Serum Concentrations of AMH and E2 and Ovarian and Uterine Traits in Gilts



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Ethics
2.2. Blood Samples and Assays
2.3. Assessment of Uterine and Ovarian Development
2.4. Histological Preparation and Analysis
2.5. Statistical Analysis
3. Results
3.1. Serum Hormone Levels, Carcass Traits and Ovarian Properties
3.2. Correlations between Ovarian Hormones and Uterine and Ovarian Traits
3.3. Intra- and Inter-Hormone Correlations
3.4. Intra-Uterus Correlations
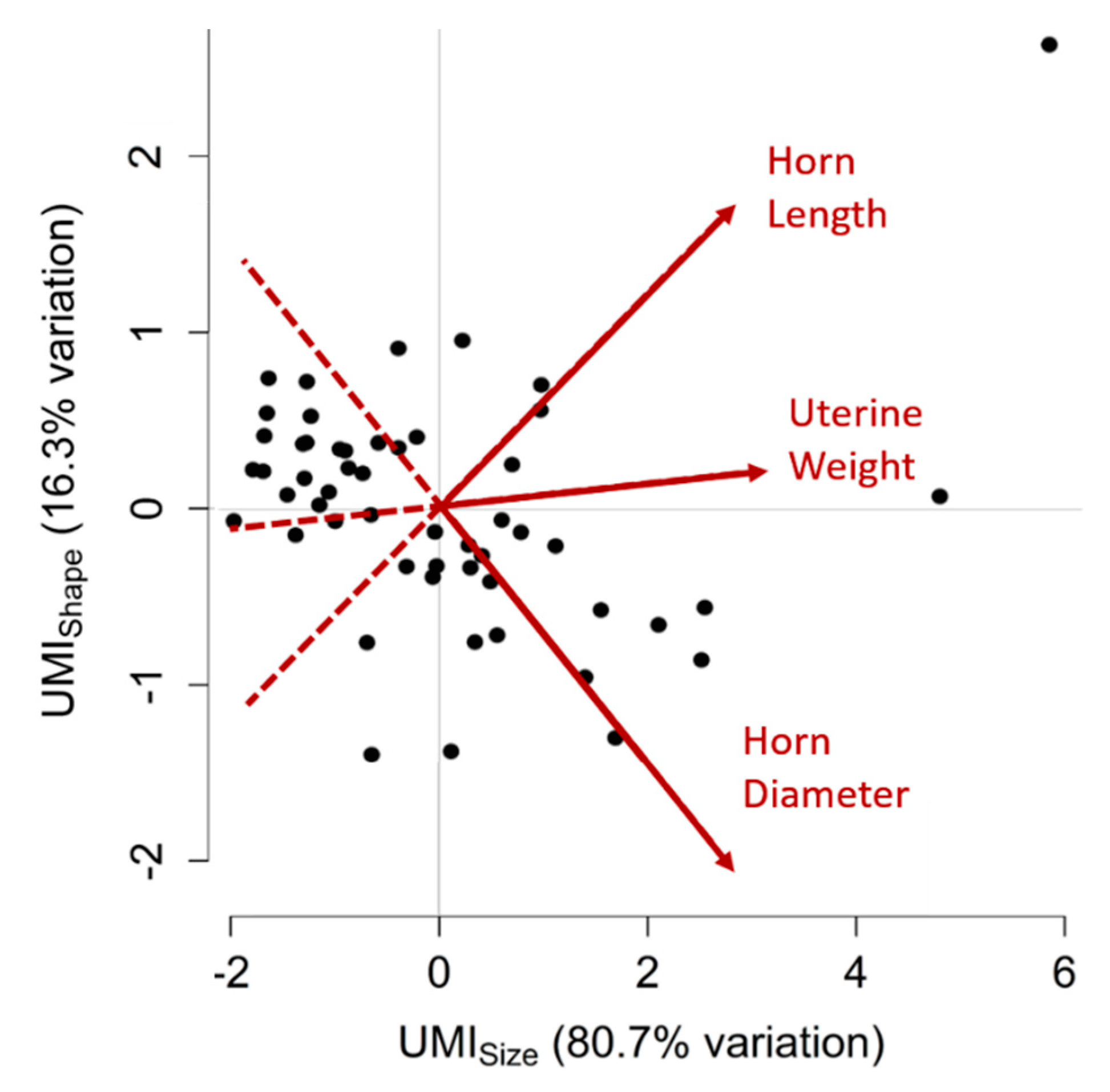
3.5. Uterine Mass Indices (UMIs)
3.6. Intra-Ovary Correlations
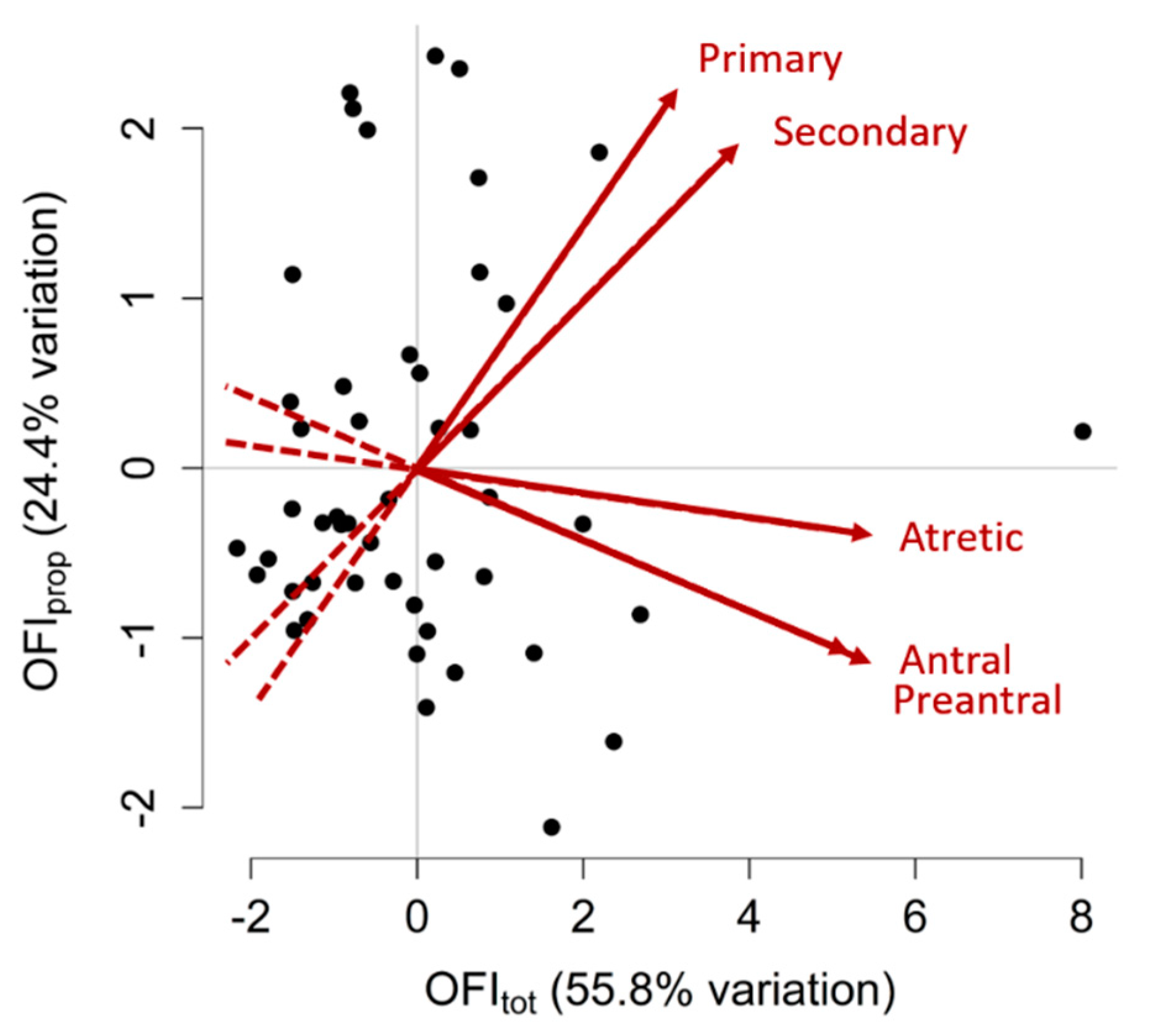
3.7. Ovarian Follicle Indices (OFIs)
