4.1. Characteristics of Runs of Homozygosity
The characteristics of the ROH detected in this study corresponds well to the results obtained by other authors. Purfield et al. [
1] identified ROH for various breeds of cattle, including European breeds: Holstein-Friesian, Limousin, Simmental, Hereford and Charolais. The authors showed that the average sum of ROH length for segments longer than 5 Mb and identified using BovineHD BeadChip microarrays was the highest for the Hereford (145 Mb) and Holstein-Friesian (115 Mb) breeds. However, the Limousin, Simmental and Charolais breeds were characterized by much lower values, respectively 45 Mb, 55 Mb and 50 Mb. In the case of all ROH (>1 Mb), the average sum of the ROH lengths was respectively higher: Hereford—245 Mb; Holstein-Frisian—195 Mb; Limousin—68 Mb; Simmental—85 Mb; and Charolais—85 Mb. Similar results were also obtained by Iacolina et al. [
23], who compared European cattle breeds to the European bison using Illumina BovineHD BeadChip microarrays. The authors observed the highest average ROH lengths for Angus and Hereford breeds for the 1–5 Mb category (approximately 200 Mb). In the case of the Holstein-Friesian, Charolais, Simmental and Limousin breeds, these values were lower—in the range of approximately 100 Mb. In turn, in the research of Peripolli et al. [
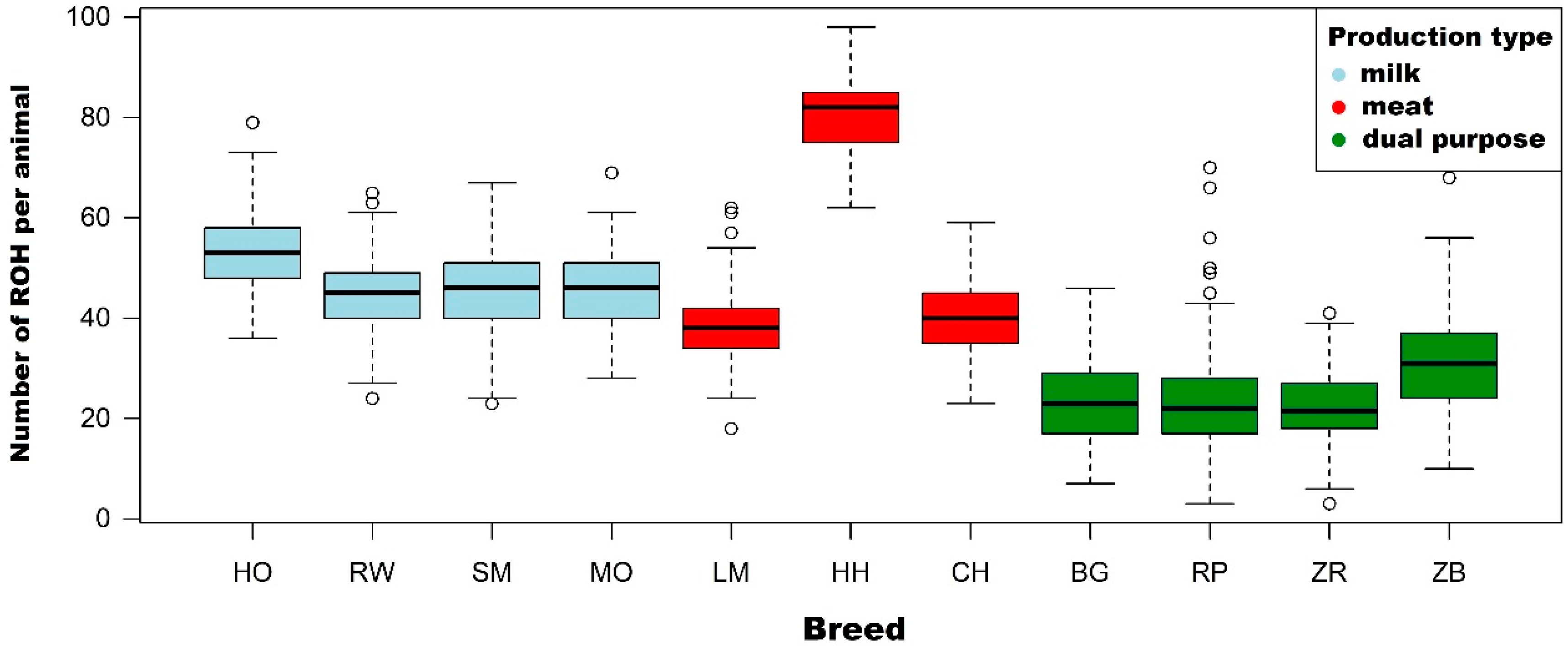
24] performed on Gyr cattle (Bos indicus), it was shown that the average sum of ROH lengths was in the range of 100 Mb per animal and the longest region was 108.9 Mb. These results are comparable with the results described in the present study for ROH with lengths >1 Mb and >4 Mb. In this study, the highest average ROH length was observed for the Hereford breed (378 Mb for ROH above 1 Mb and 253.4 Mb above 4 Mb), followed by the Holstein-Friesian Black-and-White variety (295.1 and 220.8 MB), and later, respectively, the following breeds: Charolais (163.2 Mb and 97.2 Mb), Simmental (169.1 and 87.4 Mb) and Limousin (147.4 Mb for ROH above 1 Mb and 80.3 Mb above 4 Mb). Apart from analyzing the ROH of high-production cattle breeds kept in Poland (HO, RW, SM, LM, MO, CH and HH), which are largely genetically close to their European counterparts, Polish indigenous breeds of cattle were also examined (BG, RP, ZR and ZB). These breeds are included in the genetic resources conservation programs. An analysis of ROH showed notable differences in the length and amount of ROH between native breeds and production breeds. Regarding the average number of ROH per animal for native breeds, it was shown to be in the range from 21 to 30, while for production breeds, it was in the range from 39 to 81. A similar trend can be observed for the average sum of ROH lengths—in the case of native breeds, it was lower and ranged from 105 to 135 Mb, while in the case of production breeds, it was in the range from 147 to 378 Mb.
