An 11-bp Indel Polymorphism within the CSN1S1 Gene Is Associated with Milk Performance and Body Measurement Traits in Chinese Goats

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample and Data Collection
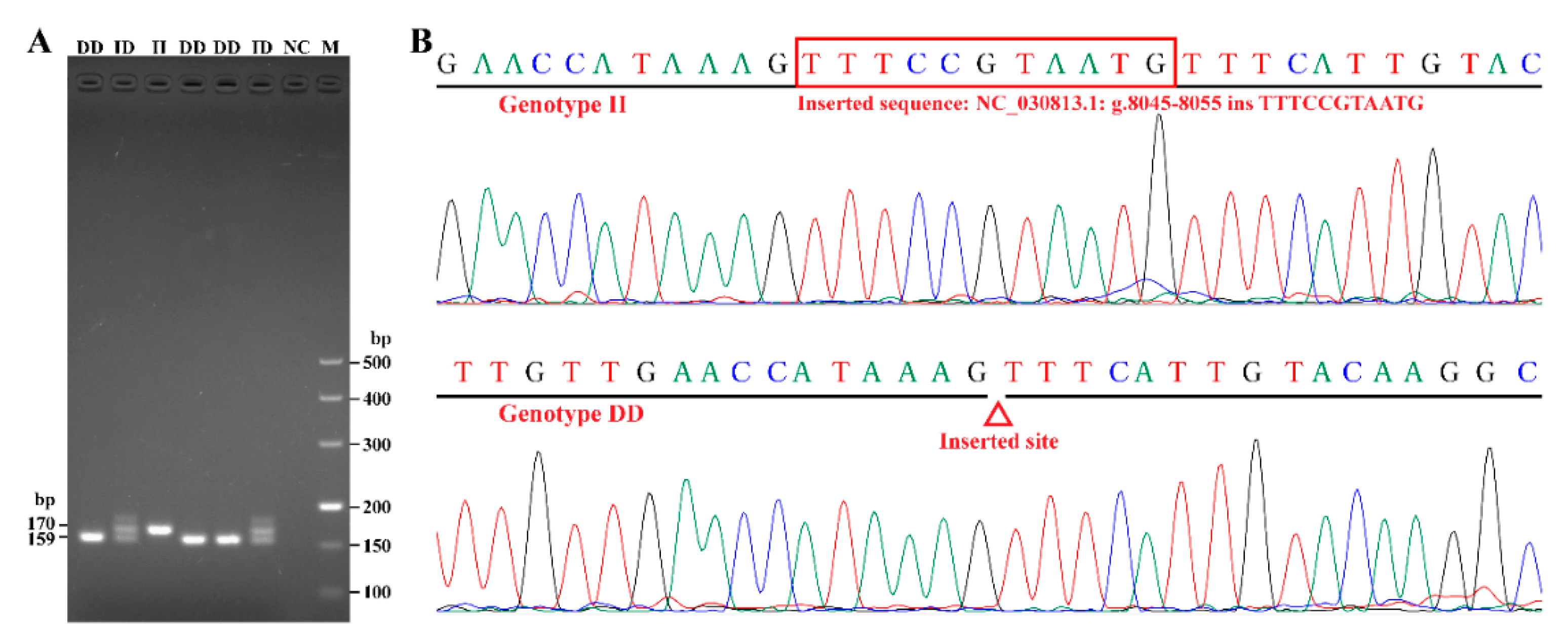
2.2. DNA Extraction and Indel Genotyping
2.3. Statistical Analysis
3. Results
3.1. Genetic Parameter of 11-bp Indel of CSN1S1 in Three Chinese Goat Breeds
3.2. Association between the 11-bp Indel and Milk Performance Traits in the Guanzhong Dairy Goat
3.3. Association Analysis of the 11-bp Indel with Body Measurement Traits in Three Goat Breeds
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, K.; Yan, H.; Xu, H.; Yang, Q.; Zhang, S.; Pan, C.; Chen, H.; Zhu, H.; Liu, J.; Qu, L.; et al. A novel indel within goat casein alpha S1 gene is significantly associated with litter size. Gene 2018, 671, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Chang, L.; Lan, X.; Asif, N.; Guan, F.; Fu, D.; Li, B.; Yan, C.; Zhang, H.; Zhang, X.; et al. Genome-wide definition of selective sweeps reveals molecular evidence of trait-driven domestication among elite goat (Capra species) breeds for the production of dairy, cashmere, and meat. Gigascience 2018, 7, giy105. [Google Scholar] [CrossRef] [PubMed]
- Peacock, C. Goats—A pathway out of poverty. Small. Rumin. Res. 2005, 60, 179–186. [Google Scholar] [CrossRef]
- Dubeuf, J.P.; Morand-Fehr, P.; Rubino, R. Situation, changes and future of goat industry around the world. Small. Rumin. Res. 2004, 51, 165–173. [Google Scholar] [CrossRef]
- FAO Commission on Genetic Resources for Food and Agriculture Assessments. The Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture; FAO: Rome, Italy, 2015; Available online: http://www.fao.org/publications/sowangr/en/ (accessed on 18 September 2019).
