Molecular Characterization of Sarcocystis Species Isolated from Sheep and Goats in Riyadh, Saudi Arabia

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Macroscopic Analysis
2.3. Microscopic Analysis
2.4. Digestion Method
2.5. Transmission Electron Microscopy (TEM)
2.6. Molecular Analysis
2.6.1. DNA Extraction and PCR Amplification
2.6.2. DNA Sequencing and Phylogenetic Analysis
2.7. Statistical Analysis
3. Results
3.1. Prevalence of Natural Infection
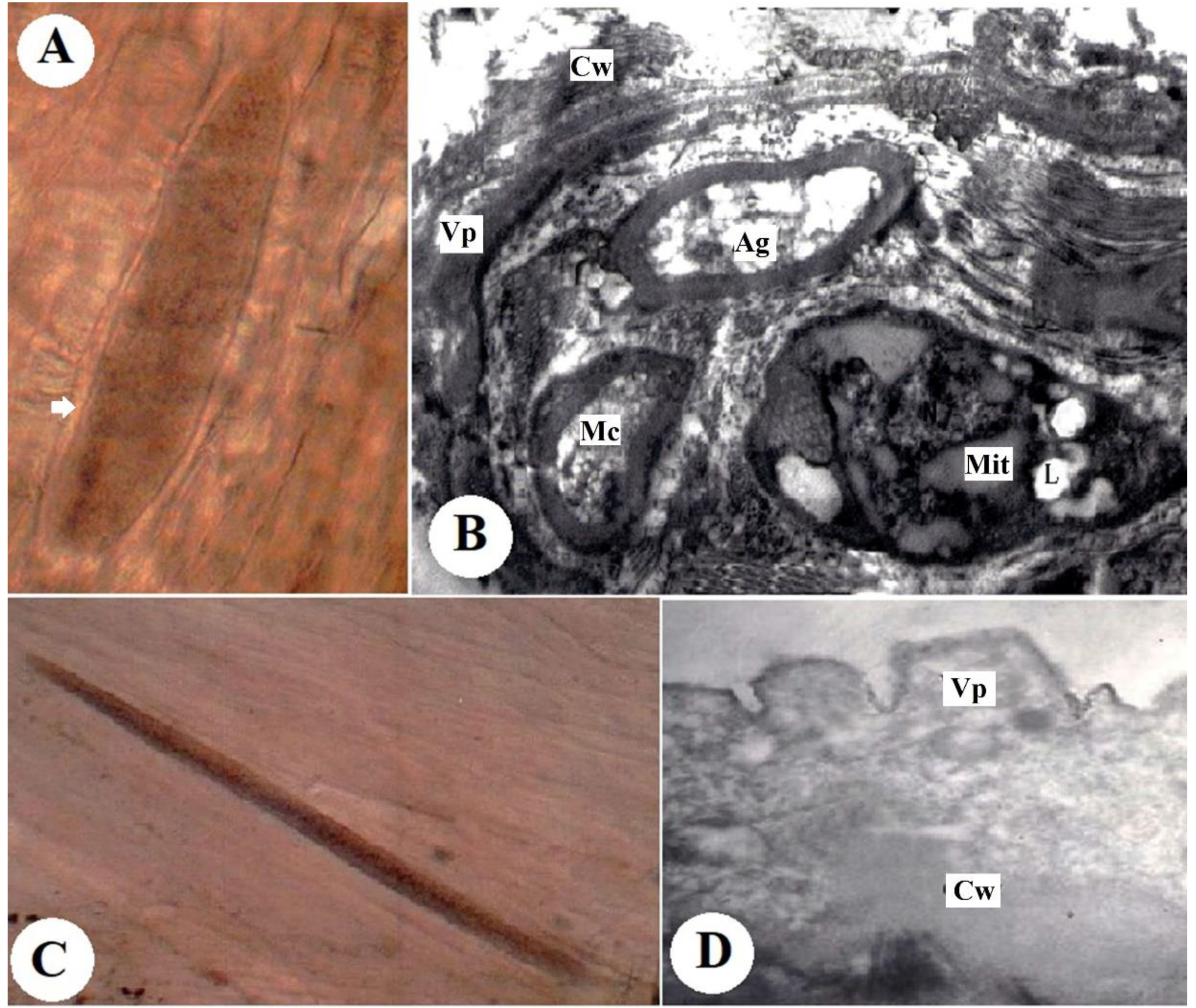
3.2. Morphological Characteristics of the Cysts
3.2.1. S. tenella Cysts
3.2.2. S. capracanis Cysts
3.3. Molecular Analysis
3.4. Molecular Characterization of the COX1 Gene
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Abdel-Ghaffar, F.; Mehlhorn, H.; Bashtar, A.R.; Al-Rasheid, K.; Sakran, T.; El-Fayoumi, H. Life cycle of Sarcocystis camelicanis infecting the camel (Camelus dromedarius) and the dog (Canis familiaris), light and electron microscopic study. Parasitol. Res. 2009, 106, 189–195. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Heydorn, A.O. The Sarcosporidia (Protozoa, Sporozoa): Life cycle and fine structure. In Advances in Parasitology; Academic Press: Cambridge, MA, USA, 1978; pp. 43–91. [Google Scholar]
- Ghaffar, F.A.; Hilali, M.; Scholtyseck, E. Ultrastructural study of Sarcocystis fusiformis (Railliet, 1897) infecting the Indian water buffalo (Bubalus bubalis) of Egypt. Trop. Parasitol. 1978, 29, 289–294. [Google Scholar]
- Dubey, J.P.; Kistner, T.P.; Callis, G. Development of Sarcocystis in mule deer transmitted through dogs and coyotes. Can. J. Zool. 1983, 61, 2904–2912. [Google Scholar] [CrossRef]
- Entzroth, R. Invasion and early development of Sarcocystis muris (Apicomplexa, Sarcocystidae) in tissue cultures. J. Protozol. 1985, 32, 446–453. [Google Scholar] [CrossRef]
- Abdel-Ghaffar, F.; Bashtar, A.R.; El-Sayed, M. Electron microscopic studies on Sarcocystis infection in sheep in Upper Egypt. Bull. Fac. Sci. Cairo Univ. 1990, 58, 33–49. [Google Scholar]
- Abdel-Ghaffar, F.; Al-Johany, A. A light and electron microscope study of Sarcocystis mitrani (sp. nov.) infecting the skink Scincus mitranus in the central region of Saudi Arabia. Parasitol. Res. 2002, 88, 102–106. [Google Scholar] [CrossRef]
- Collins, G.H.; Atkinson, E.; Charleston, W.A. Studies on Sarcocystis species III: The macrocystic species of sheep. N. Z. Vet. J. 1979, 27, 204–206. [Google Scholar] [CrossRef]
