Life-History Traits of the Black Soldier Fly, Hermetia illucens (L.) (Diptera: Stratiomyidae), Reared on Three Manure Types


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Acquisition of BSF
2.2. Acquisition of Manure
Chemical Composition of Manure
2.3. Experiment Design
Immature and Adult Life-History Traits
2.4. Bioconversion of Manure
2.5. Percent DM Reduction
2.6. Protein Content of Larvae
2.7. Statistical Analysis
3. Results
3.1. Moisture Content of Manure
3.2. Immature and Adult Life-History Traits
3.2.1. Final Larval Weight
3.2.2. Time to first prepupation
3.2.3. Prepupal Weight
3.2.4. Percent Prepupation
3.2.5. Adult Male Weight
3.2.6. Adult Male Longevity
3.2.7. Adult Female Weight
3.2.8. Adult Female Longevity
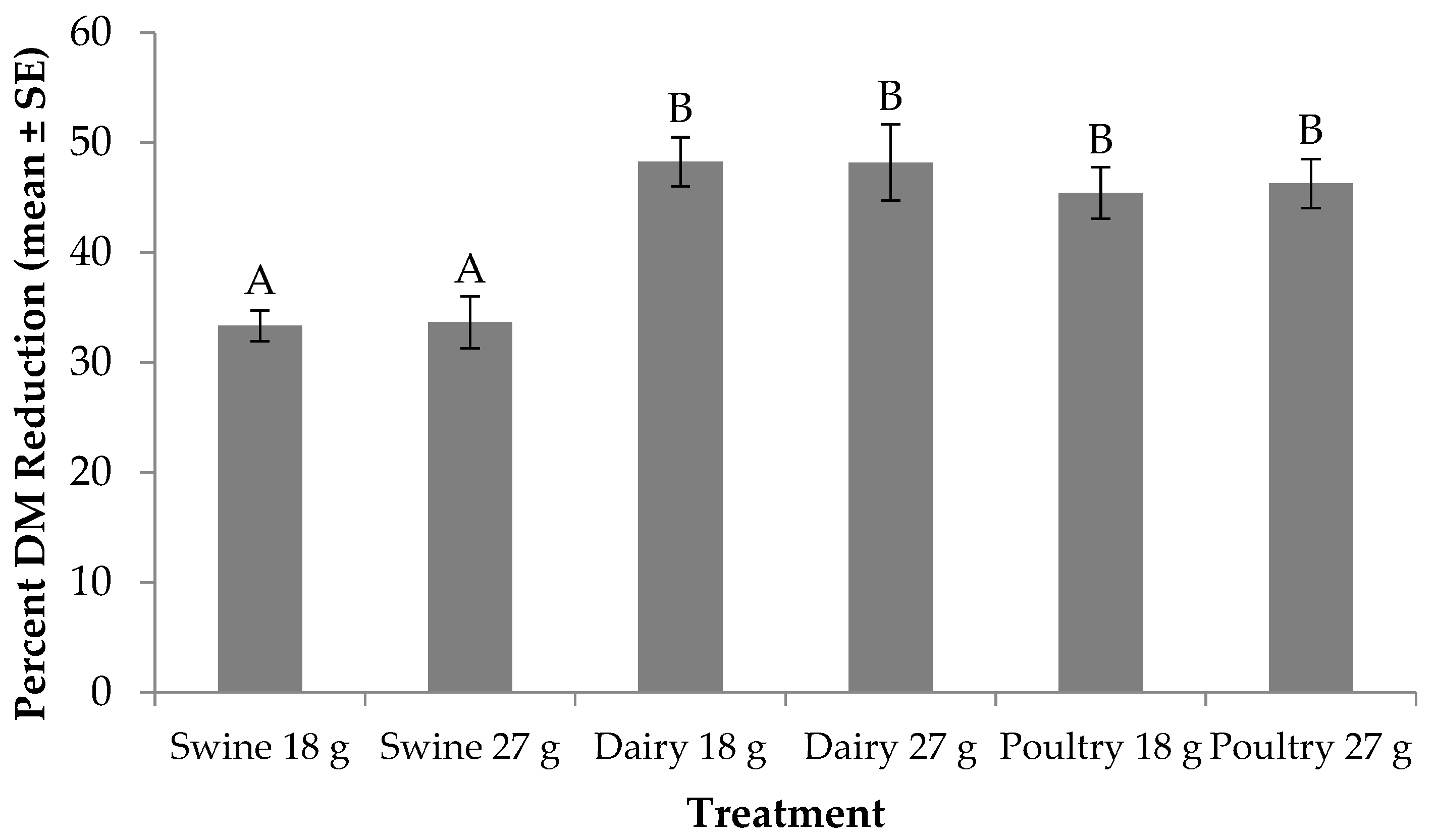
3.3. Percent DM reduction.
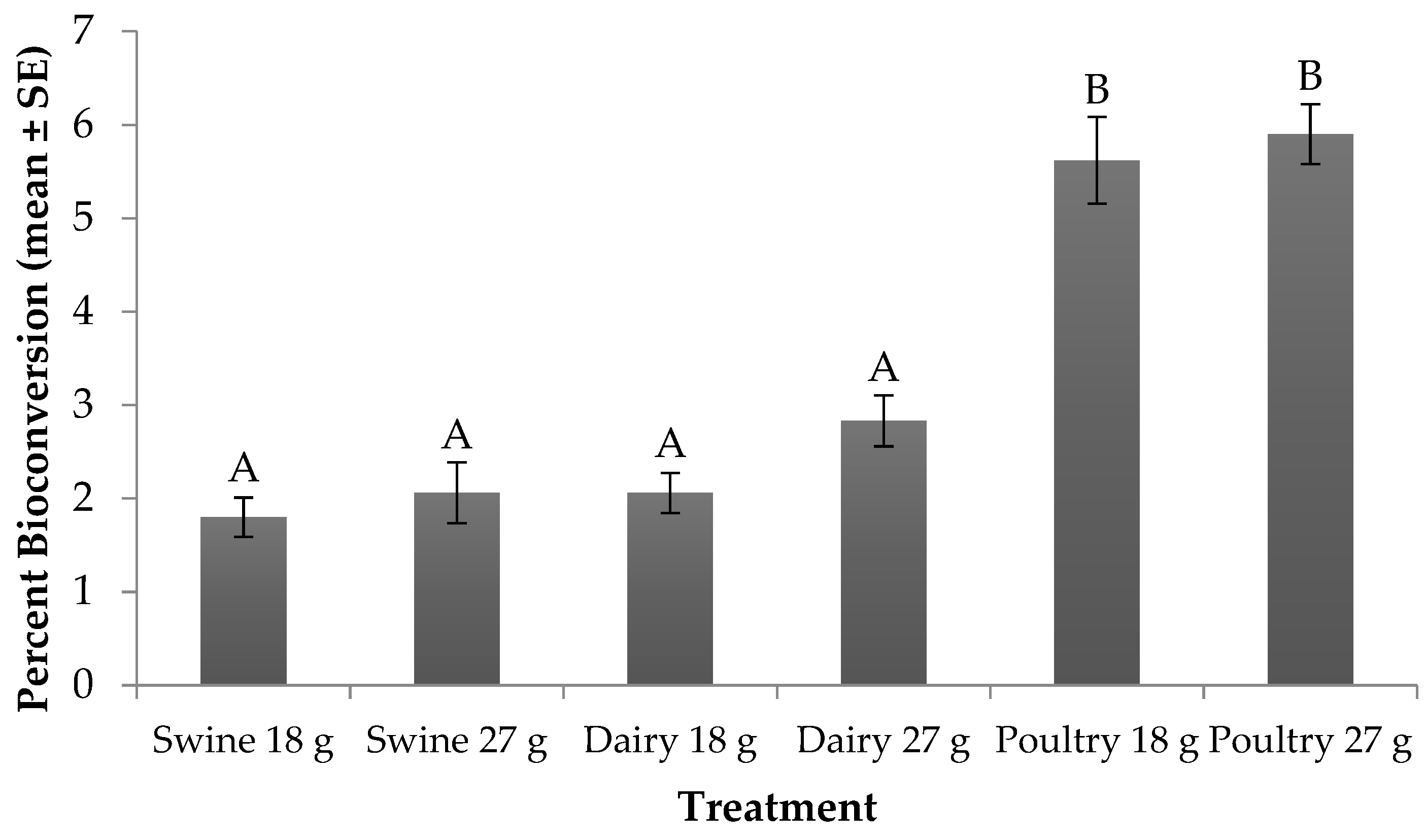
3.4. Percent Bioconversion
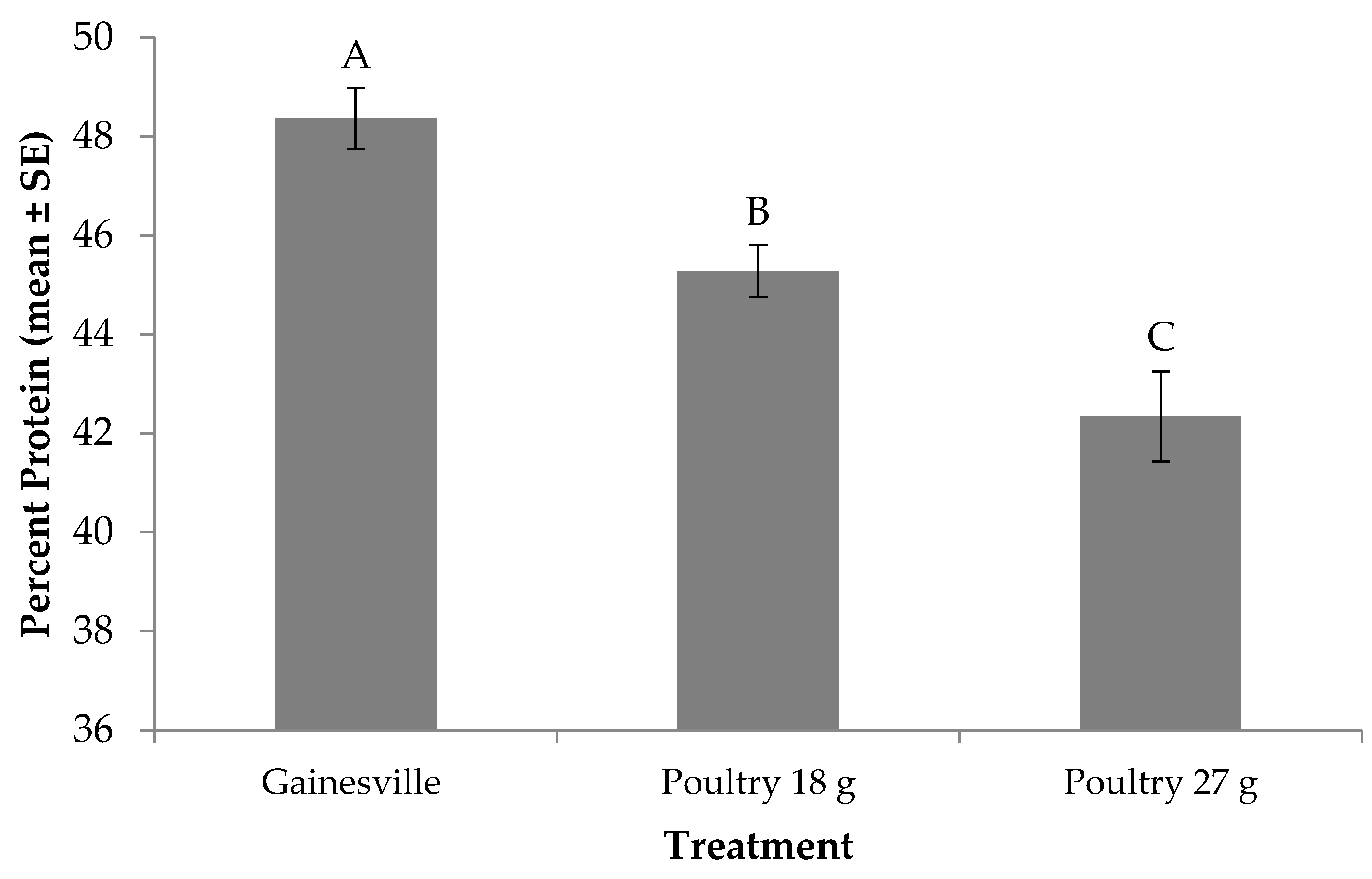
3.5. Protein Content of Prepupae
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Boland, M.J.; Rae, A.N.; Vereijken, J.M.; Meuwissen, M.P.; Fischer, A.R.; van Boekel, M.A.; Rutherfurd, S.M.; Gruppen, H.; Moughan, P.J.; Hendriks, W.H. The future supply of animal-derived protein for human consumption. Trends Food Sci. Technol. 2013, 29, 62–73. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Tran, G.; Henze, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- The United States Department of Agriculture (USDA). Overview of the United States Dairy Industry: Large Operations Increase Share of Inventory and Milk Production; USDA: Washington, DC, USA, 2010.