- Kang, Z.; Jiang, E.; Wang, K.; Pan, C.; Chen, H.; Yan, H.; Zhu, H.; Liu, J.; Qu, L.; Lan, X. Goat membrane associated ring-CH-type finger 1 (MARCH1) mRNA expression and association with litter size. Theriogenology 2019, 128, 8–16. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, S.; Li, J.; Wang, X.; Peng, K.; Lan, X.; Pan, C. Development of a touch-down multiplex PCR method for simultaneously rapidly detecting three novel insertion/deletions (indels) within one gene: An example for goat GHR gene. Anim. Biotechnol. 2019, 30, 366–371. [Google Scholar] [CrossRef]
- Jonas, E.; de Koning, D.J. Genomic selection needs to be carefully assessed to meet specific requirements in livestock breeding programs. Front. Genet. 2015, 6, 49. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Zhang, Y.; Bai, Y.; Yang, H.; Yan, H.; Liu, J.; Shi, L.; Song, X.; Li, L.; Dong, S.; et al. Relationship between SNPs of POU1F1 gene and litter size and growth traits in Shaanbei white cashmere goats. Animals 2019, 9, 114. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Wang, J.; Liu, N.; Cui, W.; Dong, W.; Xing, B.; Pan, C. Pig SOX9: Expression profiles of Sertoli cell (SCs) and a functional 18 bp indel affecting testis weight. Theriogenology 2019, 138, 94–101. [Google Scholar] [CrossRef]
- Kang, Z.; Zhang, S.; He, L.; Zhu, H.; Wang, Z.; Yan, H.; Huang, Y.; Dang, R.; Lei, C.; Chen, H.; et al. A 14-bp functional deletion within the CMTM2 gene is significantly associated with litter size in goat. Theriogenology 2019, 139, 49–57. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Q.; Zhang, S.; Zhang, X.; Pan, C.; Chen, H.; Zhu, H.; Lan, X. Genetic Effects of single nucleotide polymorphisms in the goat GDF9 gene on prolificacy: True or false positive? Animals 2019, 9, 886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, T.; Li, W.; Liu, D.; Liang, K.; Wang, X.; Li, H.; Jiang, R.; Tian, Y.; Kang, X.; Li, Z. Two insertion/deletion variants in the promoter region of the QPCTL gene are significantly associated with body weight and carcass traits in chickens. Anim. Genet. 2019, 50, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, R.; Wu, Z.; Hou, Y.; Bazer, F.W.; Wu, G. Amino acids and mammary gland development: Nutritional implications for milk production and neonatal growth. J. Anim. Sci. Biotechnol. 2016, 7, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juvarajah, T.; Wan-Ibrahim, W.I.; Ashrafzadeh, A.; Othman, S.; Hashim, O.H.; Fung, S.Y.; Abdul-Rahman, P.S. Human milk fat globule membrane contains hundreds of abundantly expressed and nutritionally beneficial proteins that are generally lacking in caprine milk. Breastfeed. Med. 2018, 13, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Grosclaude, F.; Martin, P. Casein polymorphism in the goat. In Milk Protein Polymorphism; International Dairy Federation, Ed.; Special Issue, 9702; International Dairy Federation: Brussels, Belgium, 1997; pp. 241–253. [Google Scholar]
- Martin, P.; Szymanowska, M.; Zwierzchowski, L.; Leroux, C. The impact of genetic polymorphisms on the protein composition of ruminant milks. Reprod. Nutr. Dev. 2002, 42, 433–459. [Google Scholar] [CrossRef]
