1. Introduction
Over the years, human activities have severely affected wildlife and its habitat; therefore, it is crucial to balance the needs of people and wildlife. In particular, the habitats of birds are extensively exploited by humans through the use of land and water for agriculture, associated construction, and other types of development activities [
1,
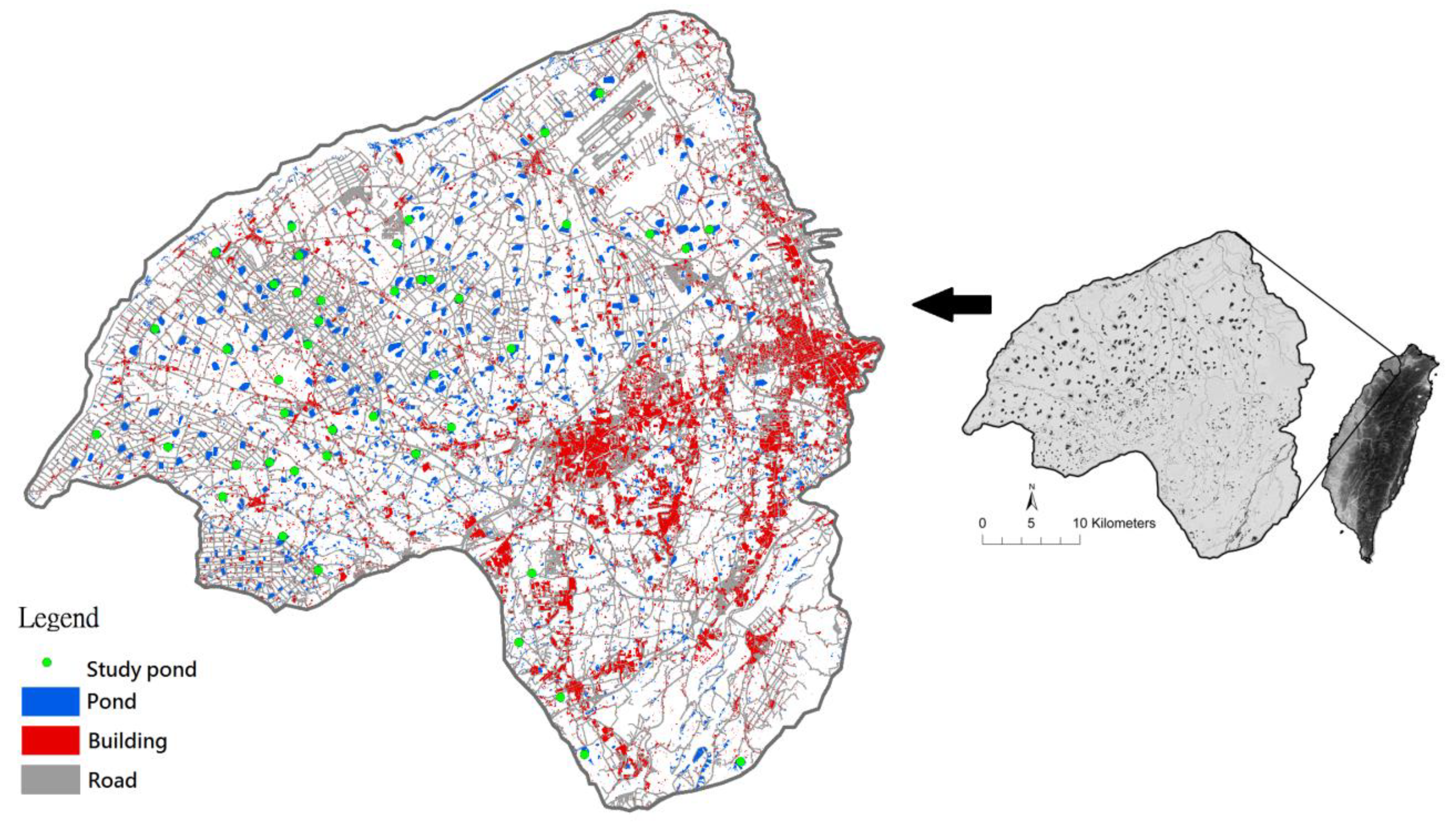
2]. In Taoyuan Tableland, Taiwan, thousands of farm ponds have been constructed for irrigation. This area had more than 3290 ponds in the 1970s, but fewer than 1800 currently exist. Farm pond configurations and the complexity of their compositions exhibit marked diversity [
3]. Pondscape was defined as “a series of water surfaces of ponds in association with various surrounding landforms, including farms, creeks, canals, roads, houses, woodlands, and other open spaces” [
3,
4,
5]. Several migratory birds stop over at pondscapes. Because of their specific habitat requirements for stopover during migration, birds provide indicators regarding habitat conditions [
6,
7]. All avian species select a suitable habitat to ensure availability of food, water, shelter from the weather and predators and feasible nesting sites to reproduce. All species in a guild display similar characteristics. However, Duelli and Obrist suggested that generalist species might not be appropriate biodiversity indicators [
8].