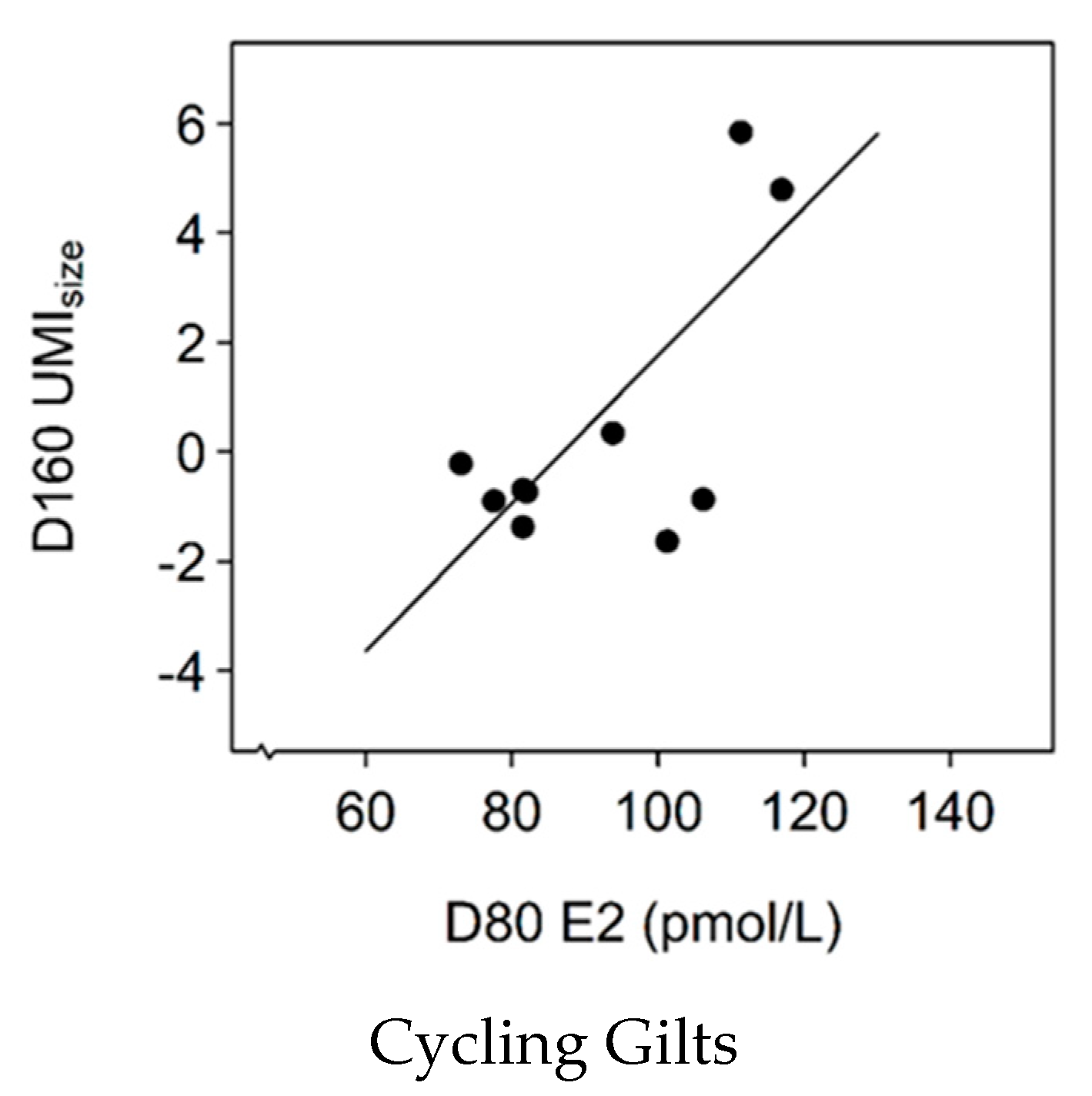
3.8. Association between E2 Levels and UMIsize
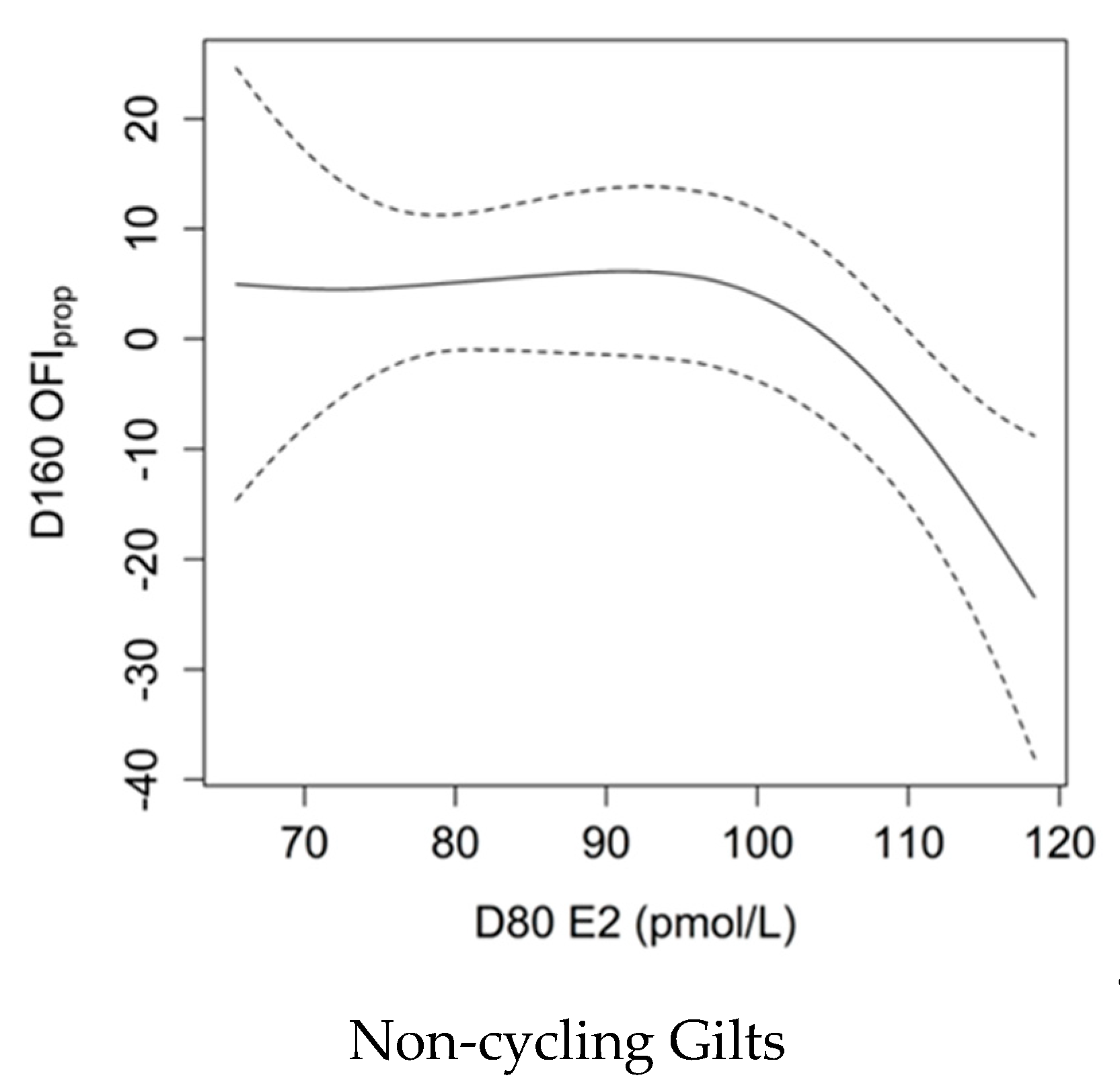
3.9. Association between E2 Levels and OFIprop
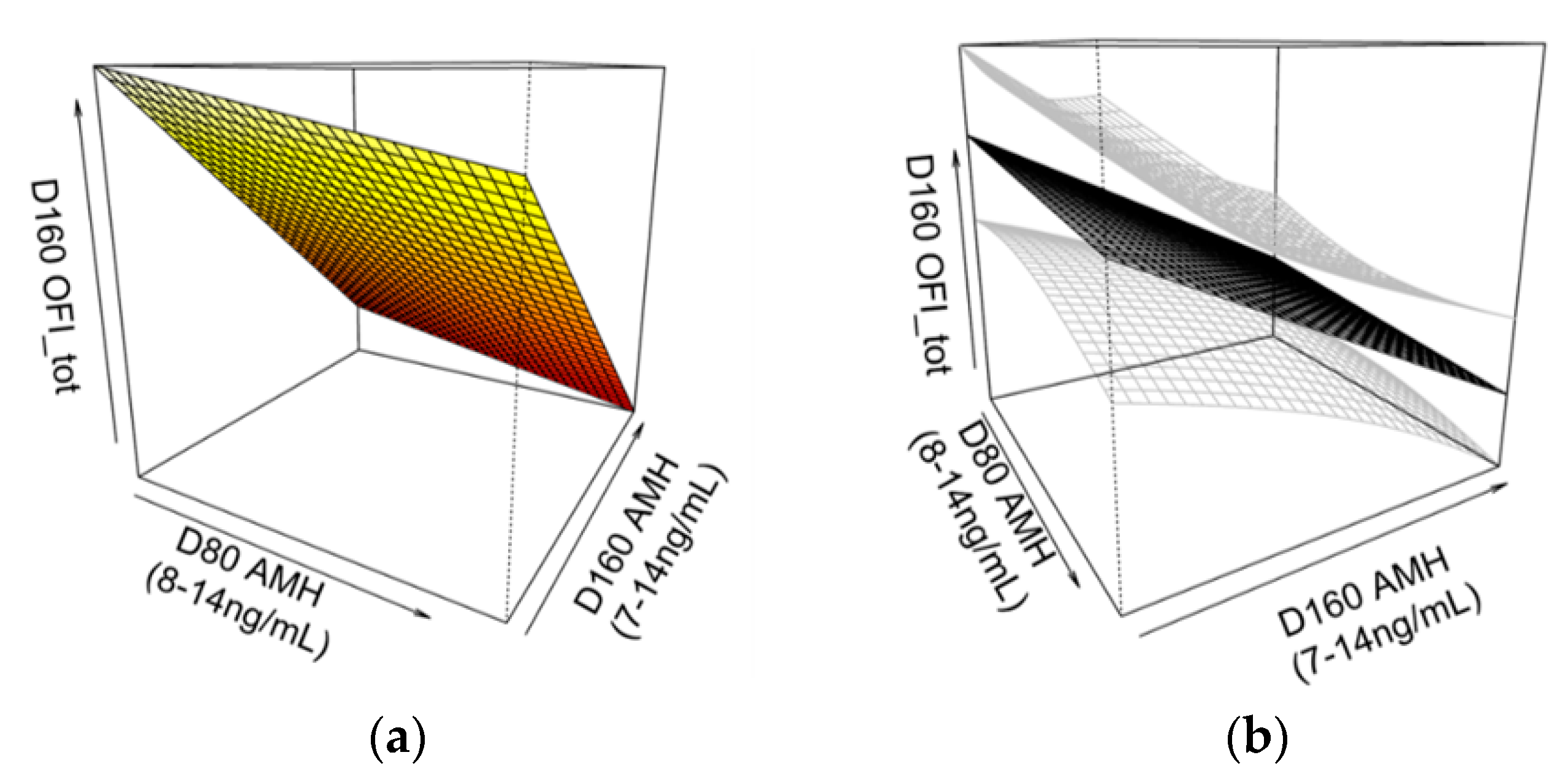
3.10. Association between AMH Levels and OFItot
3.11. Other Body Condition, Uterine and Ovarian Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hughes, P.E.; Smits, R.J.; Xie, Y.; Kirkwood, R.N. Relationships among gilt and sow live weight, P2 backfat depth, and culling rates. J. Swine Health Prod. 2010, 18, 301–305. [Google Scholar]
- Serenius, T.; Stalder, K.J.; Puonti, M. Impact of dominance effects on sow longevity. J. Anim. Breed. Genet. 2006, 123, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Plush, K.; Langendijk, P.; Kirkwood, R.N.; Pope, G.; Willis, S.; Riley, J. Key differences underlying top and bottom reproductive perfromers: Analysis of management programme data. Australian Pork Limited: 2016. Available online: https://australianpork.infoservices.com.au/items/2013-022 (accessed on 11 October 2019).
