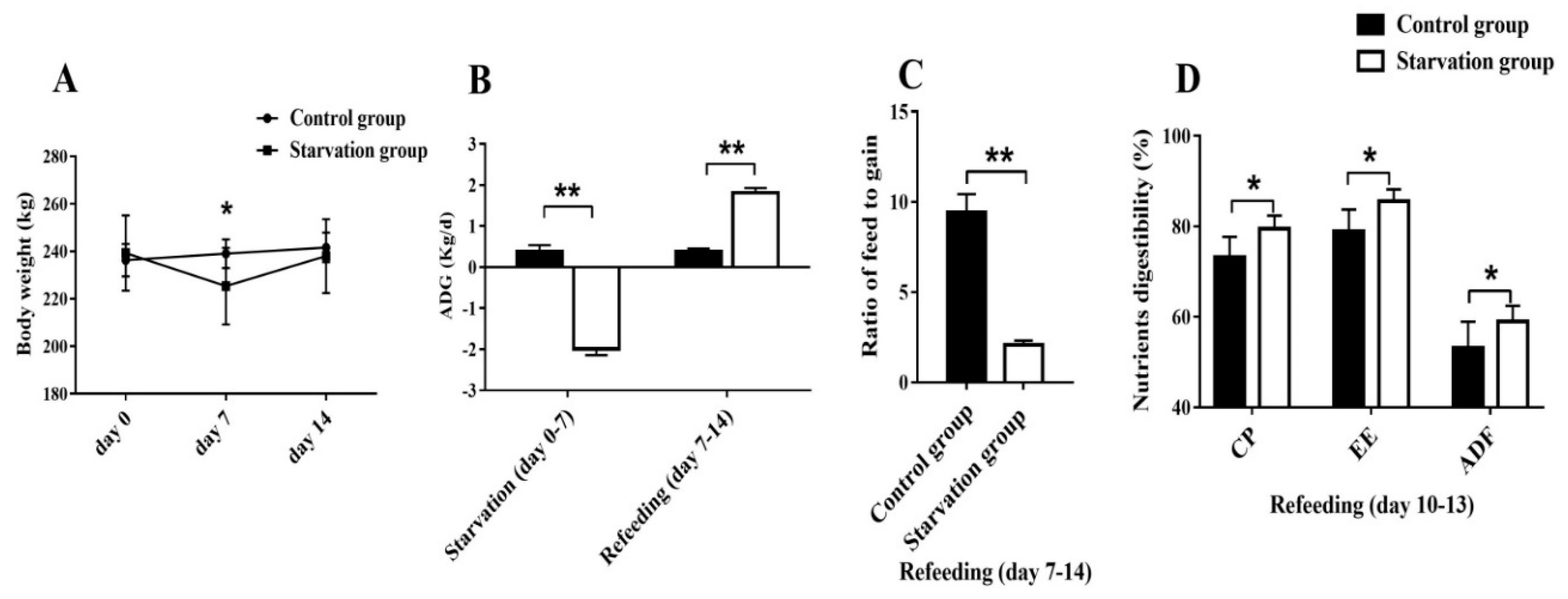
3.1. Experiment 1—Effects of Starvation and Refeeding on Yak Growth Performance
3.1.1. Growth Performance and Nutrients Digestibility during Starvation and Refeeding Periods
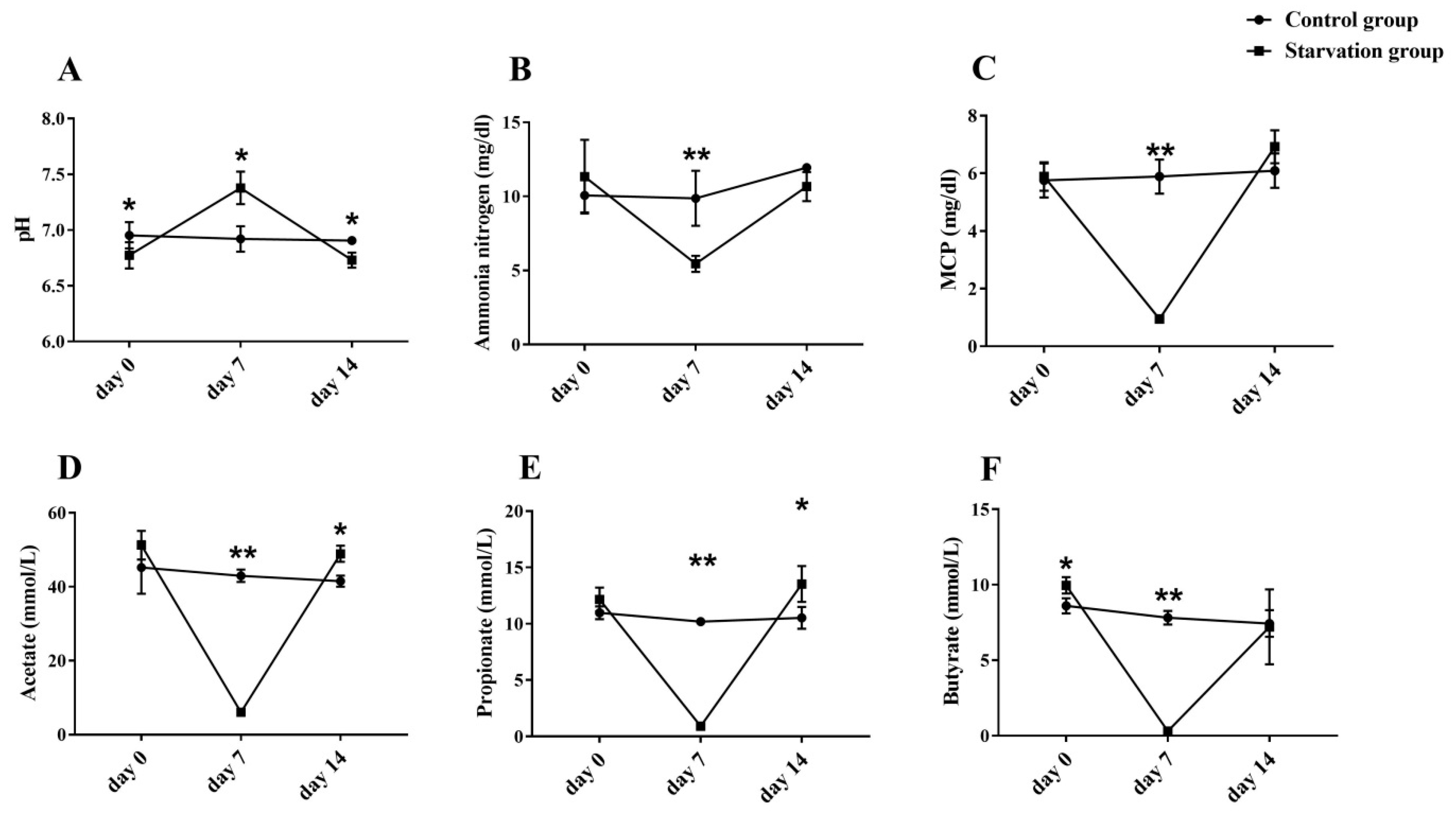
3.1.2. Blood Biochemistry during Starvation and Refeeding Periods
3.1.3. Rumen Fermentation during Starvation and Refeeding Periods
3.2. Experiment 2—Effects of Starvation and Refeeding on Microbial Composition in the Rumen of Yaks
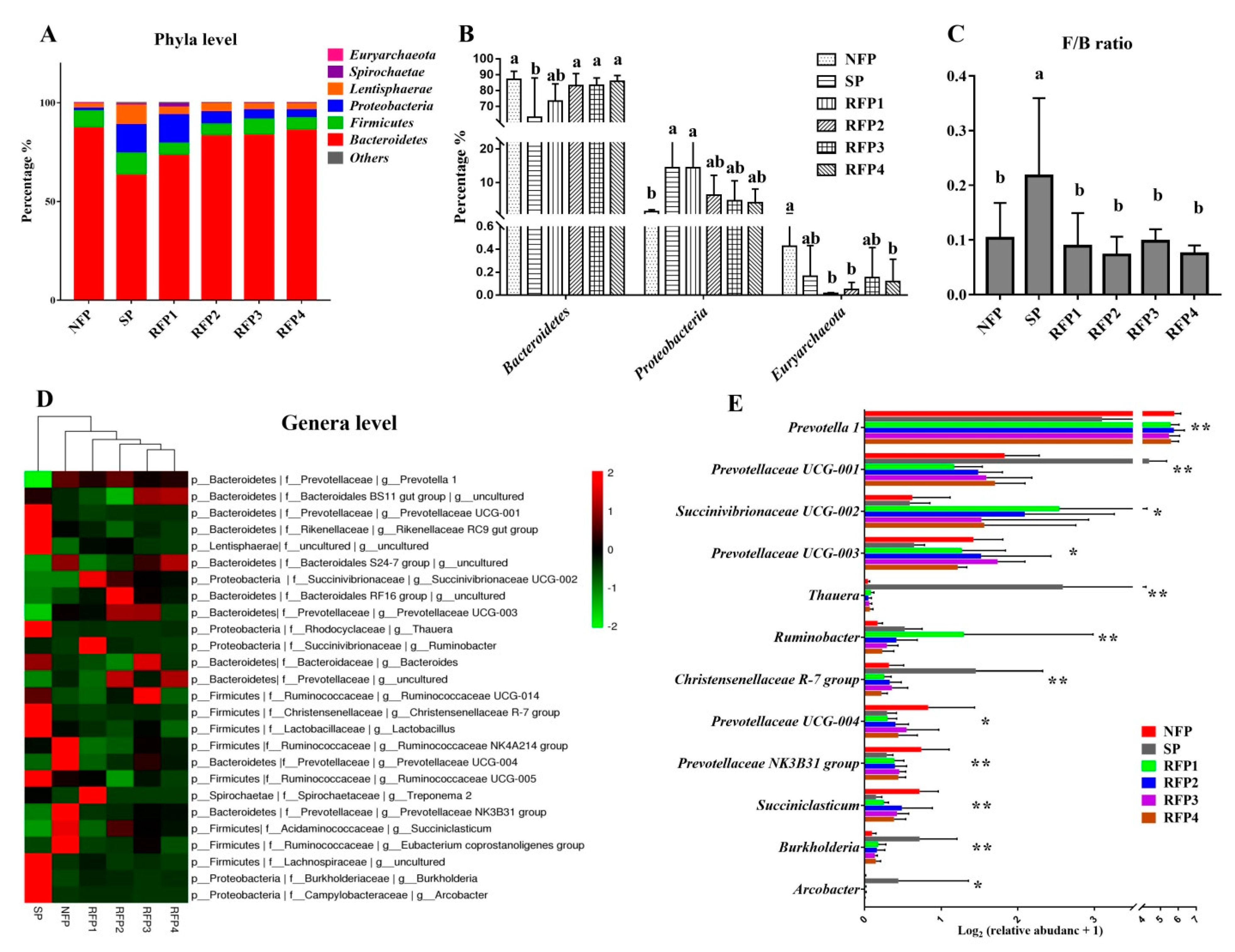
3.2.1. Rumen Fermentation Alterations throughout the Normal Feeding, Starvation and Prolonged Refeeding Periods
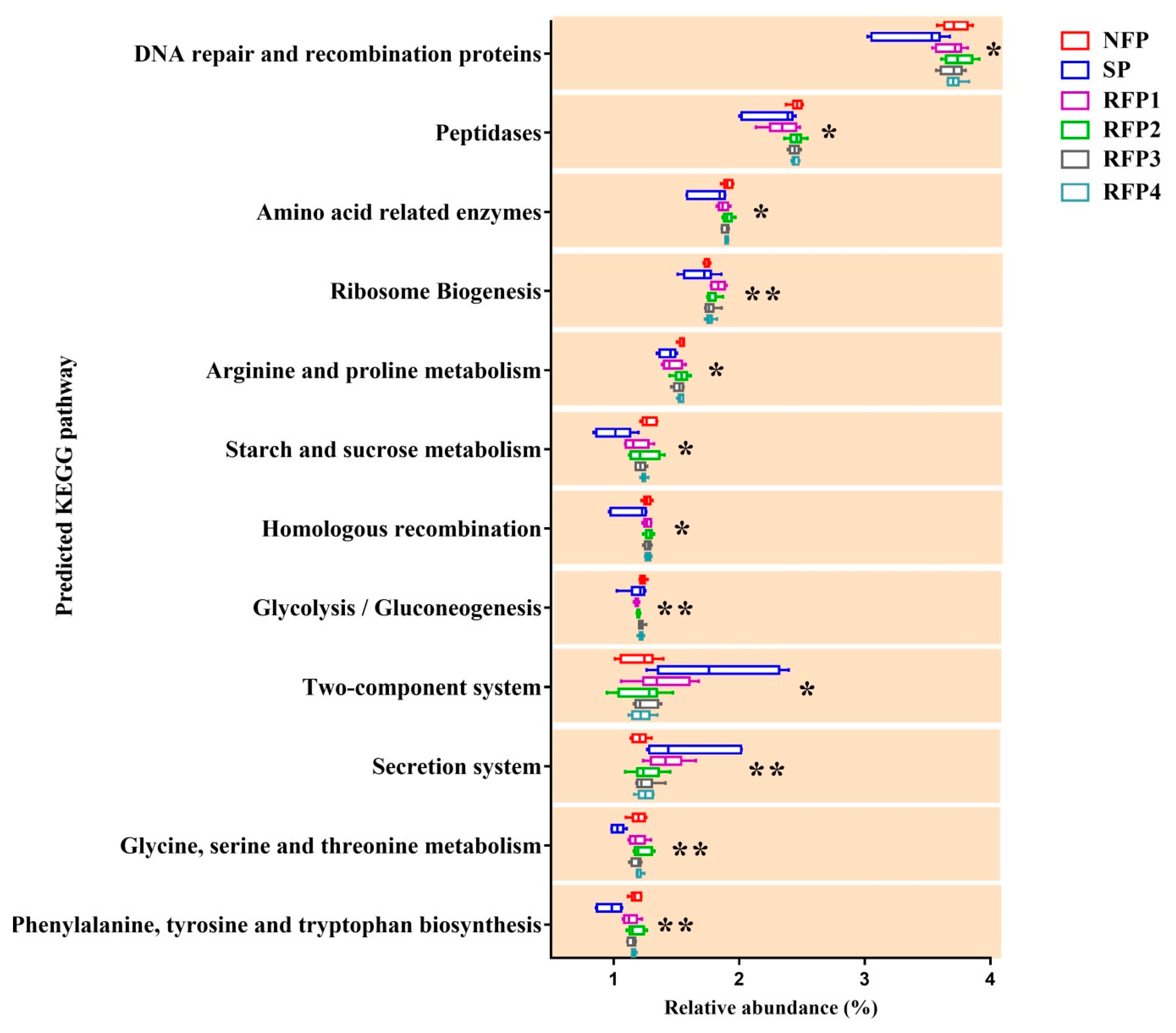
3.2.2. Rumen Microflora Alterations throughout the Normal Feeding, Starvation and Prolonged Refeeding Periods
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Xue, B.; Zhao, X.Q.; Zhang, Y.S. Seasonal changes in weight and body composition of yak grazing on alpine-meadow grassland in the Qinghai-Tibetan plateau of China. J. Anim. 2005, 83, 1908–1913. [Google Scholar] [CrossRef] [PubMed]
- Qiang, Q.; Guojie, Z.; Tao, M.; Wubin, Q.; Junyi, W.; Zhiqiang, Y.; Changchang, C.; Quanjun, H.; Jaebum, K.; Larkin, D.M. The yak genome and adaptation to life at high altitude. Nat. Genet. 2012, 44, 946–949. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Peng, Q.; Luo, X.; An, T.; Guan, J.; Wang, Z. Effects of Starvation on Lipid Metabolism and Gluconeogenesis in Yak. Asian Australas. J. Anim. Sci. 2016, 29, 1593–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tennant, B.; Malm, O.J.; Horowitz, R.E.; Levenson, S.M. Response of germfree, conventional, conventionalized and E. coli monocontaminated mice to starvation. J. Nutr. 1968, 94, 151. [Google Scholar] [CrossRef]