- Heydorn, A.O.; Mehlhorn, H. Fine structure of Sarcocystis arieticanis Heydorn, 1985 in its intermediate and final hosts (sheep and dog). Zentralbl. Bakteriol. Mikrobiol. Hyg. Ser. A 1987, 264, 353–362. [Google Scholar] [CrossRef]
- Heydorn, A.O.; Gestrich, R.; Mehlhorn, H.; Rommel, M. Proposal for a new nomenclature of the Sarcosporidia. Parasitol. Res. 1975, 48, 73–82. [Google Scholar] [CrossRef]
- Heydorn, A.O.; Matuschka, F.R. Zur Endwirtspezifität der vom Hund übertragenen Sarkosporidienarten, Final host specificity of Sarcocystis species transmitted by dogs. Zeitschrift für Parasitenkunde 1981, 66, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Al Quraishy, S.; Morsy, K.; Bashtar, A.R.; Ghaffar, F.A.; Mehlhorn, H. Sarcocystis arieticanis (Apicomplexa: Sarcocystidae) infecting the heart muscles of the domestic sheep, Ovis aries (Artiodactyla: Bovidae), from KSA on the basis of light and electron microscopic data. Parasitol. Res. 2014, 113, 3823–3831. [Google Scholar] [CrossRef]
- Hussein, H.S. The prevalence of Sarcocystis infection in Saudi Arabia Najdi sheep and camels. Biol. Sci. 1991, 1, 43–56. [Google Scholar]
- Hilali, M.; Fatani, A.; Al-Atiya, S. Isolation of tissue cysts of Toxoplasma, Isospora, Hammondia and Sarcocystis from camel (Camelus dromedarius) meat in Saudi Arabia. Vet. Parasitol. 1995, 58, 353–356. [Google Scholar] [CrossRef]
- Fatani, A.; Hilali, M.; Al-Atiya, S.; Al-Shami, S. Prevalence of Sarcocystis in camels (Camelus dromedarius) from Al-Ahsa, Saudi Arabia. Vet. Parasitol. 1996, 62, 241–245. [Google Scholar] [CrossRef]
- Shazly, M.A. Light and electron microscopic studies on Sarcocystis infecting the Dromedaries in Saudi Arabia. Egypt. J. Zool. 2000, 35, 273–285. [Google Scholar]
- Mohammed, O.B. Prevalence, Identity and Phylogeny of Sarcocystis Parasites from Gazelles in Saudi Arabia. Ph.D. Thesis, Kingston University, London, UK, 2000. [Google Scholar]
- da Silva, R.C.; Su, C.; Langoni, H. First identification of Sarcocystis tenella (Railliet, 1886) Moule, 1886 (Protozoa: Apicomplexa) by PCR in naturally infected sheep from Brazil. Vet. Parasitol. 2009, 165, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Gjerde, B.; Luzón, M.; Alunda, J.M.; de la Fuente, C. Morphological and molecular characteristics of six Sarcocystis species from red deer (Cervus elaphus) in Spain, including Sarcocystis cervicanis and three new species. Parasitol. Res. 2017, 116, 2795–2811. [Google Scholar] [CrossRef]