Anthropogenic influences (i.e., pollution, destruction, degradation, and other stress) can be monitored using bioindicators [
9,
10,
11]. In this case, generalists are the ones that could benefit from a higher abundance of habitats that are spatially heterogeneous. However, specialists are the ones that thrive in a nearly homogenous habitat, with a high occurrence rate in their own specific habitat [
12]. Thus, compared with generalists, specialists are less dependent on habitat scale and can only exist within a specific type of habitat. These species include waterfowl (Anatidae and Podicipedidae), shorebirds (Charadriidae and Scolopacidae), and wading birds (Ardeidae). They have specific habitat requirements and are generally unable to adapt to new diets or environmental conditions [
13]. Therefore, specialists are more vulnerable than generalists to anthropogenic disturbance [
14,
15].
The guild concept involves the division of birds according to their habitats and further categorization according to landscape configurations. Root, the first avian scientist to propose the guild concept, defined a guild as “a group of species that exploit the same class of environmental resources in a similar way” [
16]. He realized that the traditional taxonomic approach failed to categorize avian communities appropriately. For example, he described the “foliage-gleaning guild” as birds that obtain their food from foliage and occasionally from branches [
16]. Thus, Root grouped five species with similar diets, foraging locations, and feeding behaviors into one guild [
16].
After Root defined functional groups based on traditional guilds (considering diets and foraging strategies), other authors have followed his approach for investigating avian behavior and foraging strategies [
17,
18,
19,
20]. Studies have evaluated nesting, resting, singing, residential locations [
3,
21,
22,
23], foraging strategies, and singing locations [
24]. However, most studies using functional groups have grouped species according to subjective criteria or a single behavior, focused on a single group or selected groups, at a single location or on small spatial scales [
25,
26].
Previous studies have also evaluated environmental conditions through examining guilds within heterogeneous landscapes [
12,
27]. We selected the definition of a guild that uses habitat preference to define functional groups [
4]. Through categorization of birds as generalists or specialists, French and Picozzi [
4] demonstrated that wintering birds were influenced by land use. Avian grouping aids in identifying avian diversity according to habitat, while tackling landscape complexity [
28]. Because information is limited regarding environmental factors that affect avian guilds, previous avian studies have applied cluster analyses for the grouping of similar components of the avian community into respective functional groups [
29,
30,
31]. The previous study aimed to identify groups of birds with certain habitat preferences by constructing groups (clusters) using multivariate data [
30]. Both habitat- and landscape-scale avian community studies are required to understand habitat selection [
32,
33].
On a larger scale, landscape configurations account for variations in the richness and diversity of wintering bird species. An irrigation birds’ habitat can be evaluated according to the number of avian species it contains. Therefore, birds become a bio-indicator for different types of habitat [
34,
35]. Differences in edge disturbance affect birds differently on avian communities. To preserve biodiversity on different habitats and landscape scales, it is essential to understand the effects of different management strategies on diversity.
Avian ecologists have used guilds to avoid classified errors that can occur when considering a large number of species [
36]. However, the taxonomic diversity of entire groups and specific guilds is well debated [
37]. One drawback of using guilds is that the taxonomic approach to avian studies is not commensurate with landscape scales [
37]. Furthermore, studies using aggregate species richness or diversity indices have often been oversimplified [
38,
39].