- Hiro-Omi, Y.; Hisae, M.; Mitsuhiro, F. Changes in body composition of germ-free and conventional chickens during starvation. Comp. Biochem. Physiol. Part A Physiol. 1992, 103, 565–568. [Google Scholar] [CrossRef]
- Crawford, P.A.; Crowley, J.R.; Nandakumar, S.; Muegge, B.D.; Costello, E.K.; Micah, H.; Rob, K.; Gordon, J.I. Regulation of myocardial ketone body metabolism by the gut microbiota during nutrient deprivation. Proc. Natl. Acad. Sci. USA 2009, 106, 11276–11281. [Google Scholar] [CrossRef] [Green Version]
- Keogh, K.; Kenny, D.A.; Waters, S.M. Gene co-expression networks contributing to the expression of compensatory growth in metabolically active tissues in cattle. Sci. Rep. 2019, 9, 6093. [Google Scholar] [CrossRef] [Green Version]
- Keogh, K.; Waters, S.M.; Kelly, A.K.; Wylie, A.R.; Sauerwein, H.; Sweeney, T.; Kenny, D.A. Feed restriction and realimentation in Holstein-Friesian bulls: II. Effect on blood pressure and systemic concentrations of metabolites and metabolic hormones. J. Anim. Sci. 2015, 93, 3590. [Google Scholar] [CrossRef]
- Margrethe, T.; Sandra, S.; Byrne, D.V. Sensory profiling of textural properties of meat from dairy cows exposed to a compensatory finishing strategy. Meat Sci. 2011, 87, 73–80. [Google Scholar] [CrossRef]
- Hu, R.; Zou, H.; Wang, Z.; Cao, B.; Peng, Q.; Jing, X.; Wang, Y.; Shao, Y.; Pei, Z.; Zhang, X.; et al. Nutritional Interventions Improved Rumen Functions and Promoted Compensatory Growth of Growth-Retarded Yaks as Revealed by Integrated Transcripts and Microbiome Analyses. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Keogh, K.; Kenny, D.A.; Kelly, A.K.; Waters, S.M. Insulin secretion and signaling in response to dietary restriction and subsequent re-alimentation in cattle. Physiol. Genom. 