Food Preferences in Cats: Effect of Dietary Composition and Intrinsic Variables on Diet Selection

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Procedures to Determine Food Preferences and Database
2.3. Statistical Analysis
2.3.1. Effect of Nutrient Composition over Cats’ Food Preferences
2.3.2. Effect of Intrinsic Variables Over Cats’ Food Preferences
3. Results
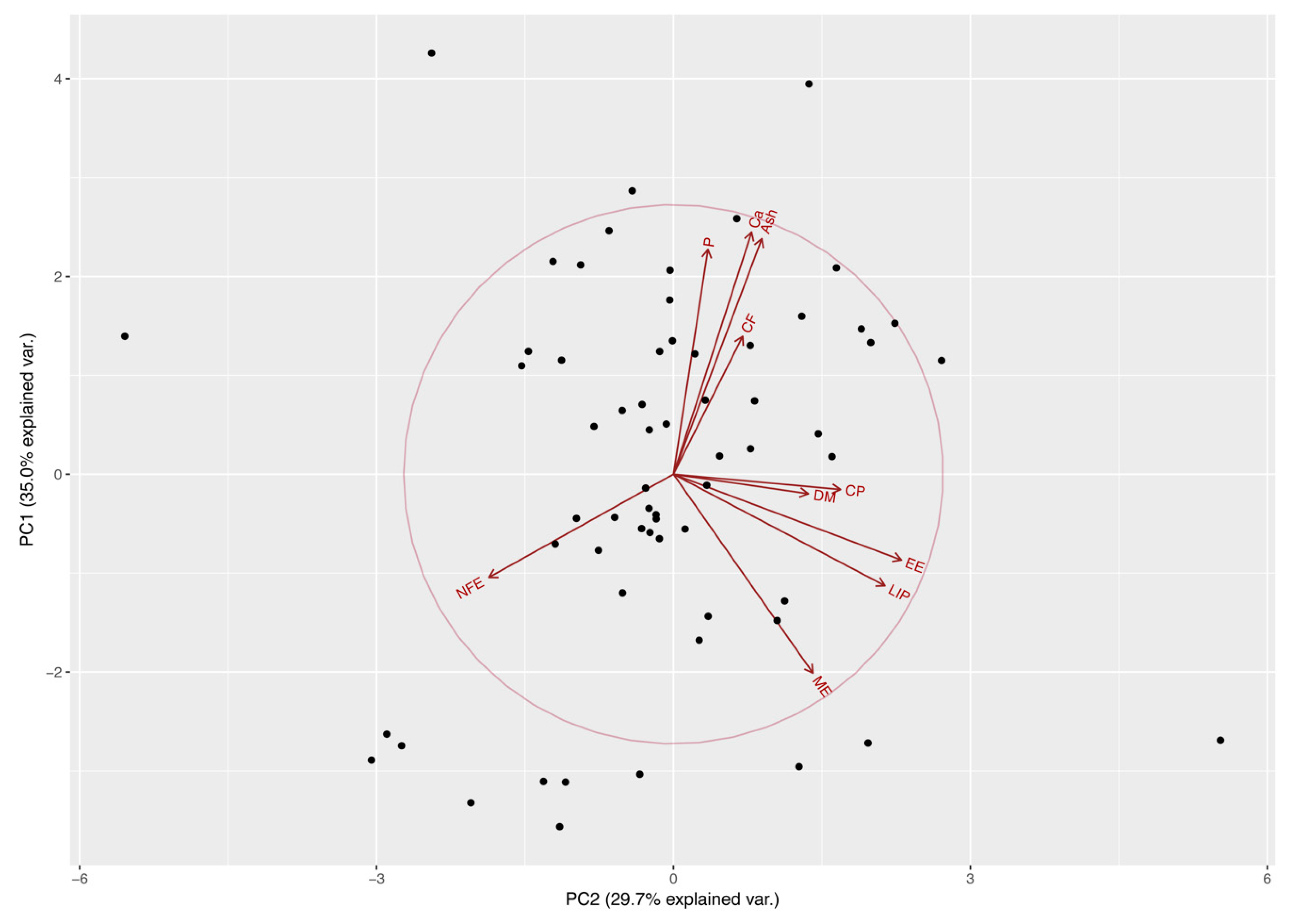
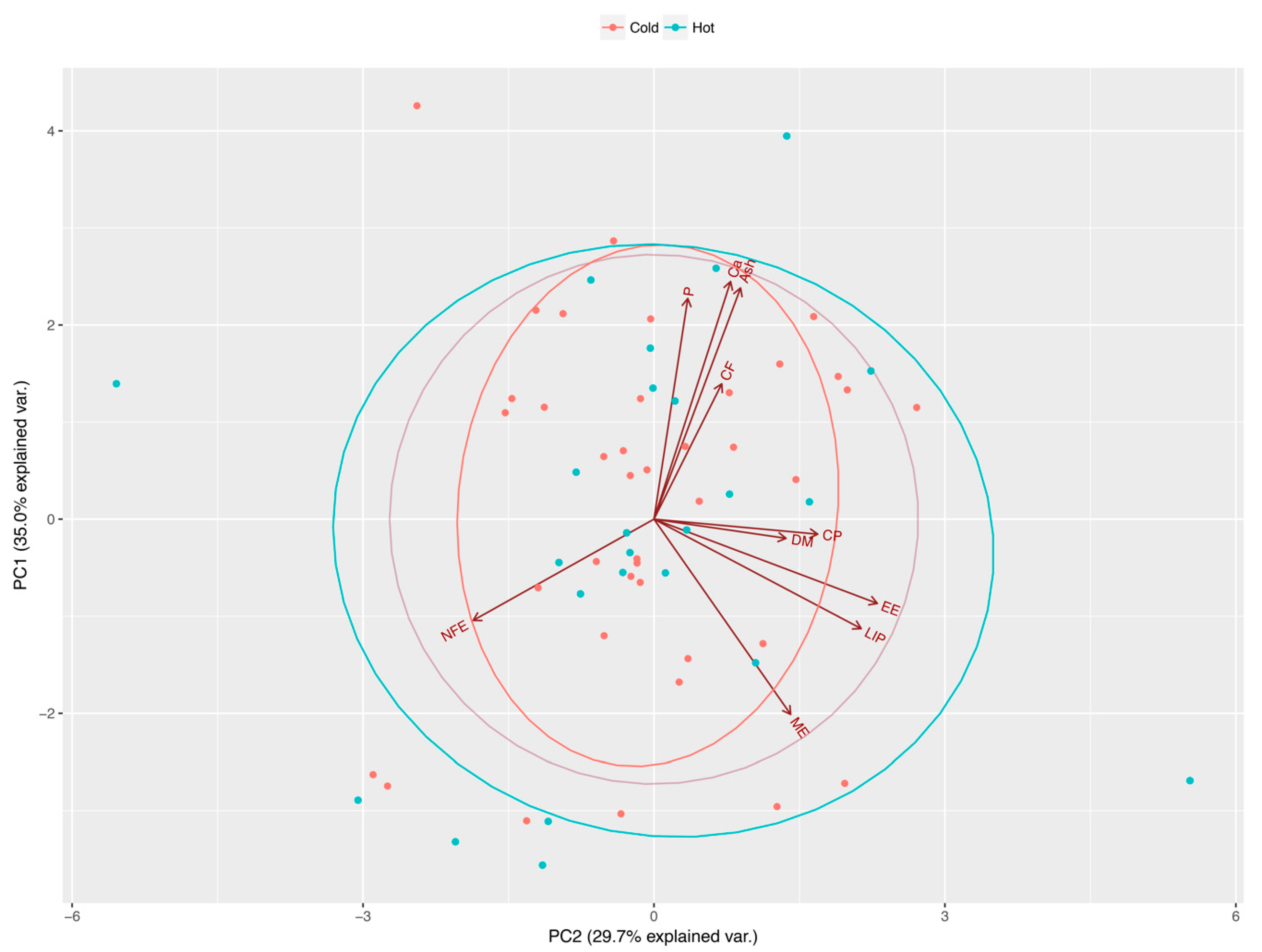
3.1. Effect of Nutrient Composition on Cats’ Food Preferences
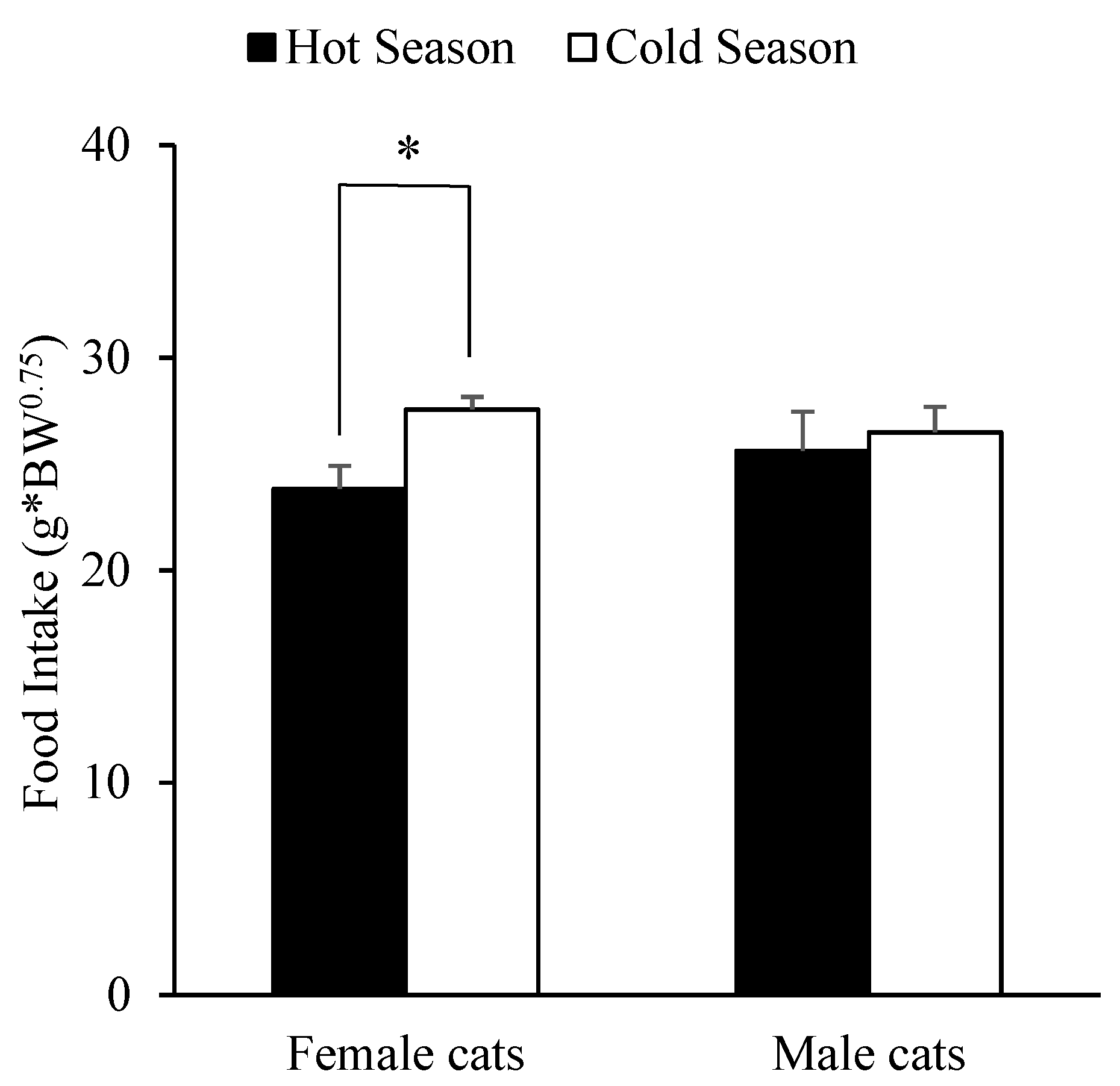
3.2. Effect of Sex, Breed, Age, Body Weight and Season on Food Intake by Cats
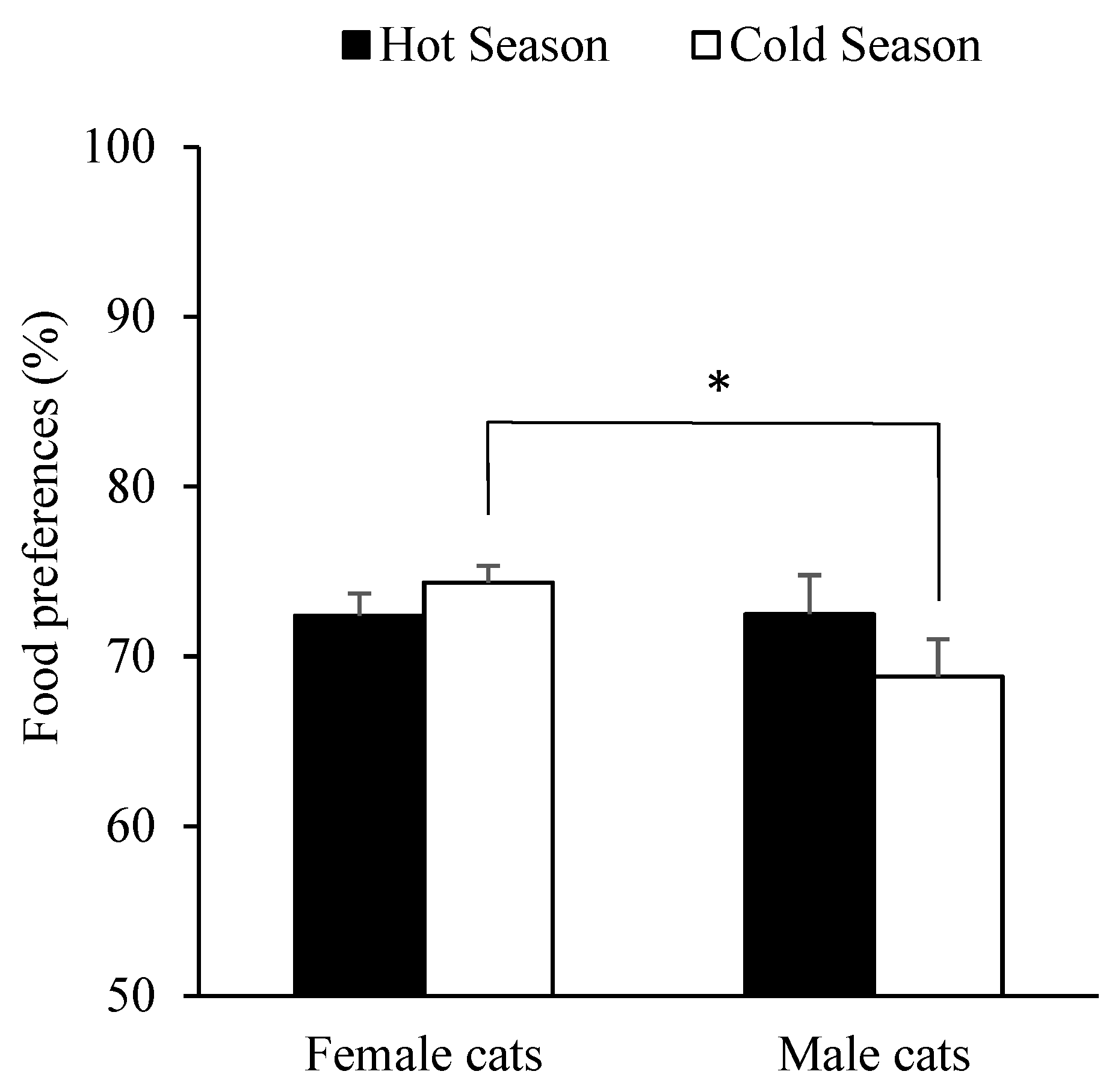
3.3. Effect of Sex, Breed, Age, Body Weight, and Season on Food Preferences in Cats
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- MacDonald, M.L.; Rogers, Q.R.; Morris, J.G. Nutrition of the Domestic Cat, a Mammalian Carnivore. Annu. Rev. Nutr. 1984, 4, 521–562. [Google Scholar] [CrossRef] [PubMed]