Although most of the farm ponds in Taoyuan are artificial, they provide food, refuge, and nesting sites for breeding birds. Thus, farm pond ecosystems in Taoyuan are crucial for these birds. Furthermore, farm ponds are wetlands for flood detention and water purification [
40,
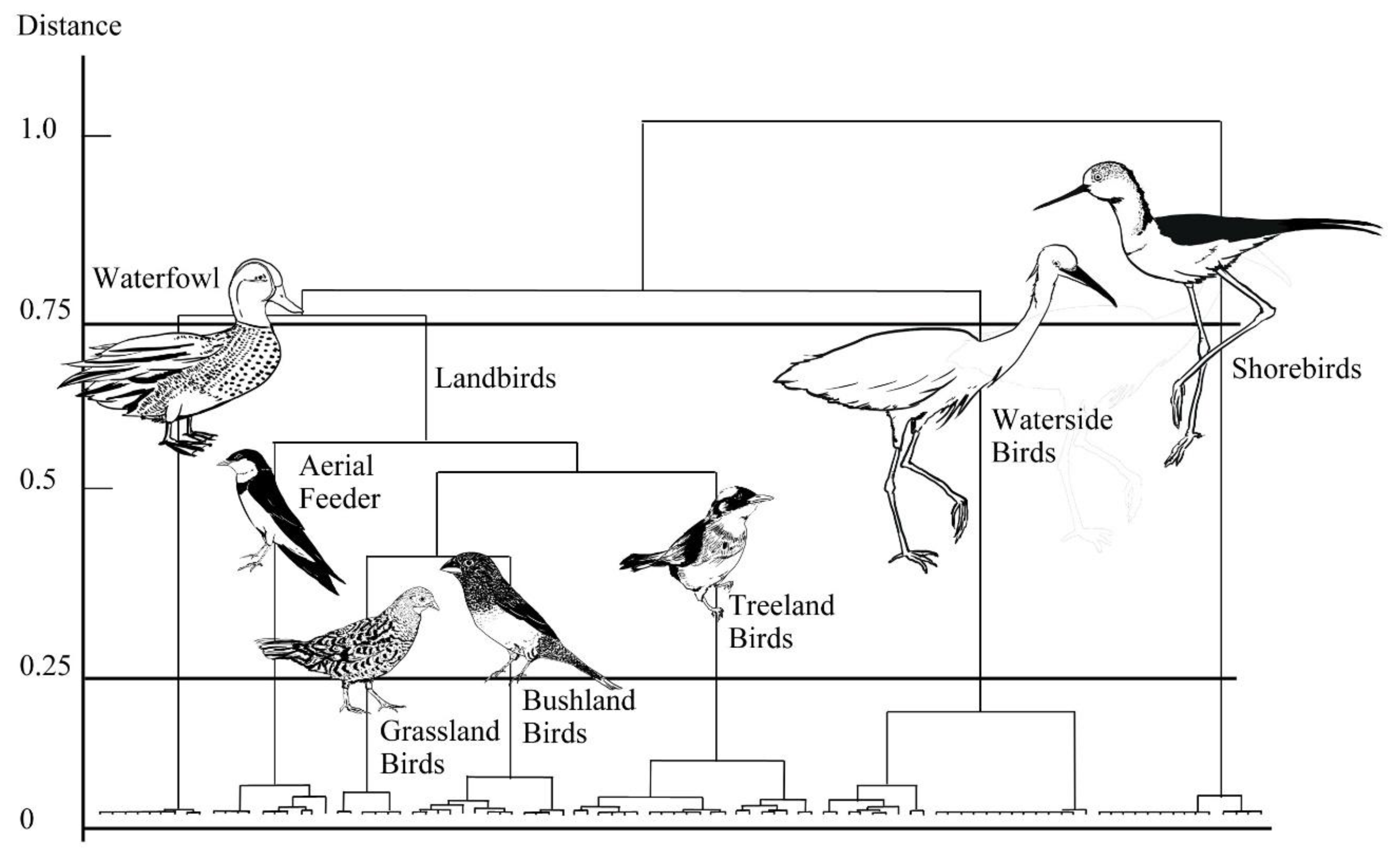
41]. Farm ponds are vital for humans as well. However, current developments, such as large-scale solar facilities, in ponds tend to destroy pondscapes; therefore, we, ecologists and bird lovers, are actively trying to protect these ponds. This study includes data from 2003 to 2004; our goal was to present bird conditions before solar facility construction to inform decision-makers of the importance of ponds to wildlife. In this study, we compared approaches for calculating species diversity in specific functional groups, which helped select an approach for fitting avian communities in irrigation ponds. The purpose of this study was to identify bird guilds through cluster analysis, simply list the birds recorded, organize them into groups, and indicate those understood to be “generalists” or “specialists”. Therefore, this study aimed to (1) characterize and analyze the waterbird species around the irrigation ponds of Taoyuan and, (2) categorize their functional groups using cluster analysis, thereby grouping birds according to different habitats. We did not study avian feeding habits (i.e., insect feeding, seed feeding, algae feeding, fish or crustacean feeding) and their food; we instead focused only on the correlation between guild (i.e., species richness and individuals) and pondscape variables.