2015, 47, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Mullins, Y.; Keogh, K.; Kenny, D.; Waters, S. Immunological status of cattle during dietary restriction and subsequent compensatory growth. J. Anim. Sci. 2018, 96, 101–102. [Google Scholar] [CrossRef]
- McCabe, M.; Cormican, P.; Keogh, K.; O’Connor, A.; O’Hara, E.; Palladino, R.; Kenny, D.; Waters, S. Illumina MiSeq Phylogenetic Amplicon Sequencing Shows a Large Reduction of an Uncharacterised Succinivibrionaceae and an Increase of the Methanobrevibacter gottschalkii Clade in Feed Restricted Cattle. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Colucci, P.E.; Macleod, G.K.; Grovum, W.L. Comparative digestion in sheep and cattle fed different forage to concentrate ratios at high and low intakes. J. Dairy Sci. 1989, 72, 1774–1785. [Google Scholar] [CrossRef]
- Shen, J.; Chai, Z.; Song, L.; Liu, J.; Wu, Y. Insertion depth of oral stomach tubes may affect the fermentation parameters of ruminal fluid collected in dairy cows. J. Dairy Sci. 2012, 95, 5978–5984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belanche, A.; de la Fuente, G.; Newbold, C.J. Effect of progressive inoculation of fauna-free sheep with holotrich protozoa and total-fauna on rumen fermentation, microbial diversity and methane emissions. FEMS Microbiol. Ecol. 2015, 91. [Google Scholar] [CrossRef] [PubMed]
- Moallem, U.; Lehrer, H.; Livshitz, L.; Zachut, M.; Yakoby, S. The effects of live yeast supplementation to dairy cows during the hot season on production, feed efficiency, and digestibility. J. Dairy Sci. 2009, 92, 343–351. [Google Scholar] [CrossRef]