- The United States Department of Agriculture (USDA). Overview of the United States Hog Industry: Breeding Herd Efficiency Continues to Increase; USDA: Washington, DC, USA, 2015.
- Fine Woodworking (FWW). Factory Farmed Nation: 2015 Edition; Taunton Press: Newtown, CT, USA, 2015. [Google Scholar]
- Pfost, D.L.; Fulhage, C.D.; Rastorfer, D. Anaerobic Lagoons for Storage/Treatment of Livestock Manure; University of Missouri-Extension: Columbia, MO, USA, 2000. [Google Scholar]
- Myers, H.M.; Tomberlin, J.K.; Lambert, B.D.; Kattes, D. Development of black soldier fly (Diptera: Stratiomyidae) larvae fed dairy manure. Environ. Entomol. 2014, 37, 11–15. [Google Scholar] [CrossRef]
- Sheppard, D.C.; Newton, G.L.; Thompson, S.A.; Savage, S. A value-added manure management-system using the black soldier fly. Bioresour. Technol. 1994, 50, 275–279. [Google Scholar] [CrossRef]
- Oonincx, D.; van Huis, A.; van Loon, J. Nutrient utilisation by black soldier flies fed with chicken, pig, or cow manure. J. Insects Food Feed 2015, 1, 131–139. [Google Scholar] [CrossRef]
- ur Rehman, K.; Cai, M.; Xiao, X.; Zheng, L.; Wang, H.; Soomro, A.A.; Zhou, Y.; Li, W.; Yu, Z.; Zhang, J. Cellulose decomposition and larval biomass production from the co-digestion of dairy manure and chicken manure by mini-livestock (Hermetia illucens L.). J. Environ. Manag. 2017, 196, 458–465. [Google Scholar] [CrossRef]
- James, M. The genus Hermetia in the United States (Diptera: Stratiomyidae). Bull. Brooklyn Entomol. Soc. 1935, 30, 165–170. [Google Scholar]
- Martínez-Sánchez, A.; Magana, C.; Salona, M.; Rojo, S. First record of Hermetia illucens (Diptera: Stratiomyidae) on human corpses in Iberian Peninsula. Forensic Sci. Int. 2011, 206, e76–e78. [Google Scholar] [CrossRef]