- Guan, D.; Mármol-Sánchez, E.; Cardoso, T.F.; Such, X.; Landi, V.; Tawari, NR.; Amills, M. Genomic analysis of the origins of extant casein variation in goats. J. Dairy Sci. 2019, 102, 5230–5241. [Google Scholar] [CrossRef]
- Yue, X.P.; Zhang, X.M.; Wang, W.; Ma, R.N.; Deng, C.J.; Lan, X.Y.; Chen, H.; Li, F.; Xu, X.R.; Ma, Y.; et al. The CSN1S1 N and F alleles identified by PCR-SSCP and their associations with milk yield and composition in Chinese dairy goats. Mol. Biol. Rep. 2011, 38, 2821–2825. [Google Scholar] [CrossRef]
- Vacca, G.M.; Dettori, M.L.; Piras, G.; Manca, F.; Paschino, P.; Pazzola, M. Goat casein genotypes are associated with milk production traits in the Sarda breed. Anim. Genet. 2014, 45, 723–731. [Google Scholar] [CrossRef]
- Mestawet, T.A.; Girma, A.; Ådnøy, T.; Devold, T.G.; Vegarud, G.E. Newly identified mutations at the CSN1S1 gene in Ethiopian goats affect casein content and coagulation properties of their milk. J. Dairy Sci. 2013, 96, 4857–4869. [Google Scholar] [CrossRef]
- Carillier-Jacquin, C.; Larroque, H.; Robert-Granié, C. Including αs1 casein gene information in genomic evaluations of French dairy goats. Genet. Sel. Evol. 2016, 48, 54. [Google Scholar] [CrossRef] [Green Version]
- Cecchinato, A.; Chessa, S.; Ribeca, C.; Cipolat-Gotet, C.; Bobbo, T.; Casellas, J.; Bittante, G. Genetic variation and effects of candidate-gene polymorphisms on coagulation properties, curd firmness modeling and acidity in milk from Brown Swiss cows. Animal 2015, 9, 1104–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noce, A.; Pazzola, M.; Dettori, M.L.; Amills, M.; Castelló, A.; Cecchinato, A.; Bittante, G.; Vacca, G.M. Variations at regulatory regions of the milk protein genes are associated with milk traits and coagulation properties in the Sarda sheep. Anim. Genet. 2016, 47, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Bonfatti, V.; Giantin, M.; Gervaso, M.; Coletta, A.; Dacasto, M.; Carnier, P. Effect of CSN1S1-CSN3 (αS1-κ-casein) composite genotype on milk production traits and milk coagulation properties in Mediterranean water buffalo. J. Dairy Sci. 2012, 95, 3435–3443. [Google Scholar] [CrossRef] [PubMed]
- Masarwi, M.; Gabet, Y.; Dolkart, O.; Brosh, T.; Shamir, R.; Phillip, M.; Gat-Yablonski, G. Skeletal effect of casein and whey protein intake during catch-up growth in young male Sprague-Dawley rats. Br. J. Nutr. 2016, 116, 59–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Miao, J.; Sun, S.; Luo, Z.; Xia, Z.; Guo, B.; Liu, F.; Chen, Y.; Ying, D.; Miao, B.; et al. The effect of dietary magnesium and caseinphosphopeptides on bone metabolism in rats. Food. Funct. 2017, 8, 4487–4495. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Sun, S.; Guo, B.; Miao, B.; Luo, Z.; Xia, Z.; Ying, D.; Liu, F.; Guo, B.; Tang, J.; et al. Bioactive peptide isolated from casein phosphopeptides promotes calcium uptake in vitro and in vivo. Food. Funct. 2018, 9, 2251–2260. [Google Scholar] [CrossRef] [PubMed]