- Stalder, K.J.; Knauer, M.; Baas, T.J.; Rothschild, M.F.; Mabry, J.W. Sow Longevity. Pig News Inf. 2004, 25, 53N–74N. [Google Scholar]
- D’Allaire, S.; Drolet, R. Culling and mortality in breeding animals. In Diseases of Swine, 8th ed.; Straw, B.E., D’Allaire, S., Mengeling, W.L., Taylor, D.J., Eds.; Iowa State University Press: Ames, IW, USA, 1999; pp. 1003–1016. [Google Scholar]
- Kroes, Y.; Van Male, J.P. Reproductive lifetime of sows in relation to economy of production. Livest. Prod. Sci. 1979, 6, 179–183. [Google Scholar] [CrossRef]
- Monniaux, D.; Clement, F.; Dalbies-Tran, R.; Estienne, A.; Fabre, S.; Mansanet, C.; Monget, P. The ovarian reserve of primordial follicles and the dynamic reserve of antral growing follicles: What is the link? Biol. Reprod. 2014, 90, 85. [Google Scholar] [CrossRef]
- Warren, L.; Murawski, M.; Wilk, K.; Zieba, D.A.; Bartlewski, P.M. Suitability of antral follicle counts and computer-assisted analysis of ultrasonographic and magnetic resonance images for estimating follicular reserve in porcine, ovine and bovine ovaries ex situ. Exp. Biol. Med. 2015, 240, 576–584. [Google Scholar] [CrossRef]
- Ireland, J.L.H.; Scheetz, D.; Jimenez-Krassel, F.; Themmen, A.P.N.; Ward, F.; Lonergan, P.; Smith, G.W.; Perez, G.I.; Evans, A.C.O.; Ireland, J.J. Antral follicle count reliably predicts number of morphologically healthy oocytes and follicles in ovaries of young adult cattle. Biol. Reprod. 2008, 79, 1219–1225. [Google Scholar] [CrossRef]
- La Marca, A.; Sighinolfi, G.; Radi, D.; Argento, C.; Baraldi, E.; Artenisio, A.C.; Stabile, G.; Volpe, A. Anti-Müllerian hormone (AMH) as a predictive marker in assisted reproductive technology (ART). Hum. Reprod. Update 2010, 16, 113–130. [Google Scholar] [CrossRef]
- Hirayama, H.; Kageyama, S.; Naito, A.; Fukuda, S.; Fujii, T.; Minamihashi, A. Prediction of superovulatory response in Japanese Black cattle using ultrasound, plasma anti-Müllerian hormone concentrations and polymorphism in the ionotropic glutamate receptor AMPA1/GRIA1. J. Reprod. Dev. 2012, 58, 380–383. [Google Scholar] [CrossRef]
- Batista, E.O.S.; Macedo, G.G.; Sala, R.V.; Ortolan, M.D.D.V.; Sa Filho, M.F.; Del Valle, T.A.; Jesus, E.F.; Lopes, R.N.V.R.; Renno, F.P.; Baruselli, P.S. Plasma anti-Müllerian hormone as a predictor of ovarian antral follicular population in Bos indicus (Nelore) and Bos taurus (Holstein) heifers. Reprod. Domest. Anim. 2014, 49, 448–452. [Google Scholar] [CrossRef]
- Torres-Rovira, L.; Gonzalez-Bulnes, A.; Succu, S.; Spezzigu, A.; Manca, M.E.; Leoni, G.G.; Sanna, M.; Pirino, S.; Gallus, M.; Naitana, S.; et al. Predictive value of antral follicle count and anti-Müllerian hormone for follicle and oocyte developmental competence during the early prepubertal period in a sheep model. Reprod. Fertil. Dev. 2014, 26, 1094–1106. [Google Scholar] [CrossRef] [PubMed]
- Claes, A.; Ball, B.A.; Scoggin, K.E.; Esteller-Vico, A.; Kalmar, J.J.; Conley, A.J.; Squires, E.L.; Troedsson, M.H. The interrelationship between anti-Müllerian hormone, ovarian follicular populations and age in mares. Equine Vet. J. 2015, 47, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Monniaux, D.; Baril, G.; Laine, A.-L.; Jarrier, P.; Poulin, N.; Cognié, J.; Fabre, S. Anti-Müllerian hormone as a predictive endocrine marker for embryo production in the goat. Reproduction 2011, 142, 845–854. [Google Scholar] [CrossRef]
- Kevenaar, M.E.; Meerasahib, M.F.; Kramer, P.; van de Lang-Born, B.M.; de Jong, F.H.; Groome, N.P.; Themmen, A.P.; Visser, J.A. Serum anti-Müllerian hormone levels reflect the size of the primordial follicle pool in mice. Endocrinology 2006, 147, 3228–3234. [Google Scholar] [CrossRef]