- Rudaitytė-Lukošienė, E.; Prakas, P.; Butkauskas, D.; Kutkienė, L.; Vepštaitė-Monstavičė, I.; Servienė, E. Morphological and molecular identification of Sarcocystis species from the sika deer (Cervus nippon), including two new species Sarcocystis frondea and Sarcocystis nipponi. Parasitol. Res. 2018, 117, 1305–1315. [Google Scholar] [CrossRef]
- Gjerde, B. Phylogenetic relationships among Sarcocystis species in cervids, cattle and sheep inferred from the mitochondrial cytochrome c oxidase subunit I gene. Int. J. Parasitol. 2013, 43, 579–591. [Google Scholar] [CrossRef]
- GJERDE, B. Sarcocystis species in red deer revisited: With a re-description of two known species as Sarcocystis elongata n. sp. and Sarcocystis truncata n. sp. based on mitochondrial COX1 sequences. Parasitology 2014, 141, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Bittencourt, M.V.; Meneses, I.D.; Ribeiro-Andrade, M.; de Jesus, R.F.; de Araújo, F.R.; Gondim, L.F. Sarcocystis species in sheep and goats: Frequency of infection and species identification by morphological, ultrastructural, and molecular tests in Bahia, Brazil. Parasitol. Res. 2016, 115, 1683–1689. [Google Scholar] [CrossRef]
- Odening, K.; Stolte, M.; Bockhardt, I. On the diagnostics of Sarcocystis in cattle: Sarcocysts of a species unusual for Bos taurus in a dwarf zebu. Vet. Parasitol. 1996, 66, 19–24. [Google Scholar] [CrossRef]
- Dubey, J.P.; Speer, C.A.; Charleston, W.A. Ultrastructural differentiation between sarcocysts of Sarcocystis hirsuta and Sarcocystis hominis. Vet. Parasitol. 1989, 34, 153–157. [Google Scholar] [CrossRef]
- Hamidinejat, H.; Moetamedi, H.; Alborzi, A.; Hatami, A. Molecular detection of Sarcocystis species in slaughtered sheep by PCR–RFLP from south-western of Iran. J. Parasit. Dis. 2014, 38, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Al-Qureishy, S.A. Sarcocystis Parasites (Coccidia, Apicomplexa) Infecting Some Slaughtered Animals in Riyadh City, Saudi Arabia. Ph.D. Thesis, King Saud University, Riyadh, Saudi Arabia, 2005. [Google Scholar]
- Dubey, J.P.; Lindsay, D.S.; Speer, C.A.; Fayer, R.; Livingston, C.W., Jr. Sarcocystis arieticanis and other Sarcocystis species in sheep in the United States. J. Parasitol. 1988, 74, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Amairia, S.; Amdouni, Y.; Rouatbi, M.; Rjeibi, M.R.; Awadi, S.; Gharbi, M. First detection and molecular identification of Sarcocystis species in small ruminants in North-West Tunisia. Transbound. Emerg. Dis. 2018, 65, 441–446. [Google Scholar] [CrossRef]