- Legrand-Defretin, V. Differences between cats and dogs: A nutritional view. Proc. Nutr. Soc. 1994, 53, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Tobie, C.; Péron, F.; Larose, C. Assessing Food Preferences in Dogs and Cats: A Review of the Current Methods. Animals 2015, 5, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Peachey, S.E.; Harper, E.J. Aging Does Not Influence Feeding Behavior in Cats. J. Nutr. 2002, 132, 1735S–1739S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradshaw, J.W.S. The Evolutionary Basis for the Feeding Behavior of Domestic Dogs (Canis familiaris) and Cats (Felis catus). J. Nutr. 2006, 136, 1927S–1931S. [Google Scholar] [CrossRef] [PubMed]
- Zoran, D.L.; Buffington, C.A. Effects of nutrition choices and lifestyle changes on the well-being of cats, a carnivore that has moved indoors. J. Am. Vet. Med Assoc. 2011, 239, 596–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, E.; Rogers, Q.R.; Morris, J.G. Feeding behavior of the cat fed laboratory and commercial diets. Nutr. Res. 1981, 1, 499–507. [Google Scholar] [CrossRef]
- Bradshaw, J.W.S.; Thorne, C.J. Feeding behaviour. In The Waltham Book of Dog and Cat Behaviour; Thorne, C.J., Ed.; Pergamon Press: Oxford, UK, 1992; pp. 118–129. [Google Scholar]
- Hullar, I.; Fekete, S.; Andrasofszky, E.; Szocs, Z.; Berkenyi, T. Factors influencing the food preference of cats. J. Anim. Physiol. Anim. Nutr. (Berl.) 2001, 85, 205–211. [Google Scholar] [CrossRef]
- Hewson-Hughes, A.K.; Hewson-Hughes, V.L.; Colyer, A.; Miller, A.T.; Hall, S.R.; Raubenheimer, D.; Simpson, S.J. Consistent proportional macronutrient intake selected by adult domestic cats (Felis catus) despite variations in macronutrient and moisture content of foods offered. J. Comp. Physiol. B 2013, 183, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Salaun, F.; Blanchard, G.; Le Paih, L.; Roberti, F.; Niceron, C. Impact of macronutrient composition and palatability in wet diets on food selection in cats. J. Anim. Physiol. Anim. Nutr. 2017, 101, 320–328. [Google Scholar] [CrossRef]
- Beaver, B.V. Feline Ingestive Behavior. In Feline Behavior, 2nd ed.; Beaver, B.V., Ed.; W.B. Saunders: St. Louis, MO, USA, 2003; Chapter 7; pp. 212–246. [Google Scholar]