4. Discussion
We compared wetland birds (i.e., waterfowl, shorebirds, and wading species) with landbirds (i.e., woodland birds, bushland birds, and grassland birds) to analyze their associations with internal and external pond variables, and the results demonstrated that the individual numbers of waterbirds were more strongly correlated with the pond variables than were the landbirds. Guild analyses further suggested that the principal factor affecting individual birds’ habitat selection was habitat availability. Therefore, the potential for environmental effects on the bird community is high for waterbirds, particularly waterside birds. For individual waterbirds to persist in the agricultural pondscape, fields should have large areas and well-designed neighboring landscapes to support such birds.
Our study results indicated that different avian guilds respond differently to environmental changes. In general, ponds with a larger area for wetland birds and wintering migratory birds could sustain population increases. The associations between population sizes and carrying capacity for individual numbers and species richness of a pond were moderate and slight, respectively, with ponds that support large concentrations of aerial feeders and landbirds as possible exceptions. However, the point at which a pond is regarded as saturated by a single species can be determined by the population sizes of other species wintering on those ponds. Our study results indicated a significant positive correlation between species richness and abundance in several guilds of waterbirds.
In this study, we compared the species-area relationships in ecological groups with similar source pool sizes. These analyses restricted the guild sizes to reduce the possibility of confounding habitat effects. The absence of a habitat of a suitable size was likely a key factor leading to the poor responses of some species in the selection of their wintering ponds. The entire habitats included WASA, MA, and FCA. The waterbirds dependent on large habitat sizes increased in number in more spatially heterogeneous areas, probably as a result of increased safety and food supply. However, other environmental factors, such as the presence of predators and availability of food may also have altered the habitat preferences of waterbirds in the study area.
First, we observed that the species richness and abundance of the wading birds were associated with PS. The total species richness and number of individual waterside birds increased with increasing PS. Second, we observed that individual waterfowl were correlated with the FCA because these specialists or interior species were more sensitive to disturbance than generalists or edge species. Migrants (family Anatidae) and residents (family Podicipedidae) tended to be more sensitive. We discovered that their habitats are far away from the road, farm, or other nonwater regimes. They appeared to be influenced by the level of human disturbance, PS, windbreak size, and pond edge length.
Compared to previous work, we found our study reveal similar result is different pond factors such as depth, size, and vegetation would influence the bird assemblages [
54]. Our study is similar in terms of findings with Froneman et al. (2001), they surveyed 59 farm ponds and found 44 bird species in the Elgin and Caledon districts of the Western Cape, South Africa [
5]. Compared to their study, we found 94 species in Taoyuan Tableland. Thus, this shows that the farm ponds in Taoyuan are really important for the wintering birds. Besides, the same result is that they found the surface area of the farm ponds as an important variable determining the presence and abundance of many waterbird species [
5]. Different from their study, our study included the human structure and found near the buildings and roads, the bird counts and diversity decreased. Anyway, the most important conclusion is that we totally agree artificial waterbodies can play as alternative refuges for biodiversity [
55,
56,
57,
58,
59].
In our study, we expected to observe the area per se hypothesis within an intermediate range of areas but not at all spatial scales. On a small spatial scale, the species–area relationship is not governed by an equation but is curvilinear on a log-log plot. On a landscape scale, the species-area relationship bends upward toward a limiting slope of unity [
60]. We realized that the habitat preferences of birds with different lifestyles must be considered when determining habitat suitability. Most species in small patches associated with the surrounding landscape are generalists, choosing between major habitats and edge habitats. In large patches, the specialists select only interior habitats [
61,
62]. Therefore, the spatially and taxonomically different species differ in their size [
63]. Different avian communities are likely to yield different land-use patches.