- Tsagkarakis, A.E.; Arapostathi, E.; Strouvalis, G. First record of the black soldier fly, Hermetia illucens, in Greece. Entomol. Hell. 2015, 24, 27–30. [Google Scholar] [CrossRef]
- Callan, E. Hermetia illucens (L.)(Diptera, Stratiomyidae), a cosmopolitan American species long established in Australia and New Zealand. Entomol. Mon. Mag. 1974, 109, 232–234. [Google Scholar]
- Sheppard, C. Housefly and lesser fly control utilizing the black soldier fly in manure management systems for caged laying hens. Environ. Entomol. 1983, 12, 1439–1442. [Google Scholar] [CrossRef]
- Zhou, F.; Tomberlin, J.K.; Zheng, L.; Yu, Z.; Zhang, J. Developmental and waste reduction plasticity of three black soldier fly strains (Diptera: Stratiomyidae) raised on different livestock manures. J. Med. Entomol. 2013, 50, 1224–1230. [Google Scholar] [CrossRef]
- Wang, Y.-S.; Shelomi, M. Review of black soldier fly (Hermetia illucens) as animal feed and human food. Foods 2017, 6, 91. [Google Scholar] [CrossRef]
- Bondari, K.; Sheppard, D. Soldier fly larvae as feed in commercial fish production. Aquaculture 1981, 24, 103–109. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Sheppard, C.; Tomberlin, J.K.; Irving, S.; Newton, L.; McGuire, M.A.; Mosley, E.E.; Hardy, R.W.; Sealey, W. Fly prepupae as a feedstuff for rainbow trout, Oncorhynchus mykiss. J. World Aquac. Soc. 2007, 38, 59–67. [Google Scholar] [CrossRef]
- Sealey, W.M.; Gaylord, T.G.; Barrows, F.T.; Tomberlin, J.K.; McGuire, M.A.; Ross, C.; St-Hilaire, S. Sensory analysis of rainbow trout, Oncorhynchus mykiss, fed enriched black soldier fly prepupae, Hermetia illucens. J. World Aquac. Soc. 2011, 42, 34–45. [Google Scholar] [CrossRef]