- Bradshaw, J.W. Mere exposure reduces cats’ neophobia to unfamiliar food. Anim. Behav. 1986, 34, 613–614. [Google Scholar] [CrossRef]
- Church, S.C.; Allen, J.A.; Bradshaw, J.W.S. Anti-apostatic food selection by the domestic cat. Anim. Behav. 1994, 48, 747–749. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, J.W.S.; Healey, L.M.; Thorne, C.J.; Macdonald, D.W.; Arden-Clark, C. Differences in food preferences between individuals and populations of domestic cats Felis silvestris catus. Appl. Anim. Behav. Sci. 2000, 68, 257–268. [Google Scholar] [CrossRef]
- Castonguay, T.W. Dietary dilution and intake in the cat. Physiol. Behav. 1981, 27, 547–549. [Google Scholar] [CrossRef]
- Kane, E.; Morris, J.G.; Rogers, Q.R. Acceptability and digestibility by adult cats of diets made with various sources and levels of fat. J. Anim. Sci. 1981, 53, 1516–1523. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ruiz, N.R.; López-Díaz, J.A.; Wall-Medrano, A.; Jiménez-Castro, J.A.; Angulo, O. Oral fat perception is related with body mass index, preference and consumption of high-fat foods. Physiol. Behav. 2014, 129, 36–42. [Google Scholar] [CrossRef]
- Inselman-Temkin, B.R.; Flynn, J.P. Sex-dependent effects of gonadal and gonadotropic hormones on centrally-elicited attack in cats. Brain Res. 1973, 60, 393–410. [Google Scholar] [CrossRef]
- Caine-Bish, N.L.; Scheule, B. Gender Differences in Food Preferences of School-Aged Children and Adolescents. J. Sch. Health 2009, 79, 532–540. [Google Scholar] [CrossRef]
- Rolls, E.T.; Kellerhals, M.B.; Nichols, T.E. Age differences in the brain mechanisms of good taste. NeuroImage 2015, 113, 298–309. [Google Scholar] [CrossRef]
- Horwitz, D.; Soulard, Y.; Junien-Castagna, A. Comportamiento alimentario del gato. In Enciclopedia de la Nutrición Clínica Felina; Pibot, P., Biourge, V., Elliott, D., Eds.; Royal Canine: Nueva York, Estados Unidos, 2010; pp. 439–478. [Google Scholar]
- DMC. Información Meteológica. Available online: http://www.meteochile.cl/PortalDMC-web/index.xhtml (accessed on 4 July 2018).