- Stewart, C.S.; Duncan, S.H. The Effect of Avoparcin on Cellulolytic Bacteria of the Ovine Rumen. Microbiology 1985, 131, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Makkar, H.P.; Becker, K. Purine quantification in digesta from ruminants by spectrophotometric and HPLC methods. Br. J. Nutr. 1999, 81, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Tanja, M.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Lu, Z.; Xu, Z.; Chen, Z.; Shen, Z. Associations among dietary non-fiber carbohydrate, ruminal microbiota and epithelium G-protein-coupled receptor, and histone deacetylase regulations in goats. Microbiome 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.; Vega Thurber, R.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Kirton, A.H.; Paterson, D.J.; Duganzich, D.M. Effect of pre-slaughter starvation in cattle. J. Anim. Sci. 1972, 34, 555–559. [Google Scholar] [CrossRef]
- Shaoyu, Z.; Zhisheng, W.; Quanhui, P.; Bai, X.; Lizhi, W.; Huawei, Z.; Rui, H.; Yixiao, Z.; Xinyu, Z.; Jianzhong, L. Effects of Starvation and Refeeding on Growth Performance, Nutrient Apparent Digestibility and Serum Indices of Yaks. Chin. J. Anim. Nutr. 2018, 30, 4991–5001. [Google Scholar]
- Bergman, E.N. Energy Contributions of Volatile Fatty-Acids from the Gastrointestinal-Tract in Various Species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef]
- Canfield, R.W.; Butler, W.R. Energy balance and pulsatile LH secretion in early postpartum dairy cattle. Domest. Anim. Endocrinol. 1990, 7, 323–330. [Google Scholar] [CrossRef]
- Ryan, W.J. Compensatory growth in cattle and sheep. Nutr. Abstr. Rev. 1990, 60, 653–664. [Google Scholar]
- Wilson, P.; Osbourn, D. Compensatory growth after undernutrition in mammals and birds. Biol. Rev. Camb. Philos. Soc. 1960, 35, 324–363. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Han, I.K.; Woo, J.-H.; Lee, H.J.; Jang, K.; Myung, K.H.; Sun-young, K. Compensatory Growth in Dairy Heifers: The Effect of a Compensatory Growth Pattern on Growth Rate and Lactation Performance. J. Dairy Sci. 1997, 80, 519–524. [Google Scholar] [CrossRef]