- Whaeeb, S.T.; Khalaf, A.A.F.I.A. Molecular Study of Four Species of Sarcocystis Isolated from Sheeps Esophagus and Inter Skeleton Muscle in Baghdad. WJPR 2016, 5, 158–165. [Google Scholar]
- Heydorn, A.O.; Haralambidis, S. Development of Sarcocystis capracanis Fischer, 1979. Berl. Munch. Tierarztl. Wochenschr. 1982, 95, 265–271. [Google Scholar]
- Dubey, J.P.; Calero-Bernal, R.; Rosenthal, B.M.; Speer, C.A.; Fayer, R. Sarcocystosis of Animals and Humans; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Saito, M.; Shibata, Y.; Kobayashi, T.; Kobayashi, M.; Kubo, M.; Itagaki, H. Ultrastructure of the cyst wall of Sarcocystis species with canine final host in Japan. J. Vet. Med. Sci. 1996, 58, 861–867. [Google Scholar] [CrossRef]
- Claveria, F.G.; Pedro-Lim, M.R.S.; Tan, J.E.; Flores-Cruz, M.J. Sarcocystis capracanis infection in Philippine domestic goats (Capra hircus): Ultrastructural studies. Philipp. J. Sci. 2004, 133, 33–37. [Google Scholar]
- Morsy, K.; Saleh, A.; Al-Ghamdi, A.; Abdel-Ghaffara, F.; Al-Rasheid, K.; Bashtar, A.R.; Al Quraishy, S.; Mehlhorn, H. Prevalence pattern and biology of Sarcocystis capracanis infection in the Egyptian goats: A light and ultrastructural study. Vet. Parasitol. 2011, 181, 75–82. [Google Scholar] [CrossRef]
- Hu, J.J.; Liu, T.T.; Liu, Q.; Esch, G.W.; Chen, J.Q.; Huang, S.; Wen, T. Prevalence, morphology, and molecular characteristics of Sarcocystis species in domestic goats (Capra hircus) from Kunming, China. Parasitol. Res. 2016, 115, 3973–3981. [Google Scholar] [CrossRef] [PubMed]
- Vlemmas, I.; Kanakoudis, G.; Tsangaris, T.H.; Theodorides, I.; Kaldrymidou, E. Ultrastructure of Sarcocystis tenella (Sarcocystis ovicanis). Vet. Parasitol. 1989, 33, 207–217. [Google Scholar] [CrossRef]
- Formisano, P.; Aldridge, B.; Alony, Y.; Beekhuis, L.; Davies, E.; Del Pozo, J.; Dunn, K.; English, K.; Morrison, L.; Sargison, N.; et al. Identification of Sarcocystis capracanis in cerebrospinal fluid from sheep with neurological disease. Vet. Parasitol. 2013, 193, 252–255. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Genes | Primers | Sequences | Fragment Size | References |
|---|---|---|---|---|
| COX1 | SF1 | ATGGCGTACAACAATCATAAAGAA | 1100 bp | [21,22] |