- Kroeckel, S.; Harjes, A.-G.; Roth, I.; Katz, H.; Wuertz, S.; Susenbeth, A.; Schulz, C. When a turbot catches a fly: Evaluation of a pre-pupae meal of the black soldier fly (Hermetia illucens) as fish meal substitute—Growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 2012, 364, 345–352. [Google Scholar] [CrossRef]
- Hale, O.M. Dried Hermetia illucens larvae (Diptera: Stratiomyidae) as a feed additive for poultry. J. Ga. Entomol. Soc. 1973, 8, 16–20. [Google Scholar]
- Newton, G.L.; Booram, C.V.; Barker, R.W.; Hale, O.M. Dried Hermetia illucens larvae meal as a supplement for swine. J. Anim. Sci. 1977, 44, 395–400. [Google Scholar] [CrossRef]
- Liu, Q.; Tomberlin, J.K.; Brady, J.A.; Sanford, M.R.; Yu, Z. Black soldier fly (Diptera: Stratiomyidae) larvae reduce Escherichia coli in dairy manure. Environ. Entomol. 2008, 37, 1525–1530. [Google Scholar] [CrossRef]
- Lalander, C.H.; Fidjeland, J.; Diener, S.; Eriksson, S.; Vinnerås, B. High waste-to-biomass conversion and efficient Salmonella spp. reduction using black soldier fly for waste recycling. Agron. Sustain. Dev. 2015, 35, 261–271. [Google Scholar] [CrossRef]
- Beskin, K.V.; Holcomb, C.D.; Cammack, J.A.; Crippen, T.L.; Knap, A.H.; Sweet, S.T.; Tomberlin, J.K. Larval digestion of different manure types by the black soldier fly (Diptera: Stratiomyidae) impacts associated volatile emissions. Waste Manag. 2018, 74, 213–220. [Google Scholar] [CrossRef]