- Mulsant, P.; Lecerf, F.; Fabre, S.; Schibler, L.; Monget, P.; Lanneluc, I.; Pisselet, C.; Riquet, J.; Monniaux, D.; Callebaut, I.; et al. Mutation in bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola Mérino ewes. Proc. Natl. Acad. Sci. USA. 2001, 98, 5104–5109. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Hu, W.; Di, R.; Liu, Q.; Wang, X.; Zhang, X.; Zhang, J.; Chu, M. Expression analysis of the prolific candidate genes, BMPR1B, BMP15, and GDF9 in Small Tail Han ewes with three fecundity (FecB gene) genotypes. Animals 2018, 8, 166. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, G.W.; Lord, E.A.; Penty, J.M.; Dodds, K.G.; Broad, T.E.; Cambridge, L.; Sunden, S.L.; Stone, R.T.; Crawford, A.M. The Booroola fecundity (FecB) gene maps to sheep chromosome 6. Genomics 1994, 22, 148–153. [Google Scholar] [CrossRef]
- Lan, X.Y.; Chen, H.; Pan, C.Y.; Li, R.B.; Li, X.C.; Fang, X.T. Relationship between polymorphisms of CSN3, CSN1S2 and β-lg genes and litter sizes of Xinong Saanen dairy goat. Sci. Agric. Sin. 2005, 11, 2333–2338. [Google Scholar]
- Lan, X.Y.; Zhao, H.Y.; Li, Z.J.; Zhou, R.; Pan, C.Y.; Lei, C.Z.; Chen, H. Exploring the novel genetic variant of PITX1 gene and its effect on milk performance in dairy goats. J. Integr. Agric. 2013, 12, 118–126. [Google Scholar] [CrossRef]
- Wang, X.Y.; Yang, Q.; Wang, K.; Zhang, S.H.; Pan, C.Y.; Chen, H.; Qu, L.; Yan, H.L.; Lan, X.Y. A novel 12-bp indel polymorphism within the GDF9 gene is significantly associated with litter size and growth traits in goats. Anim. Genet. 2017, 48, 735–736. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Jiang, E.; Wang, K.; Zhang, Y.; Yan, H.; Qu, L.; Chen, H.; Lan, X.; Pan, C. Two insertion/deletion variants within SPAG17 gene are associated with goat body measurement traits. Animals 2019, 9, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic. Acids. Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Li, Z.Q.; Zhang, Z.; He, Z.D.; Tang, W.; Li, T.; Zeng, Z.; He, L.; Shi, YY. A partition-ligation-combination-subdivision EM algorithm for haplotype inference with multiallelic markers: Update of the SHEsis (http://analysis.bio-x.cn). Cell. Res. 2009, 19, 519–523. [Google Scholar] [CrossRef]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA. 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yang, Q.; Wang, K.; Yan, H.; Pan, C.; Chen, H.; Liu, J.; Zhu, H.; Qu, L.; Lan, X. Two strongly linked single nucleotide polymorphisms (Q320P and V397I) in GDF9 gene are associated with litter size in cashmere goats. Theriogenology 2019, 125, 115–121. [Google Scholar] [CrossRef]
- Lan, X.Y.; Pan, C.Y.; Chen, H.; Zhang, C.L.; Li, J.Y.; Zhao, M.; Lei, C.Z.; Zhang, A.L.; Zhang, L. An AluI PCR-RFLP detecting a silent allele at the goat POU1F1 locus and its association with production traits. Small. Rumin. Res. 2007, 73, 8–12. [Google Scholar] [CrossRef]
- Ren, F.; Yu, S.; Chen, R.; Lv, X.; Pan, C. Identification of a novel 12-bp insertion/deletion (indel) of iPS-related Oct4 gene and its association with reproductive traits in male piglets. Anim. Reprod. Sci. 2017, 178, 55–60. [Google Scholar] [CrossRef]
- Nagamine, C.M.; Chan, K.; Lau, Y.F. A PCR antifact generation of heteroduplexes. Am. J. Hum. Genet. 1989, 45, 337–339. [Google Scholar]