- Yambayamba, E.S.; Price, M.A.; Foxcroft, G.R. Hormonal status, metabolic changes, and resting metabolic rate in beef heifers undergoing compensatory growth. J. Anim. Sci. 1996, 74, 57–69. [Google Scholar] [CrossRef]
- Fox, D.G.; Johnson, R.R.; Preston, R.L.; Dockerty, T.R.; Klosterman, E.W. Protein and energy utilization during compensatory growth in beef cattle. J. Anim. Sci. 1972, 9, 424–434. [Google Scholar] [CrossRef]
- Ryan, W.J.; Williams, I.H.; Moir, R.J. Compensatory growth in sheep and cattle. II. Changes in body composition and tissue weights. Aust. J. Agric. Res. 1993, 44, 1623–1633. [Google Scholar] [CrossRef]
- Ozutsumi, Y.; Tajima, K.; Takenaka, A.; Itabashi, H. Real-Time PCR Detection of the Effects of Protozoa on Rumen Bacteria in Cattle. Curr. Microbiol. 2006, 52, 158–162. [Google Scholar] [CrossRef]
- Johnson, D.E.; Ward, G.M. Estimates of animal methane emissions. Environ. Monit. Assess. 1996, 42, 133–141. [Google Scholar] [CrossRef]
- Jiandui, M. Dynamics in Rumen and Distribution along Gastrointestinal Tracts of Bacteria and Methanogen in Yak. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2016. [Google Scholar]
- Emerson, E.L.; Weimer, P.J. Fermentation of model hemicelluloses by Prevotella strains and Butyrivibrio fibrisolvens in pure culture and in ruminal enrichment cultures. Appl. Microbiol. Biotechnol. 2017, 101, 4269–4278. [Google Scholar] [CrossRef]
- Watanabe, Y.; Nagai, F.; Morotomi, M. Characterization of Phascolarctobacterium succinatutens sp. nov., an asaccharolytic, succinate-utilizing bacterium isolated from human feces. Appl. Environ. Microbiol. 2012, 78, 511–518. [Google Scholar] [CrossRef]