- Zheng, L.; Li, Q.; Zhang, J.; Yu, Z. Double the biodiesel yield: Rearing black soldier fly larvae, Hermetia illucens, on solid residual fraction of restaurant waste after grease extraction for biodiesel production. Renew. Energy 2012, 41, 75–79. [Google Scholar] [CrossRef]
- Čičková, H.; Newton, G.L.; Lacy, R.C.; Kozanek, M. The use of fly larvae for organic waste treatment. Waste Manag. 2015, 35, 68–80. [Google Scholar] [CrossRef]
- Charlton, A.; Dickinson, M.; Wakefield, M.; Fitches, E.; Kenis, M.; Han, R.; Zhu, F.; Kone, N.; Grant, M.; Devic, E. Exploring the chemical safety of fly larvae as a source of protein for animal feed. J. Insects Food Feed 2015, 1, 7–16. [Google Scholar] [CrossRef]
- Sheppard, D.C.; Tomberlin, J.K.; Joyce, J.A.; Kiser, B.C.; Sumner, S.M. Rearing methods for the black soldier fly (Diptera: Stratiomyidae). J. Med. Entomol. 2002, 39, 695–698. [Google Scholar] [CrossRef]
- Franson, M.A.H. Total Solids Dried at 103-105C: Standard Methods for the Examination of Water and Wastewater; American Public Health Association; American Water Works Association; Water Environment Federation: Washington, DC, USA, 1989. [Google Scholar]
- Hogsette, J.A. New diets for production of house flies and stable flies (Diptera, Muscidae) in the laboratory. J. Econ. Entomol. 1992, 85, 2291–2294. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Sheppard, D.C.; Joyce, J.A. Selected life-history traits of black soldier flies (Diptera: Stratiomyidae) reared on three artificial diets. Ann. Entomol. Soc. Am. 2002, 95, 379–386. [Google Scholar] [CrossRef]