- Almeida, F.R.C.L.; Costermans, N.G.J.; Soede, N.M.; Bunschoten, A.; Keijer, J.; Kemp, B.; Teerds, K.J. Presence of anti-Müllerian hormone (AMH) during follicular development in the porcine ovary. PLoS ONE 2018, 13, e0197894. [Google Scholar] [CrossRef]
- Steel, A.; Athorn, R.Z.; Grupen, C.G. Anti-Müllerian hormone and oestradiol as markers of future reproductive success in juvenile gilts. Anim. Reprod. Sci. 2018, 195, 197–206. [Google Scholar] [CrossRef]
- Schwarz, T.; Kopyra, M.; Nowicki, J. Physiological mechanisms of ovarian follicular growth in pigs--a review. Acta Vet. Hung. 2008, 56, 369–378. [Google Scholar] [CrossRef]
- Bidlingmaier, F.; Butenandt, O.; Knorr, D. Plasma gonadotropins and estrogens in girls with idiopathic precocious puberty. Pediatr. Res. 1977, 11, 91–94. [Google Scholar] [CrossRef]
- Nelson, A.H.; Mabry, J.W.; Benyshek, L.L.; Marks, M.A. Correlated response in reproduction, growth and composition to selection in gilts for extremes in age at puberty and backfat. Livest. Prod. Sci. 1990, 24, 237–247. [Google Scholar] [CrossRef]
- Patterson, J.L.; Beltranena, E.; Foxcroft, G.R. The effect of gilt age at first estrus and breeding on third estrus on sow body weight changes and long-term reproductive performance. J. Anim. Sci. 2010, 88, 2500–2513. [Google Scholar] [CrossRef]
- Young, L.D.; Leymaster, K.A.; Christenson, R.K. Opportunities for indirect selection for uterine capacity in the swine. J. Anim. Sci. Suppl. 1996, 74, 199. [Google Scholar]
- Lents, C.A.; Cushman, R.A.; Freking, B.A. Measures of the ovaries and uterus during development of gilts selected for differences in uterine capacity. J. Anim. Sci. 2014, 92, 2433–2439. [Google Scholar] [CrossRef] [PubMed]
- Foxcroft, G.R.; Dixon, W.T.; Dyck, M.K.; Novak, S.; Harding, J.C.S.; Almeida, F.C.R.L. Prenatal programming of postnatal development in the pig. Soc. Reprod. Fertil. Suppl. 2009, 66, 213–231. [Google Scholar] [PubMed]
- Schwarz, T.; Murawski, M.; Wierzchoś, E.; Bartlewski, P. An ultrasonographic study of ovarian antral follicular dynamics in prepubertal gilts during the expected activation of the hypothalamo-pituitary-ovarian axis. J. Reprod. Dev. 2013, 59, 409–414. [Google Scholar] [CrossRef]
- Knox, R.V. Physiology and endocrinology symposium: Factors influencing follicle development in gilts and sows and management strategies used to regulate growth for control of estrus and ovulation1. J. Anim. Sci. 2019, 97, 1433–1445. [Google Scholar] [CrossRef]
- Di Clemente, N.; Goxe, B.; Remy, J.J.; Cate, R.; Josso, N.; Vigier, B.; Salesse, R. Inhibitory effect of AMH upon the expression of aromatase and LH receptors by cultured granulosa-cells of rat and porcine immature ovaries. Endocrine 1994, 2, 553–558. [Google Scholar]
- Garrel, G.; Racine, C.; L’Hôte, D.; Denoyelle, C.; Guigon, C.J.; di Clemente, N.; Cohen-Tannoudji, J. Anti-Müllerian hormone: A new actor of sexual dimorphism in pituitary gonadotrope activity before puberty. Sci. Rep. 2016, 6, 23790. [Google Scholar] [CrossRef]
- Lashen, H.; Dunger, D.B.; Ness, A.; Ong, K.K. Peripubertal changes in circulating anti-Müllerian hormone levels in girls. Fertil. Steril. 2013, 99, 2071–2075. [Google Scholar] [CrossRef]
- Dufour, J.J.; Grasso, F.; Daviault, E.; Matton, P. Relationship between two morphological types of ovaries and their follicular microscopic population before puberty and their ensuing follicular development and ovulation rates at puberty in gilts. J. Anim. Sci. 1988, 66, 932–938. [Google Scholar] [CrossRef]