- Binbin, L.; Yuejian, M.; Linda, B.; Bakken, L.R.; Asa, F.R. Strains in the genus Thauera exhibit remarkably different denitrification regulatory phenotypes. Environ. Microbiol. 2013, 15, 2816–2828. [Google Scholar] [CrossRef]
- Pope, P.B.; Smith, W.; Denman, S.E.; Tringe, S.G.; Barry, K.; Hugenholtz, P.; Mcsweeney, C.S.; Mchardy, A.C.; Morrison, M. Isolation of Succinivibrionaceae implicated in low methane emissions from Tammar wallabies. Science 2011, 333, 646–648. [Google Scholar] [CrossRef] [PubMed]
- Hamlin, L.J.; Hungate, R.E. Culture and physiology of a starch-digesting bacterium (Bacteroides amylophilus n. sp.) from the bovine rumen. J. Bacteriol. 1956, 72, 548–554. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Content | |
|---|---|---|
| Ingredients | Corn | 20.30 |
| Wheat bran | 6.30 | |
| Soybean meal | 2.80 | |
| Rapeseed meal | 3.50 | |
| Distilled grains | 35.00 | |
| Rice straw | 30.00 | |
| CaHPO4 | 0.77 | |
| CaCO3 | 0.04 | |
| NaHCO3 | 0.42 | |
| NaCl | 0.52 | |
| Premix 1 | 0.35 | |
| Total | 100.00 | |
| Nutrient levels | Dry Matter (DM) | 90.73 |
| NEmf (MJ/kg) 2 | 5.10 | |
| Crude Protein (CP) | 12.56 | |
| Neutral Detergent Fiber (NDF) | 54.14 | |
| Acid Detergent Fiber (ADF) | 41.05 | |
| Ca | 0.73 | |
| Total phosphorus (TP) | 0.40 | |
| Items | Periods | Groups | SEM | p-Value | |
|---|---|---|---|---|---|
| Control Group | Starvation Group | ||||
| GLU (mmol/L) | Day 0 | 3.89 | 3.94 | 0.48 | 0.779 |
| Day 7 | 3.84 a | 3.19 b | 0.53 | 0.029 | |
| Day 14 | 3.63 | 3.46 | 0.47 | 0.409 | |
| BHBA (mmol/L) | Day 0 | 0.28 | 0.28 | 0.06 | 0.719 |
| Day 7 | 0.26 b | 0.39 a | 0.08 | 0.01 | |