- Yu, G.; Cheng, P.; Chen, Y.; Li, Y.; Yang, Z.; Chen, Y.; Tomberlin, J.K. Inoculating poultry manure with companion bacteria influences growth and development of black soldier fly (Diptera: Stratiomyidae) larvae. Environ. Entomol. 2011, 40, 30–35. [Google Scholar] [CrossRef]
- Ishii, S.; Yan, T.; Vu, H.; Hansen, D.L.; Hicks, R.E.; Sadowsky, M.J. Factors controlling long-term survival and growth of naturalized Escherichia coli populations in temperate field soils. Microbes Environ. 2009, 0912220144. [Google Scholar] [CrossRef]
- Cammack, J.A.; Tomberlin, J.K. The impact of diet protein and carbohydrate on select life-history traits of the black soldier fly Hermetia illucens (L.)(Diptera: Stratiomyidae). Insects 2017, 8, 56. [Google Scholar] [CrossRef]
- Gobbi, P.; Martínez-Sánchez, A.; Rojo, S. The effects of larval diet on adult life-history traits of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Eur. J. Entomol. 2013, 110, 461. [Google Scholar] [CrossRef]
- Nguyen, T.T.X.; Tomberlin, J.K.; Vanlaerhoven, S. Influence of Resources on Hermetia illucens (Diptera: Stratiomyidae) Larval Development. J. Med. Entomol. 2013, 50, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Tomberlin, J.K.; Vanlaerhoven, S. Ability of black soldier fly (Diptera: Stratiomyidae) larvae to recycle food waste. Environ. Entomol. 2015, 44, 406–410. [Google Scholar] [CrossRef]
- Martin, J.H.; Loehr, R.C.; Pilbeam, T.E. Animal manures as feedstuffs: Nutrient characteristics. Agric. Wastes 1983, 6, 131–166. [Google Scholar] [CrossRef]
- Chen, S.; Liao, W.; Liu, C.; Wen, Z.; Kincaid, R.L.; Harrison, J.H.; Elliott, D.C.; Brown, M.D.; Solana, A.E.; Stevens, D.J. Value-Added Chemicals from Animal Manure; Pacific Northwest National Laboratory: Richland, WA, USA, 2003.