- Lahoz, B.; Alabart, J.L.; Cocero, M.J.; Monniaux, D.; Echegoyen, E.; Sanchez, P.; Folch, J. Anti-Müllerian hormone concentration in sheep and its dependence of age and independence of BMP15 genotype: An endocrine predictor to select the best donors for embryo biotechnologies. Theriogenology 2014, 81, 347–357. [Google Scholar] [CrossRef]
- Tummaruk, P.; Kesdangsakonwut, S. Uterine size in replacement gilts associated with age, body weight, growth rate, and reproductive status. Czech J. Anim. Sci. 2014, 59, 511–518. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (days) | Median | Mean | SD | Range | |
|---|---|---|---|---|---|
| Serum hormone levels: | |||||
| AMH (ng/mL) | 80 | 11.1 | 10.9 | 1.3 | (7.9–13.9) |
| AMH (ng/mL) | 160 | 12.0 | 11.8 | 1.2 | (8.9–14.0) |
| E2 (pmol/L) | 80 | 94.7 | 94.7 | 15.7 | (65.5–131.0) |
| E2 (pmol/L) | 160 | 104.2 | 106.2 | 14.7 | (72.6–143.1) |
| Carcass traits: | |||||
| CW (kg) | 160 | 78.0 | 78.1 | 10.0 | (58.1–112.4) |
| P2 fat score (mm) | 160 | 13.2 | 13.4 | 2.9 | (8.4–24.0) |
| Uterine traits: | |||||
| Weight (g) | 160 | 77.6 | 102.0 | 70.9 | (26.7–370.9) |
| Diameter (mm) | 160 | 12.3 | 12.3 | 2.3 | (8.6–17.2) |
| Length (mm) | 160 | 496.3 | 533.4 | 176.5 | (252.5–1450.0) |
| Ovarian follicle counts: | |||||
| Primary | 160 | 49,440 | 51,543 | 24,376 | (14,000–120,000) |
| Secondary | 160 | 31,360 | 37,374 | 21,636 | (9600–104,640) |
| Preantral | 160 | 3840 | 5502 | 5479 | (0–29,964) |
| Antral | 160 | 5600 | 7004 | 5430 | (640–29,091) |
| Atretic | 160 | 22,880 | 28,756 | 21,970 | (1920–107,636) |
| Total (Healthy) | 160 | 92,840 | 101,423 | 44,126 | (42,880–275,814) |
| Total (Healthy + Atretic) | 160 | 112,427 | 130,179 | 58,613 | (52,160–383,451) |
| D80 | D160 | |||
|---|---|---|---|---|
| AMH | E2 | AMH | E2 | |
| Uterine Weight | −0.25 ^ | 0.23 | 0.02 | −0.08 |
| Horn Diameter | −0.23 | 0.04 | 0.10 | 0.02 |
| Horn Length | −0.25 ^ | 0.21 | 0.17 | −0.09 |
| Primary | 0.02 | 0.02 | 0.21 | 0.00 |
| Secondary | 0.04 | −0.18 | 0.23 | −0.03 |
| Preantral | 0.00 | −0.24 ^ | −0.07 | 0.37 * |
| Antral | 0.02 | −0.13 | 0.08 | 0.26 ^ |
| Atretic | −0.10 | −0.16 | 0.18 | 0.11 |
| D80 | D160 | ||||
|---|---|---|---|---|---|
| AMH | E2 | AMH | E2 | ||
| D80 | AMH | ||||
| E2 | 0.23 ^ | ||||
| D160 | AMH | 0.13 | −0.05 | ||
| E2 | −0.07 | −0.32 * | −0.29 * | ||
| Uterine Weight | Horn diameter | Horn Length | |
|---|---|---|---|
| Uterine Weight | |||
| Horn diameter | 0.76* | ||
| Horn Length | 0.84 * | 0.52 * |
| Primary | Secondary | Preantral | Antral | Atretic | |
|---|---|---|---|---|---|
| Primary | |||||
| Secondary | 0.52 * | ||||
| Preantral | 0.17 | 0.28 ^ | |||
| Antral | 0.22 | 0.23 | 0.84 * | ||
| Atretic | 0.22 | 0.48 * | 0.61 * | 0.72 * |
| Follicle Type | Loading | |
|---|---|---|
| OFItot | OFIprop | |
| Primary | 0.28 | 0.66 |
| Secondary | 0.36 | 0.57 |
| Preantral | 0.50 | −0.33 |
| Antral | 0.52 | −0.34 |
| Atretic | 0.51 | −0.10 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steel, A.; Athorn, R.Z.; Grupen, C.G. Serum Concentrations of AMH and E2 and Ovarian and Uterine Traits in Gilts. Animals 2019, 9, 811. https://doi.org/10.3390/ani9100811
Steel A, Athorn RZ, Grupen CG. Serum Concentrations of AMH and E2 and Ovarian and Uterine Traits in Gilts. Animals. 2019; 9(10):811. https://doi.org/10.3390/ani9100811
Chicago/Turabian StyleSteel, Alicia, Rebecca Z. Athorn, and Christopher G. Grupen. 2019. "Serum Concentrations of AMH and E2 and Ovarian and Uterine Traits in Gilts" Animals 9, no. 10: 811. https://doi.org/10.3390/ani9100811
APA StyleSteel, A., Athorn, R. Z., & Grupen, C. G. (2019). Serum Concentrations of AMH and E2 and Ovarian and Uterine Traits in Gilts. Animals, 9(10), 811. https://doi.org/10.3390/ani9100811