According to the results obtained, we hypothesized that much higher mean length values and ROH numbers obtained for highly selected breeds in comparison to native breeds may be associated with strong directional selection and widespread and intensive use of artificial insemination. It has been proven that the use of biotechnics and intensive artificial selection significantly reduce genetic variability and increase the degree of relationship between animals [
25]. In addition, native breeds were characterized by low, compared to commercial breeds, average amounts of ROH with a length of more than 8 Mb (2.2–3.8 in the case of conserved breeds and 3.1–10.1 in production breeds). Due to the fact that the occurrence of long segments of ROH is related to the recent relationship within the parental population [
26], small amounts of long ROH segments indicate a low degree of the close relationship between animals from conserved herds. Similar differences between native breeds and production breeds were also observed by other authors [
1]. The implementation of the genetic resources conservation program itself (Program for the Conservation of Genetic Resources of Farm Animals), which focuses on minimizing inbreeding within protected populations, seems to be of great importance for the obtained results. Not without significance is the admission in the protected populations of a certain share of the blood of other breeds, which increases the genetic variation and diversity of segregating haplotypes.
Differences in the ROH statistics between highly selected and native breeds may also result from the intensity of selection used within these populations. Kim et al. [
27] studied Holstein cattle from populations with varying intensity of selection and observed that in animals undergoing intensive selection, the average length of ROH per animal was about 6.67 Mb, while for extensively selected animals, it was 6.26 Mb. In the case of an average amount of ROH per animal, the values were as follows: 40.4 for intensively selected animals and 31.1 for extensively selected animals. The same trend can be seen in the results of the present research by comparing intensively the selected production breeds and native breeds covered by the genetic resources conservation program.
4.2. Runs of Homozygosity as a Tool of Inbreeding Estimation
F
ROH (inbreeding coefficient calculated on the basis of ROH) is widely regarded as a reliable measure of individual autozygosity and provides information on the degree of inbreeding of individual herds of animals, taking into account both past and recent relatedness of individuals [
3,
28]. The literature data also indicated that F
ROH may be a better estimator of individual autozygosity than those based on pedigree data (F
PED) [
28]. However, it should be noted that the use of medium density SNP arrays, such as the one used in this research, leads to limited identification of short ROH that contribute significantly to ancient inbreeding [
17].