- Cheng, J.Y.; Chiu, S.L.; Lo, I.M. Effects of moisture content of food waste on residue separation, larval growth and larval survival in black soldier fly bioconversion. Waste Manag. 2017, 67, 315–323. [Google Scholar] [CrossRef]
- Fletcher, M.; Axtell, R.; Stinner, R. Longevity and fecundity of Musca domestica (Diptera: Muscidae) as a function of temperature. J. Med. Entomol. 1990, 27, 922–926. [Google Scholar] [CrossRef]
- Banks, I.J.; Gibson, W.T.; Cameron, M.M. Growth rates of black soldier fly larvae fed on fresh human faeces and their implication for improving sanitation. Trop. Med. Int. Health 2014, 19, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Meneguz, M.; Gasco, L.; Tomberlin, J.K. Impact of pH and feeding system on black soldier fly (Hermetia illucens, L.; Diptera: Stratiomyidae) larval development. PLoS ONE 2018, 13, e0202591. [Google Scholar] [CrossRef]
- ur Rehman, K.; Rehman, A.; Cai, M.; Zheng, L.; Xiao, X.; Somroo, A.A.; Wang, H.; Li, W.; Yu, Z.; Zhang, J. Conversion of mixtures of dairy manure and soybean curd residue by black soldier fly larvae (Hermetia illucens L.). J. Clean. Prod. 2017, 154, 366–373. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Cranfill, K.; McGuire, M.A.; Mosley, E.E.; Tomberlin, J.K.; Newton, L.; Sealey, W.; Sheppard, C.; Irving, S. Fish offal recycling by the black soldier fly produces a foodstuff high in omega-3 fatty acids. J. World Aquac. Soc. 2007, 38, 309–313. [Google Scholar] [CrossRef]
- Furman, D.P.; Young, R.D.; Catts, E.P. Hermetia illucens (Linnaeus) as a factor in the natural control of Musca domestica Linnaeus. J. Econ. Entomol. 1959, 52, 917–921. [Google Scholar] [CrossRef]
- Lalander, C.; Diener, S.; Zurbrügg, C.; Vinnerås, B. Effects of feedstock on larval development and process efficiency in waste treatment with black soldier fly (Hermetia illucens). J. Clean. Prod. 2019, 208, 211–219. [Google Scholar] [CrossRef]
- Pimentel, A.C.; Montali, A.; Bruno, D.; Tettamanti, G. Metabolic adjustment of the larval fat body in Hermetia illucens to dietary conditions. J. Asia-Pac. Entomol. 2017, 20, 1307–1313. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| % | Swine | Dairy | Poultry |
|---|---|---|---|
| N | 2.10 ± 0.02 A | 2.10 ± 0.03 A | 2.40 ± 0.04 B |
| P | 2.42 ± 0.08 A | 0.66 ± 0.11 B | 2.43 ± 0.05 A |
| K | 0.01 ± 0.02 A | 0.57 ± 0.00 B | 2.44 ± 0.01 C |
| Ca | 4.36 ± 0.05 A | 1.96 ± 0.19 B | 14.0 ± 0.47 C |
| Mg | 0.87 ± 0.03 A | 0.67 ± 0.00 B | 0.56 ± 0.00 B |
| Na | 0.55 ± 0.01 A | 0.32 ± 0.02 B | 0.58 ± 0.00 A |
| Zn | 0.08 ± 0.00 A | 0.02 ± 0.00 B | 0.04 ± 0.00 C |
| Fe | 0.01 ± 0.00 A | 0.27 ± 0.04 B | 0.19 ± 0.02 B |
| Cu | 0.01 ± 0.00 A | 0.00 ± 0.00 B | 0.00 ± 0.00 B |
| Mn | 0.05 ± 0.00 A | 0.02± 0.00 B | 0.05 ± 0.00 A |