In this study, we compared the species-pondscape relationships among ecological groups with their surrounding areas. We restricted these analyses to guild pool sizes to limit the confounding effects of areas. The variables of local determinants of community structures were associated with the amount of farmlands as well as the amount of urban environments. Low-rise residential houses and high-density apartments were observed to affect species richness.
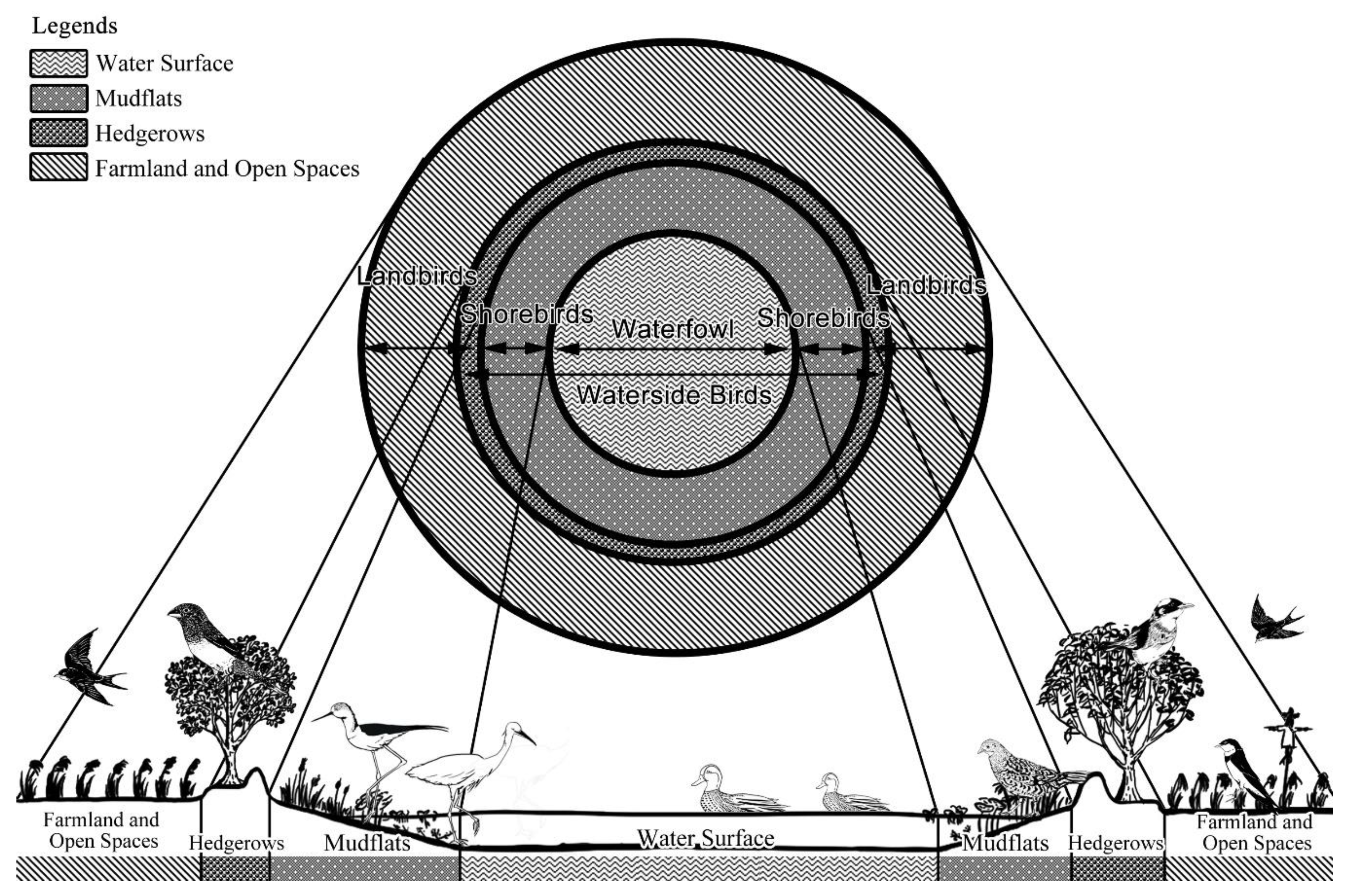
Waterside bird richness displayed a correlation with %BUILD within a radius of 100 ha from the pond’s geometric center because the specialists detected from the pond’s core to the waterfront were more sensitive to anthropogenic disturbance than generalists (i.e., landbirds) were (
Table 4). The wading bird species displayed a correlation with %FARM within a radius of 100 ha from the pond’s geometric center. The richness of the waterside bird guild was correlated negatively with increasing urbanization level (indicated by %BUILD); however, the richness correlated positively with increasing green spaces (i.e., farmlands and grasslands). Because we combined environmental factors such as water and edge species of different sizes, foraging modes, and trees from the pond’s core to the waterfront, it is likely that the increase in anthropogenic areas was the principal reason for their decline. In addition, the farmlands, which might translate to greater insect abundance, were strongly correlated with wading bird abundance.