- Zhang, X.Y.; Wu, X.F.; Jia, W.C.; Pan, C.Y.; Li, X.C.; Lei, C.Z.; Chen, H.; Lan, X.Y. Novel nucleotide variations, haplotypes structure and associations with growth related traits of goat AT motif-binding factor (ATBF1) gene. Asian Australas. J. Anim. 2015, 28, 1394–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Młynek, K.; Oler, A.; Zielińska, K.; Tkaczuk, J.; Zawadzka, W. The effect of selected components of milk and ripening time on the development of the hardness and melting properties of cheese. Acta. Sci. Pol. Technol. Aliment. 2018, 17, 133–140. [Google Scholar] [PubMed]
- Pazzola, M.; Stocco, G.; Dettori, M.L.; Bittante, G.; Vacca, G.M. Effect of goat milk composition on cheesemaking traits and daily cheese production. J. Dairy. Sci. 2019, 102, 3947–3955. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, H.; Cheng, L.; Edwards, G.R.; Hickford, J.G. Effect of DGAT1 variant (K232A) on milk traits and milk fat composition in outdoor pasture-grazed dairy cattle. N. Z. J. Agric. Res. 2019, 1–13. [Google Scholar] [CrossRef]
- Xiang, R.; MacLeod, I.M.; Bolormaa, S.; Goddard, M.E. Genome-wide comparative analyses of correlated and uncorrelated phenotypes identify major pleiotropic variants in dairy cattle. Sci. Rep. 2017, 7, 9248. [Google Scholar] [CrossRef]
- Vacca, G.M.; Dettori, M.L.; Balia, F.; Luridiana, S.; Mura, M.C.; Carcangiu, V.; Pazzola, M. Sequence polymorphisms at the growth hormone GH1/GH2-N and GH2-Z gene copies and their relationship with dairy traits in domestic sheep (Ovis aries). Mol. Biol. Rep. 2013, 40, 5285–5294. [Google Scholar] [CrossRef]
- Bennett, T.; Desmond, A.; Harrington, M.; McDonagh, D.; Fitzgerald, R.J.; Flynn, A.; Cashman, K.D. The effect of high intakes of casein and casein phosphopeptide on calcium absorption in the rat. Br. J. Nutr. 2000, 83, 673–680. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.H.; Suh, G.H. Effect of the amount of body condition loss from the dry to near calving periods on the subsequent body condition change, occurrence of postpartum diseases, metabolic parameters and reproductive performance in Holstein dairy cows. Theriogenology 2003, 60, 1445–1456. [Google Scholar] [CrossRef]
- Roche, J.R.; Meier, S.; Heiser, A.; Mitchell, M.D.; Walker, C.G.; Crookenden, M.A.; Riboni, M.V.; Loor, J.J.; Kay, J.K. Effects of precalving body condition score and prepartum feeding level on production, reproduction, and health parameters in pasture-based transition dairy cows. J. Dairy Sci. 2015, 98, 7164–7182. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.L.; Friggens, N.C.; Ashworth, C.J.; Chagunda, M.G.G. Association between body energy content in the dry period and post-calving production disease status in dairy cattle. Animal 2017, 11, 1590–1598. [Google Scholar] [CrossRef] [Green Version]