In many studies on cattle, a strong or moderate correlation between F
PED and F
ROH values was observed: 0.73 for ROH longer than 1 Mb and 0.70 for ROH longer than 10 Mb [
1] and from 0.485 to 0.715 for ROH longer than 1 Mb [
28]. In our previous study [
29] performed on Polish Holstein-Friesian cattle, slightly lower values of the correlation coefficients between F
ROH and F
PED were observed (in the range from 0.308 to 0.505 for ROH with a length of more than 1 Mb). This is mainly the result of using Spearman’s rank correlation coefficients, more appropriate for data sets with a large number of outliers and those not showing a normal distribution [
30], but often giving lower correlation values.
It is worth mentioning that the correlation between F
ROH and F
PED coefficients rises with the increase in the length of ROH segments used for calculations. According to Marras et al. [
26], this is due to the fact that ROH reflects both past and recent animal relatedness, while F
PED coefficients are based on pedigree records that may not contain information from many past generations. Therefore, F
ROH coefficient calculated on the basis of long ROH better reflects the recent relationship, and the F
ROH–F
PED correlation is usually higher considering only long ROH segments in the calculation. In studies conducted on pigs, Saura et al. [
31] showed that the mean value of F
ROH coefficient calculated for ROH with lengths >5 Mb was close to F
PED, while the mean for F
ROH calculated for ROH longer than 5 Mb was about four times lower than the average F
PED. Another study was presented by Scraggs et al. [
32], which suggested that F
PED does not determine true kinship within the Wagyu cattle population, as there were clear differences between F
ROH and F
PED coefficients. The authors showed much lower values of F
PED coefficients compared to F
ROH. These results are consistent with data obtained by other authors for cattle [
26,
33] and pigs [