| SR9 | ATATCCATACCRCCATTGCCCAT |
| Animals | No. Examined (%) | No. Infected (%) |
|---|---|---|
| Sheep | Niemy 108 | 38 (35.18%) |
| Najdy 90 | 39 (43.33%) | |
| Sawakny 32 | 14 (43.75%) | |
| 230 | 91 (39.56%) | |
| Goats | 84 | 36 (42.85%) |
| Total | 314 | 127 (40.44%) |
| Categories | Sheep | Goats | ||
|---|---|---|---|---|
| No. Examined | No. Infected (%) | No. Examined | No. Infected (%) | |
| Young males (under 1 year) | 147 | 64 (43.53%) | 59 | 28 (47.45%) |
| Adult males (above 1 year) | 53 | 19 (35.84%) | 5 | 2 (40%) |
| Young females (under 1 year) | 21 | 9 (42.85%) | 17 | 6 (35.29%) |
| Adult females (above 1 year) | 9 | 3 (33.33%) | 3 | 0 (0%) |
| Total | 230 | 95 (41.30%) | 84 | 36 (42.85%) |
| Animals | No. Infected | Esophagus | Tongue | Diaphragm | Heart | Skeletal Muscles |
|---|---|---|---|---|---|---|
| No. (%) | No. (%) | No. (%) | No. (%) | No. (%) | ||
| Niemy | 38 | 11 | 14 | 26 | 23 | 32 |
| (28.94%) | (36.84%) | (68.42%) | (60.52%) | (84.21%) | ||
| Najdy | 39 | 8 | 5 | 24 | 14 | 17 |
| (20.51%) | (12.82%) | (61.53%) | (35.89%) | (43.58%) | ||
| Sawakny | 14 | 5 | 3 | 12 | 8 | 11 |
| (35.71%) | (21.42%) | (85.71%) | (57.14%) | (78.57%) | ||
| Goats | 36 | 9 | 7 | 21 | 17 | 22 |
| (25%) | (19.44%) | (58.33%) | (47.22%) | (61.11%) | ||
| Total | 127 | 33 | 29 | 83 | 62 | 82 |
| (25.98%) | (22.83%) | (65.35%) | (48.81%) | (64.56%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Metwally, D.M.; Al-Damigh, M.A.; Al-Turaiki, I.M.; El-Khadragy, M.F. Molecular Characterization of Sarcocystis Species Isolated from Sheep and Goats in Riyadh, Saudi Arabia. Animals 2019, 9, 256. https://doi.org/10.3390/ani9050256
Metwally DM, Al-Damigh MA, Al-Turaiki IM, El-Khadragy MF. Molecular Characterization of Sarcocystis Species Isolated from Sheep and Goats in Riyadh, Saudi Arabia. Animals. 2019; 9(5):256. https://doi.org/10.3390/ani9050256
Chicago/Turabian StyleMetwally, Dina M., Mashael A. Al-Damigh, Isra M. Al-Turaiki, and Manal F. El-Khadragy. 2019. "Molecular Characterization of Sarcocystis Species Isolated from Sheep and Goats in Riyadh, Saudi Arabia" Animals 9, no. 5: 256. https://doi.org/10.3390/ani9050256
APA StyleMetwally, D. M., Al-Damigh, M. A., Al-Turaiki, I. M., & El-Khadragy, M. F. (2019). Molecular Characterization of Sarcocystis Species Isolated from Sheep and Goats in Riyadh, Saudi Arabia. Animals, 9(5), 256. https://doi.org/10.3390/ani9050256