- Abecassis, I.; Sedgewick, A.J.; Romkes, M.; Buch, S.; Nukui, T.; Kapetanaki, M.G.; Vogt, A.; Kirkwood, J.M.; Benos, P.V.; Tawbi, H. PARP1 rs1805407 increases sensitivity to PARP1 inhibitors in cancer cells suggesting an improved therapeutic strategy. Sci. Rep. 2019, 9, 3309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganapathipillai, S.; Laval, G.; Hoffmann, I.S.; Castejon, A.M.; Nicod, J.; Dick, B.; Frey, F.J.; Frey, B.M.; Cubeddu, L.X.; Ferrari, P. CYP11B2-CYP11B1 haplotypes associated with decreased 11 beta-hydroxylase activity. J. Clin. Endocrinol. Metab. 2005, 90, 1220–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J. Studies on Genetic Diversity of CSN1S1, BMPR-1B, BMP15 and IGFBP3 genes in two dairy goat breeds. Master’s Thesis, Northwest A&F University, Xianyang, China, 2006. [Google Scholar]
- Farias, F.H.G.; Dahlqvist, J.; Kozyrev, S.V.; Leonard, D.; Wilbe, M.; Abramov, S.N.; Alexsson, A.; Pielberg, G.R.; Hansson-Hamlin, H.; Andersson, G.; et al. A rare regulatory variant in the MEF2D gene affects gene regulation and splicing and is associated with a SLE sub-phenotype in Swedish cohorts. Eur. J. Hum. Genet. 2019, 27, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Markljung, E.; Jiang, L.; Jaffe, J.D.; Mikkelsen, T.S.; Wallerman, O.; Larhammar, M.; Zhang, X.; Wang, L.; Saenz-Vash, V.; Gnirke, A.; et al. ZBED6, a novel transcription factor derived from a domesticated DNA transposon regulates IGF2 expression and muscle growth. PLoS. Biol. 2009, 7, e1000256. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Thakran, P. Intron specificity in pre-mRNA splicing. Curr. Genet. 2018, 64, 777–784. [Google Scholar] [CrossRef]
- Hashiramoto, A.; Konishi, Y.; Murayama, K.; Kawasaki, H.; Yoshida, K.; Tsumiyama, K.; Tanaka, K.; Mizuhara, M.; Shiotsuki, T.; Kitamura, H.; et al. A variant of death-receptor 3 associated with rheumatoid arthritis interferes with apoptosis-induction of T cell. J. Biol. Chem. 2018, 293, 1933–1943. [Google Scholar] [CrossRef] [Green Version]
- Hecker, M.; Boxberger, N.; Illner, N.; Fitzner, B.; Schröder, I.; Winkelmann, A.; Dudesek, A.; Meister, S.; Koczan, D.; Lorenz, P.; et al. A genetic variant associated with multiple sclerosis inversely affects the expression of CD58 and microRNA-548ac from the same gene. PLoS. Genet. 2019, 15, e1007961. [Google Scholar] [CrossRef]

{kind=link}
| Breeds | Total Sizes | Genotype Frequencies (N) | Allele Frequencies | Population Parameters | χ2 (p Value) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| II | ID | DD | I | D | Ho | He | Ne | PIC | |||
| GZDG | 235 | 0.170 (40) | 0.429 (82) | 0.481 (113) | 0.345 | 0.655 | 0.548 | 0.452 | 1.824 | 0.350 | 12.173 (p = 4.85 × 10−4) |
| SBWC | 1092 | 0.295 (322) | 0.454 (496) | 0.251 (274) | 0.522 | 0.478 | 0.501 | 0.499 | 1.996 | 0.375 | 8.809 (p = 0.003) |
| HNBG | 278 | 0.255 (71) | 0.385 (107) | 0.360 (100) | 0.448 | 0.552 | 0.505 | 0.495 | 1.978 | 0.372 | 13.670 (p = 2.18 × 10−4) |
| Milk Performance Indices | Genotypes (N) | p Value | ||
|---|---|---|---|---|
| II (11) | ID (35) | DD (46) | ||
| Fat content (%) | 4.61 ± 0.40 | 4.36 ± 0.20 | 3.86 ± 0.15 | 0.103 |
| Protein content (%) | 3.93 a ± 0.21 | 3.73 ab ± 0.12 | 3.38 b ± 0.07 | 0.006 |
| TS content (%) | 13.73 a ± 0.57 | 13.38 ab ± 0.34 | 12.44 b ± 0.21 | 0.018 |
| SNF content (%) | 8.99 a ± 0.19 | 8.91 a ± 0.13 | 8.46 b ± 0.08 | 0.003 |
| Lactose content (%) | 3.88 ± 0.09 | 4.02 ± 0.08 | 3.95 ± 0.04 | 0.481 |