31], in which the F
ROH coefficient was characterized by higher values than F
PED, suggesting that F
PED may underestimate the degree of inbreeding of the studied populations.
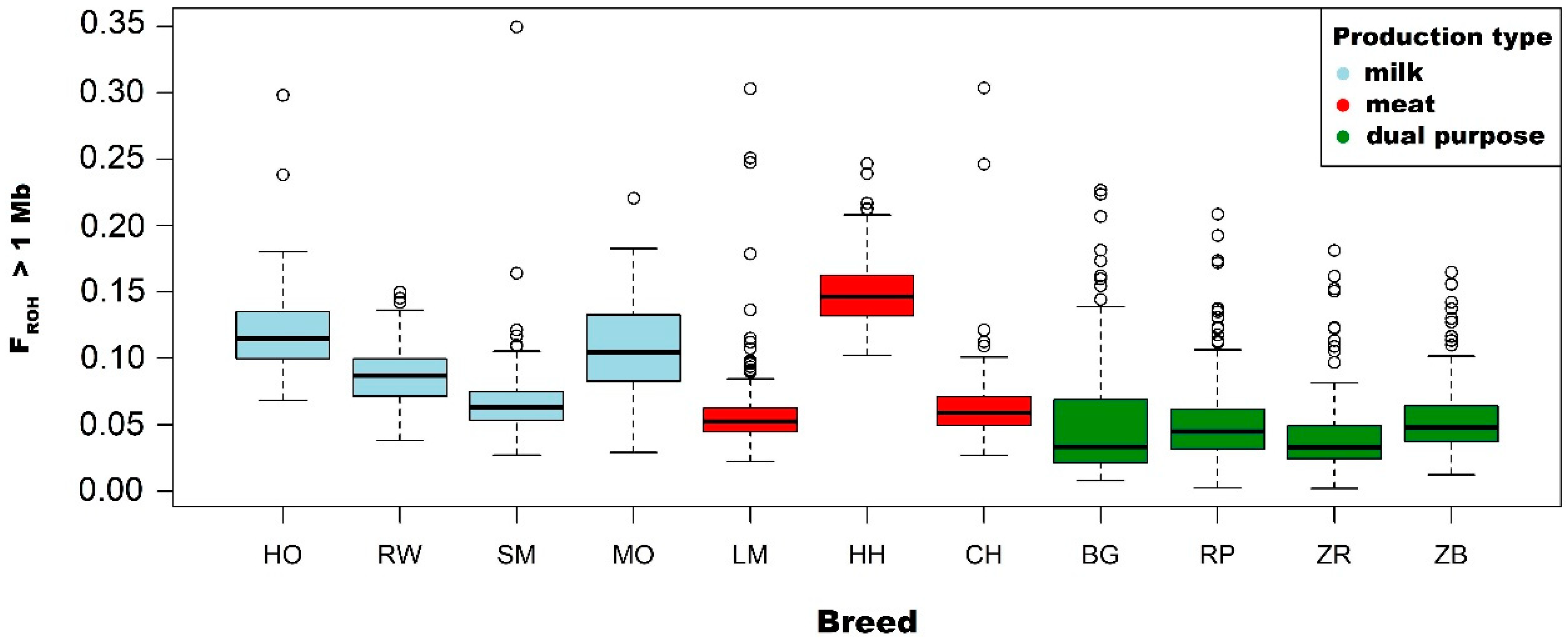
Due to the information presented above, in this research, thes FROH coefficient was used to assess the level of inbreeding of the selected cattle population maintained in Poland. The highest mean values of inbreeding coefficient calculated for ROH with a length above 1 Mb were observed in the Hereford (0.151), Holstein-Friesian Black-and-White variety (0.118) and Montbeliarde (0.108). The lowest FROH values, as expected, were noted for native breeds: White-Backed (0.051), Polish Red (0.053), Polish Red-and-White (0.042) and Polish Black-and-White (0.054). In addition, mean FROH coefficients calculated for ROH longer than 8 and 16 Mb were the highest in Montbeliarde, Holstein-Friesian Black-and-White and Hereford breeds, which suggests a relatively high degree of relatedness between animals and a reduction in genetic variation.
The inbreeding level estimated in this work for individual cattle breeds generally corresponds to the results obtained by other authors. In the study of Ferenčaković et al. [
28], F
ROH inbreeding coefficients for ROH with lengths above 1 Mb was calculated and high average autozygosity levels (0.151) were found for strongly selected Brown Swiss cattle population and significantly lower for native cattle (0.052 for Pinzgauer cattle and 0.066 for Tyrol Gray). In addition, in another study, Ferenčaković et al. [
20] observed varying F
ROH values, depending on the cattle breed: 0.156 for brown Swiss cattle; 0.088 for Fleckvieh cattle; 0.099 for Norwegian Red cattle; 0.087 for Tyrol Gray cattle and 0.09 for Simmental cattle. In the case of Holstein cattle maintained in the USA, F
ROH calculated for ROH with a length of more than 5 Mb was low and amounted to 0.038 [
34]. In Holstein cattle kept in Europe, higher F