| Day 14 | 0.25 | 0.24 | 0.05 | 0.855 | |
| TG (mmol/L) | Day 0 | 0.28 | 0.26 | 0.06 | 0.506 |
| Day 7 | 0.28 a | 0.17 b | 0.04 | 0.001 | |
| Day 14 | 0.25 | 0.19 | 0.06 | 0.056 | |
| TC (mmol/L) | Day 0 | 1.18 | 1.15 | 0.41 | 0.865 |
| Day 7 | 1.26 | 1.46 | 0.44 | 0.319 | |
| Day 14 | 1.11 | 1.25 | 0.38 | 0.425 | |
| NEFA (mmol/L) | Day 0 | 0.23 | 0.23 | 0.10 | 0.979 |
| Day 7 | 0.18 b | 1.01 a | 0.31 | 0.001 | |
| Day 14 | 0.26 | 0.22 | 0.10 | 0.675 | |
| BUN (mmol/L) | Day 0 | 4.27 | 4.01 | 0.28 | 0.072 |
| Day 7 | 4.44 a | 3.63 b | 0.27 | 0.001 | |
| Day 14 | 4.49 a | 2.67 b | 0.46 | 0.000 | |
| TP (g/L) | Day 0 | 64.18 | 65.55 | 7.96 | 0.692 |
| Day 7 | 63.53 | 61.20 | 8.42 | 0.527 | |
| Day 14 | 64.10 | 66.25 | 11.05 | <0.000 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, H.; Hu, R.; Wang, Z.; Shah, A.M.; Zeng, S.; Peng, Q.; Xue, B.; Wang, L.; Zhang, X.; Wang, X.; et al. Effects of Nutritional Deprivation and Re-Alimentation on the Feed Efficiency, Blood Biochemistry, and Rumen Microflora in Yaks (Bos grunniens). Animals 2019, 9, 807. https://doi.org/10.3390/ani9100807
Zou H, Hu R, Wang Z, Shah AM, Zeng S, Peng Q, Xue B, Wang L, Zhang X, Wang X, et al. Effects of Nutritional Deprivation and Re-Alimentation on the Feed Efficiency, Blood Biochemistry, and Rumen Microflora in Yaks (Bos grunniens). Animals. 2019; 9(10):807. https://doi.org/10.3390/ani9100807
Chicago/Turabian StyleZou, Huawei, Rui Hu, Zhisheng Wang, Ali Mujtaba Shah, Shaoyu Zeng, Quanhui Peng, Bai Xue, Lizhi Wang, Xiangfei Zhang, Xueying Wang, and et al. 2019. "Effects of Nutritional Deprivation and Re-Alimentation on the Feed Efficiency, Blood Biochemistry, and Rumen Microflora in Yaks (Bos grunniens)" Animals 9, no. 10: 807. https://doi.org/10.3390/ani9100807