| Density | 1028.80 ± 0.45 | 1029.25 ± 0.46 | 1028.27 ± 0.27 | 0.144 |
| FPD (°C) | 0.50a ± 0.01 | 0.50 a ± 0.01 | 0.48 b ± 0.01 | 0.043 |
| Acidity (pH) | 7.49a ± 0.51 | 6.79 ab ± 0.33 | 6.22 b ± 0.19 | 0.046 |
| Breeds | Body Measurement Traits | Genotypes (N) | p Value | ||
|---|---|---|---|---|---|
| II (11) | ID (35) | DD (46) | |||
| GZDG | Body height (cm) | 67.98 c ± 3.09 | 69.05 b ± 2.89 | 71.26 a ± 0.90 | 0.009 |
| Body length (cm) | 75.54 b ± 0.98 | 77.24 a ± 1.18 | 77.60 a ± 0.68 | 0.043 | |
| Height at hip cross (cm) | 68.98 ± 2.91 | 69.33 ± 1.11 | 70.81 ± 0.95 | 0.571 | |
| Chest circumference (cm) | 89.18 ± 0.94 | 88.28 ± 0.98 | 88.92 ± 0.82 | 0.841 | |
| Chest width (cm) | 28.70 ± 0.85 | 27.81 ± 0.55 | 29.17 ± 0.49 | 0.180 | |
| Hip width (cm) | 21.94 ± 1.41 | 23.54 ± 0.59 | 22.26 ± 0.67 | 0.293 | |
| Cannon circumference (cm) | 11.34 ± 0.40 | 10.89 ± 0.21 | 10.30 ± 0.27 | 0.099 | |
| II (247) | ID (380) | DD (195) | |||
| SBWC | Body height (cm) | 56.62 c ± 0.29 | 58.11 b ± 0.19 | 60.12 a ± 0.33 | 1.61 × 10−16 |
| Body length (cm) | 66.42 b ± 0.31 | 67.95 a ±0.23 | 68.62 a ± 0.33 | 1.23 × 10−6 | |
| Height at hip cross (cm) | 59.41 c ± 0.29 | 60.66 b ± 0.21 | 61.97 a ± 0.30 | 4.22 × 10−9 | |
| Chest circumference (cm) | 84.97 b ± 0.49 | 86.78 a ± 0.35 | 86.93 a ± 0.50 | 0.003 | |
| Chest width (cm) | 19.59 b ± 0.18 | 20.36 a ±0.14 | 20.48 a ±0.18 | 3.90 × 10−4 | |
| Chest depth (cm) | 30.61 ± 01.16 | 30.15 ± 0.19 | 30.46 ± 0.18 | 0.851 | |
| Cannon circumference (cm) | 8.13 b ± 0.05 | 8.32 a ± 0.03 | 8.26 a ± 0.05 | 0.003 | |
| II (26) | ID (39) | DD (36) | |||
| HNBG | Body height (cm) | 51.50 b ± 0.89 | 53.63 a ± 0.64 | 53.98 a ± 0.65 | 0.049 |
| Body length (cm) | 54.71 ± 1.14 | 56.50 ± 0.75 | 57.55 ± 0.64 | 0.069 | |
| Body weight (kg) | 26.79 b ± 1.48 | 29.35 ab ± 1.17 | 31.34 a ± 1.00 | 0.043 | |
| Chest circumference (cm) | 71.04 ± 1.39 | 73.00 ± 1.11 | 74.40 ± 0.90 | 0.137 | |
| Chest width (cm) | 15.10 ± 0.33 | 15.61 ± 0.33 | 15.51 ± 0.26 | 0.521 | |
| Chest depth (cm) | 25.83 b ± 0.47 | 26.67 ab ± 0.34 | 27.28 a ± 0.32 | 0.033 | |
| Hip width (cm) | 13.26 ± 0.35 | 13.51 ± 0.26 | 14.10 ± 0.17 | 0.069 | |
| Cannon circumference (cm) | 7.92 ± 0.13 | 8.08 ± 0.10 | 8.01 ± 0.09 | 0.621 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Wang, K.; Liu, J.; Zhu, H.; Qu, L.; Chen, H.; Lan, X.; Pan, C.; Song, X. An 11-bp Indel Polymorphism within the CSN1S1 Gene Is Associated with Milk Performance and Body Measurement Traits in Chinese Goats. Animals 2019, 9, 1114. https://doi.org/10.3390/ani9121114
Zhang Y, Wang K, Liu J, Zhu H, Qu L, Chen H, Lan X, Pan C, Song X. An 11-bp Indel Polymorphism within the CSN1S1 Gene Is Associated with Milk Performance and Body Measurement Traits in Chinese Goats. Animals. 2019; 9(12):1114. https://doi.org/10.3390/ani9121114
Chicago/Turabian StyleZhang, Yanghai, Ke Wang, Jinwang Liu, Haijing Zhu, Lei Qu, Hong Chen, Xianyong Lan, Chuanying Pan, and Xiaoyue Song. 2019. "An 11-bp Indel Polymorphism within the CSN1S1 Gene Is Associated with Milk Performance and Body Measurement Traits in Chinese Goats" Animals 9, no. 12: 1114. https://doi.org/10.3390/ani9121114