ROH coefficients were found in the range from 0.081 for ROH > 1 Mb to 0.046 for ROH > 5 Mb [
1]. F
ROH values for the Polish Holstein-Friesian cattle described in this paper was slightly higher than that observed for other populations of Holstein-Friesian cattle in Europe; however, similar to the other high-production European dairy breeds [
1].
4.3. Analysis of Genomic Regions with a High Frequency of ROH Occurrence
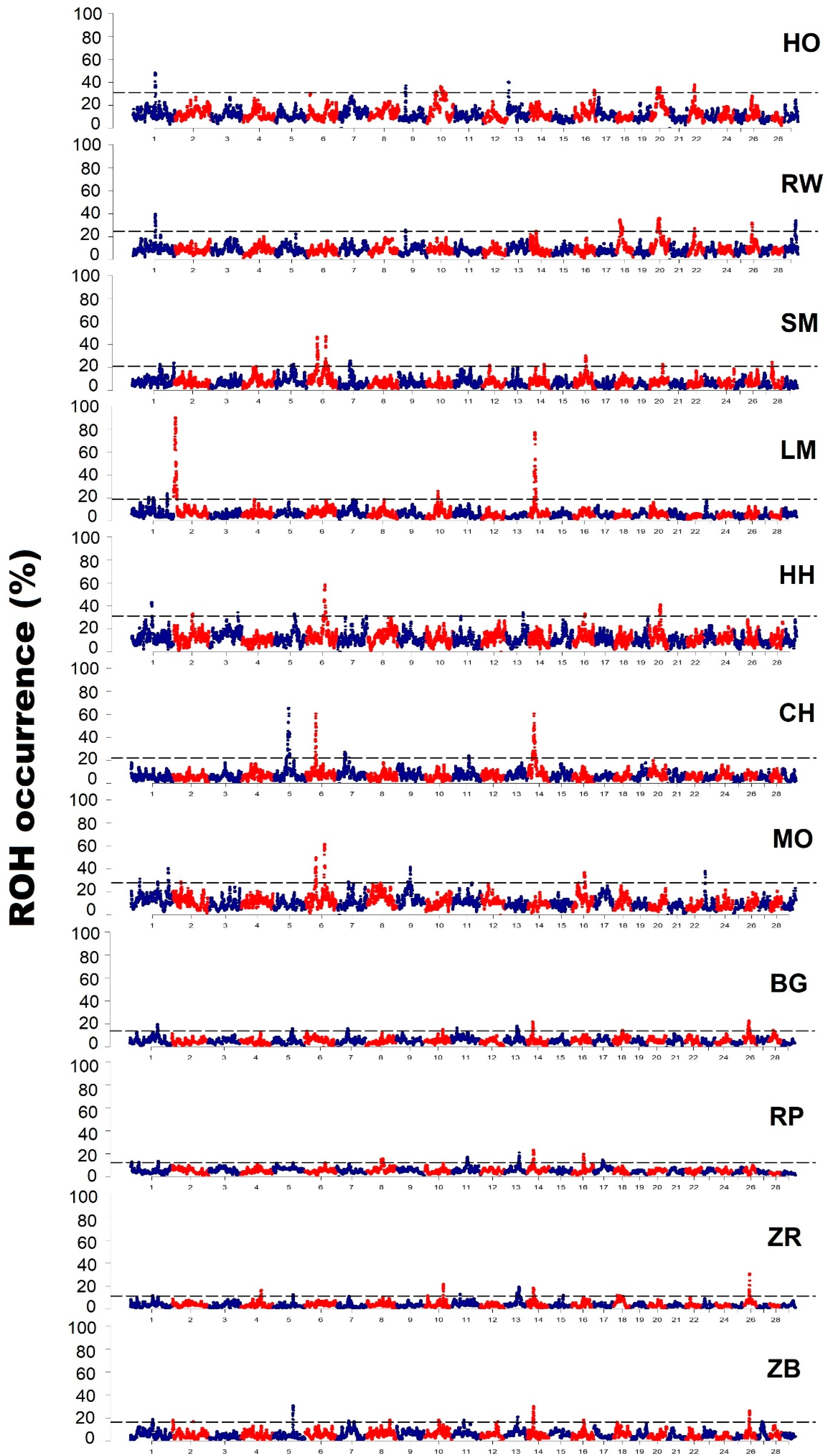
The examination of runs of homozygosity distribution across the genome shows that their dispersal in particular regions of the genome is unique and they can form characteristic patterns depending on a given population. These genomic regions with the highest frequency of ROH occurrence were called by Nothnagel et al. [
13] and Pemberton et al. [
14] with a term “ROH islands”. In studies of many authors, it is suggested that these regions are shaped by the influence of strong selection pressure on variants located at a given locus [
10,
11,
12,
14]. In addition, Zhang et al. [
25] confirmed a significant correlation between short regions with a high frequency of occurrence of ROH and genomic regions considered to be under the influence of selection using methods based on F
ST coefficient and iHS (integrated haplotype score).
An overview of the available literature indicates that many authors have observed and described the occurrence of ROH islands and their relationship with QTL for important production traits in cattle. Purfield et al. [
1] observed that the genomic regions located on BTA 7, BTA 14, BTA 16 and BTA 18 were characterized by a high frequency of ROH occurrence and included important genes related to immune traits, muscularity and ease of calving. In particular, BTA 5 and BTA 9 were characterized by an increased number of long ROH above 20 Mb, within which there were numerous QTL regions associated with the production of milk fat and growth characteristics of cattle [
1].