Investigators extensively debated the field domains of the area per se hypothesis and species–habitat hypothesis [
64]. However, the generalized principles of ecological designs have yet to be determined, and there is no final consensus on which species-habitat hypothesis is more relevant. As described, birds respond to food and roost sites during habitat selection. The numbers of individual birds of a particular species have been correlated with the requirements for grasslands, mudflats, open shorelines, and canopies or water surfaces for horizontal heterogeneity [
65,
66]. Therefore, bird-habitat relationships result from the responses of birds using habitats for different activities, such as foraging, molting (i.e., that of the mute swan and greylag goose) [
67,
68], and roosting in winter. Birds can select pondscape configurations according to their preference.
In this study, the irrigation pond areas were dominated by fields separated by hedgerows and windbreaks, and woods of various sizes were scattered in these areas. The distributions of avian species within such mosaic landscapes were discontinuous, depending on the preferred habitat locations, density-dependent processes, and quality of individual patches. These configurations are surrounded by built-up areas, rivers, roads, and farmlands. In our pondscape evaluations, we used the selected parameters to measure the spatial arrangement of wooded and aquatic landscapes as well as to evaluate the significance of their differences. Previous studies have used these parameters to measure temporal changes in actual landscapes and changes in intensively used landscapes [
69,
70]. Increasing the pond area increases the pond core area, thereby benefiting specialist species by enhancing the population persistence associated with water depth, water level fluctuation, vegetation, salinity, topography, food type, food accessibility, size, and connectivity [
14,
15,
19]. Our results indicated a requirement for relevant conservation scenarios to focus on vulnerable sites, which might be targeted for enlargement by habitat creation at their woody edges, on the basis that large pond habitats are broadly beneficial for biodiversity. Thus, we suggest a study on yearly or interannual variability of wintering waterbirds to analyze the effect of habitat changes on birds.
5. Conclusions
To construct waterbird refuges in Taoyuan, Taiwan, for securing habitats for wintering waterbirds in the areas of anthropogenic influence, existing and potential irrigation ponds must be identified. Pond conservation for bird refuges is difficult because of increasing urban development, which exerts pressure on avian communities [
43,
71]. Changes in land use [
72], particularly consolidating farming practices with urban construction, affect avian communities in ponds. Evaluating the pond habitats of winter birds might provide useful information for simulating the pond environment to identify the criteria of their habitat selection behaviors. On the habitat and landscape scales, specific selection according to avian assemblages is required for identifying large areas within fields to support various waterbird species.
The number of farm ponds in Taoyuan Tableland is decreasing, thus providing evidence of the negative effects of landscape quality on the avian distribution and species in these fragmented habitats. In our avian population studies, the functional groups associated with pondscape configurations provided an effective tool for determining linear relationships based on the concept of landscape ecology. We used theoretical models to quantify the influence of landforms on avian groups. Our study provided substantial evidence that artificial ponds also influence wintering waterbirds. The final results regarding ponds may aid stakeholders and land managers to identify areas for the establishment of large-scale solar facilities in pond areas for superior management of wildlife. In conclusion, our study provided a comprehensive view of farm pond-bird interaction in Taoyuan Tableland, thus aiding decision-makers in enacting policies beneficial to both humans and the environment.
The results related to the condition of the birds at farm ponds in Taoyuan Tableland from 2003 to 2004. We, therefore, can compare the current condition of birds with that of 15 years ago for a better understanding of the relationships of human activities and wildlife biodiversity.




{kind=link}
{kind=link}
{kind=link}