| S | 0.78 ± 0.01 A | 0.41 ± 0.02 B | 0.86 ± 0.00 C |
| B | 0.08 ± 0.00 A | 0.00 ± 0.00 B | 0.00 ± 0.00 B |
| Manure Type | Feed Amount (g/2 days) | Initial (%) |
|---|---|---|
| Swine | 18 | 73.94 ± 0.41 A |
| 27 | ||
| Dairy | 18 | 83.92 ± 0.63 B |
| 27 | ||
| Poultry | 18 | 77.24 ± 0.29 C |
| 27 | ||
| Gainesville | 27 | 70.00 ± 0.56 D |
| Treatment | Final Larval Weight (g) | Time to First Prepupation (d) | Prepupal Weight (g) | Percent Prepupation | Adult Male Weight (g) | Adult Male Longevity (d) | Adult Female Weight (g) | Adult Female Longevity (d) | |
|---|---|---|---|---|---|---|---|---|---|
| Swine | 18 | 0.1264 ± 0.0063 A | 20.3 ± 1.1 A | 0.0678 ± 0.0029 A | 69.5 ± 5. 3 A | 0.0273 ± 0.0016 A | 4.9 ± 0.3 A | 0.0320 ± 0.0018A | 4.7 ± 0.1 A |
| 27 | 0.1189 ± 0.0132 A | 16.7 ± 0.6 B,C | 0.0827 ± 0.0009 A,B | 83.8 ± 6.0 A,B,C | 0.0321 ± 0.0019 B | 5.5 ± 0.2 A | 0.0405 ± 0.0028 B,C | 5.1 ± 0.2 A,B | |
| Dairy | 18 | 0.1306 ± 0.0093 A | 18.6 ± 0.7 A,B | 0.0668 ± 0.0016 A | 76.7 ± 3.4 A,B | 0.0271 ± 0.0004 A | 5.1 ± 0.2 A | 0.0330 ± 0.0009 B,C | 5.9 ± 0.2 A |
| 27 | 0.1339 ± 0.0118 A | 15.6 ± 0.6 C | 0.0990 ± 0.0054 B,C | 92.7 ± 3.1 B,C | 0.0334 ± 0.0017 B,C | 5.7 ± 0.4 A | 0.0427 ± 0.0023 B | 6.0 ± 0.2 B,C | |
| Poultry | 18 | 0.1419 ± 0.0089 A | 11.6 ± 0.2 D | 0.0866 ± 0.0016 A,B | 93.7 ± 2.7 B,C | 0.0371 ± 0.0012 C | 7.2 ± 0.2 B | 0.0420 ± 0.0019 B,C | 6.5 ± 0.2 C,D |
| 27 | 0.1541 ± 0.0168 A | 11.3 ± 0.3 D,E | 0.1137 ± 0.0085 C,D | 95.3 ± 1.7 C | 0.0483 ± 0.0022 D | 8.2 ± 0.3 C | 0.0598 ± 0.0044 C | 7.2 ± 0.3 D | |
| Gainesville 2 | 27 | 0.2019 ± 0.0020 B | 9.0 ± 0.0 E | 0.1324 ± 0.0008 D | 86.0 ± 2.9 A,B,C | 0.0516 ± 0.0014 D | 8.1 ± 0.1 B,C | 0.0653 ± 0.0014 C | 6.7 ± 0.3 C,D |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miranda, C.D.; Cammack, J.A.; Tomberlin, J.K. Life-History Traits of the Black Soldier Fly, Hermetia illucens (L.) (Diptera: Stratiomyidae), Reared on Three Manure Types. Animals 2019, 9, 281. https://doi.org/10.3390/ani9050281
Miranda CD, Cammack JA, Tomberlin JK. Life-History Traits of the Black Soldier Fly, Hermetia illucens (L.) (Diptera: Stratiomyidae), Reared on Three Manure Types. Animals. 2019; 9(5):281. https://doi.org/10.3390/ani9050281
Chicago/Turabian StyleMiranda, Chelsea D., Jonathan A. Cammack, and Jeffery K. Tomberlin. 2019. "Life-History Traits of the Black Soldier Fly, Hermetia illucens (L.) (Diptera: Stratiomyidae), Reared on Three Manure Types" Animals 9, no. 5: 281. https://doi.org/10.3390/ani9050281
APA StyleMiranda, C. D., Cammack, J. A., & Tomberlin, J. K. (2019). Life-History Traits of the Black Soldier Fly, Hermetia illucens (L.) (Diptera: Stratiomyidae), Reared on Three Manure Types. Animals, 9(5), 281. https://doi.org/10.3390/ani9050281