In this study, several genomic regions with extremely high levels of autozygosity and frequency of ROH were observed in individual breeds of cattle. An example of such a region may be the initial sequence of chromosome 2 in the Limousin breed, within which there is a myostatin gene locus (
MSTN), which is a strong QTL for muscle traits in this breed [
35]. A similar region on BTA 2 was found by Marras et al. [
26] by studying Piedmontese cattle, where almost 90% of all animals were characterized by the occurrence of ROH in the proximal part of the second chromosome. Other regions of interest, potentially influenced by the directional selection, may be two regions located on chromosome 5 and 6 in the Charolais breed, within which genes related to growth factors and coat color are located. One of these genes is
ERBB3—encoding the epidermal growth factor receptor. The literature showed that
ERRB3 controls the proliferation and myogenic diversity of muscle stem cells [
36]. Then, there are the
INHBC and
INHBE genes that are part of the TGFB signaling pathway—a transforming growth factor beta, which is an essential regulator of fibroblast proliferation, collagen synthesis in muscle tendons and in the case of muscular tissue can affect muscle atrophy [
37]. Therefore, these genes may potentially be associated with meat traits of Charolais cattle that are subject to selection in this breed. Moreover, in various studies [
38] within this ROH island, the
PMEL gene was proposed as the one associated with the white coat color of Charolais cattle. Another region with high homozygosity was observed in the proximal part of chromosome 14. This region was characterized by a high frequency of occurrence of ROH, in the range of 60% to 80% in the population of LM and CH breeds (respectively); however, elevated levels of homozygosity were also observed for this region in other breeds of cattle: RW, BG, RP, ZB and ZR. The identified region included the
DGAT1 gene locus, with known influence on fat percentage in milk and
FABP4 gene associated in numerous previous studies with transport, regulation and lipid metabolism [
39]. In addition, the
FABP4 gene was presented as affecting the qualitative characteristics of meat in cattle and also affecting fat content in meat [
40]. In turn, in the studies of Zhou et al. [
41], the effect of the
FABP4 gene on the amount of milk produced and protein content in milk was also observed.
When comparing the results of this research to literature data, a large similarity of the location of ROH islands identified in different breeds of cattle can be observed. In studies of Mészáros et al. [
42], a ROH island was observed in Tyrol Gray cattle located on chromosome 6 in the 36–41 Mb position. In the same chromosomal region, in this study, regions with an increased incidence of ROH were identified for the Montbeliarde, Simmental and Charolais breeds. The research performed on domestic cattle allowed the detection of four ROH islands, two of which were also identified in this study: the first on BTA 6 in the location of 38.2–39.4 Mb, which coincides with the ROH islands observed in the MO, SM and CH breeds; the second on BTA 16 with the location of 43.8–45.0 Mb, which coincides with the regions with high ROH frequency for the MO, SM, HH, RP and ZB breeds [
43]. In the studies of Marras et al. [
26], the authors observed characteristic ROH islands on chromosomes 2 and 6. The ROH island on chromosome 2 was characteristic to Piemontese cattle and was associated with the MSTN gene. A similar effect resulting from strong selection pressure on the
MSTN gene was observed in this research in Limousin cattle. In contrast, ROH island located on chromosome 6 in Holstein and Italian Brown dairy cattle included
ABCG2 and
FAM13A1 genes, which are known to be associated with milk traits [
44,
45]. The same genes were identified in regions with a high incidence of ROH in this study in the following cattle breeds: SM, CH, MO (
ABCG2 gene) and HO, RW, SM, CH, MO and ZB (
FAM13A1 gene).
In the present study, in regions with a high frequency of ROH occurrence, a number of other genes associated with cattle production traits were also observed. An example of such a gene identified in the ROH island on chromosome 2 is the
STAT1 gene observed in ZB cattle. Cobanoglu et al. [
46]’s study demonstrated the important role of this gene in the regulation process of transcription of other genes involved in the synthesis of milk proteins and fat metabolism. In dairy cattle breeds HO and RW, within ROH islands, the
GHR gene was observed, which has a significant impact on the quantity and composition of milk [
47,
48]. In addition, the same breeds have demonstrated the potential effect of selection pressure on the
TRH gene belonging to the thyrotropin secretion signaling pathway, which has a significant effect on growth hormone secretion and prolactin [
49]. Literature data confirm that exogenous stimulation of these hormones promotes a significant increase in milk yield [
50,
51]. Further examples of genes identified in ROH islands in the RW, BG, ZR and ZB breeds are SUFU and BTRC, that regulate the proliferation of lactate stem cells. In humans, the
SUFU gene acts as a negative regulator of the hedgehog signaling pathway, whereas the
BTRC gene controls the self-renewal process of the mammary gland stem cells [
52]. In BG, ZR and ZB breeds, in the ROH islands, we also detected the
CHUK gene, which, in the literature, is described as one of the genes responsible for the development of mammary gland and lactation [
53]. In addition, genes participating in the Wnt signaling pathway were identified in RW, SM, BG and RP cattle breeds. The Wnt signaling pathway is responsible for the regulation of cells proliferation, their morphology, apoptosis processes and differentiation as well as the development of the mammary gland [
53]. Another group of genes identified in the ROH islands are genes related to the signaling pathway of the angiogenesis process observed in the following breeds: HO, SM, MO, RP and HH, which may also be associated with the development of the mammary gland and affect its blood supply [
54]. In addition, in the case of conserved breeds, the highest number of genes identified in the ROH islands belonged to the metabolic pathways associated with the activation of B, T cells and inflammatory processes involving cytokine and the chemokine signaling pathway, which may be related to the higher resistance to diseases characteristic of these breeds.
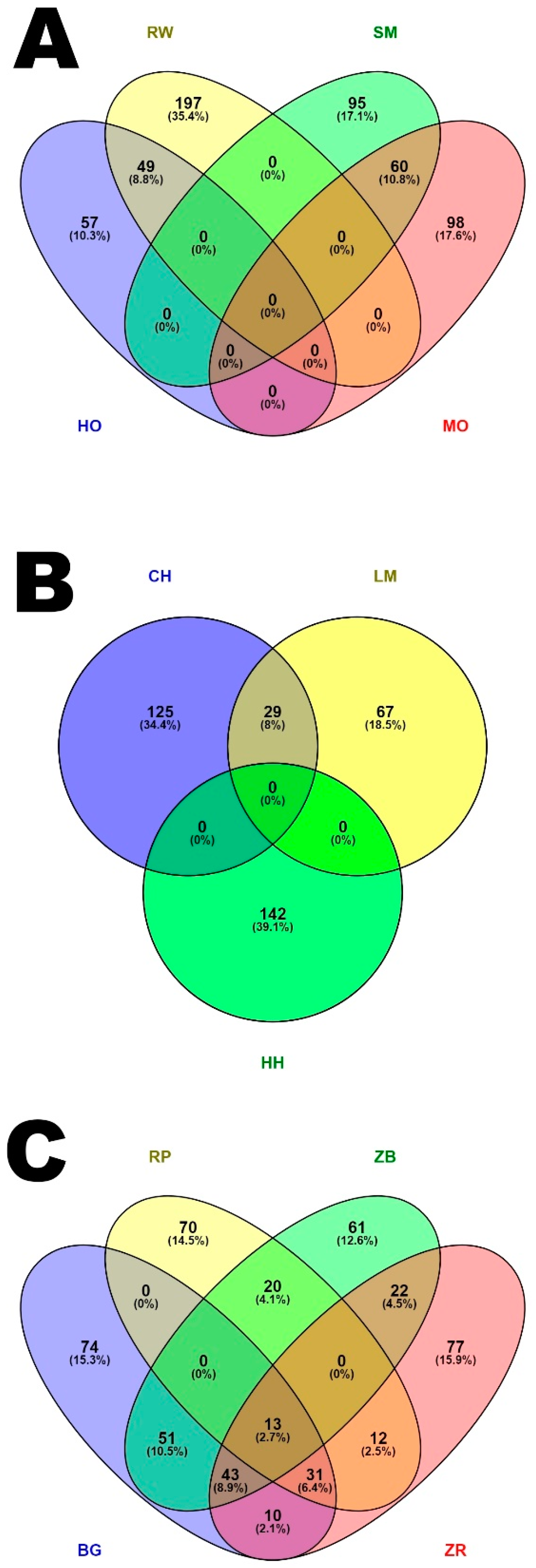
When analyzing genes occurring in ROH islands and observed in more than one breed, 13 genes were found between BG, RP, ZB and ZR cattle breeds. These genes have been described previously as affecting growth in cattle and humans (
TGS1,
LYN,
CHCHD7,
SDR16C5,
TMEM68) [
55,
56,
57,
58], ease of calving (
RPS20) [
59] and traits related to puberty (
FAM110B) [
60]. These traits are also subject of selection in cattle of different breeds and may have been fixed during a long-term breeding process.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}