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2005. [Google Scholar]
- R_Core_Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Dohoo, R.; Martin, W.; Stryhn, H. Veterinary Epidemiologic Research, 2nd ed.; VER Inc.: Charlottetown, PE, Canada, 2009; p. 865. [Google Scholar]
- Pinheiro, J.; Bates, D. Mixed-Effects Models in S and S-Plus; Springer: New York, NY, USA, 2000. [Google Scholar]
- Thrusfield, M.; Christley, R. Veterinary Epidemiology, 4th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2018; p. 888. [Google Scholar]
- Heinze, G.; Wallisch, C.; Dunkler, D. Variable selection—A review and recommendations for the practicing statistician. Biom. J. 2018, 60, 431–449. [Google Scholar] [CrossRef]
- Di Rienzo, J.; Guzman, A.; Casanoves, F. A multiple comparisons method based on the distribution of the root node distance of a binary tree obtained by average linkage of the matrix of Euclidean distances between treatment means. J. Agric. Biol. Environ. Stat. 2002, 7, 129–142. [Google Scholar] [CrossRef]
- Dahiru, T. p-Value, a true test of statistical significance? A cautionary note. Ann. Ib. Postgrad. Med. 2008, 6, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, G.C.; Koppel, K. Pet Food Palatability Evaluation: A Review of Standard Assay Techniques and Interpretation of Results with a Primary Focus on Limitations. Animals 2015, 5, 43–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AAFCO. AAFCO Methods for Substaining Nutritional Adequacy of Dog and Cat Foods; Association of American Feed Control Officials: Champaign, IL, USA, 2013. [Google Scholar]
- Kerr, K.R. COMPANION ANIMALS SYMPOSIUM: Dietary management of feline lower urinary tract symptoms1,2. J. Anim. Sci. 2013, 91, 2965–2975. [Google Scholar] [CrossRef] [PubMed]
- BSAVA. Manual de Medicina Felina; Ediciones S: Barcelona, España, 2014; p. 726. [Google Scholar]
- Laflammme, D. Focus on Nutrition: Cats and carbohydrates: Implications for health and disease. Compendium (Yardley, PA) 2010, 32, E1–E3. [Google Scholar]
- Fekete, S.G.; Hullár, I.; Andrásofszky, E.; Kelemen, F. Effect of different fibre types on the digestibility of nutrients in cats. J. Anim. Physiol. Anim. Nutr. 2004, 88, 138–142. [Google Scholar] [CrossRef]
- Asarian, L.; Geary, N. Modulation of appetite by gonadal steroid hormones. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2006, 361, 1251–1263. [Google Scholar] [CrossRef] [Green Version]
- Park, H.-J.; Lee, S.-E.; Oh, J.-H.; Seo, K.-W.; Song, K.-H. Leptin, adiponectin and serotonin levels in lean and obese dogs. BMC Vet. Res. 2014, 10, 113. [Google Scholar] [CrossRef]
- Bell, D.D.; Zucker, I. Sex differences in body weight and eating: Organization and activation by gonadal hormones in the rat. Physiol. Behav. 1971, 7, 27–34. [Google Scholar] [CrossRef]
- Vester, B.M.; Belsito, K.R.; Swanson, K.S.; Keel, T.; Graves, T.K. Impact of ovariohysterectomy and food intake on body composition, physical activity, and adipose gene expression in cats1. J. Anim. Sci. 2009, 87, 594–602. [Google Scholar] [CrossRef] [Green Version]
- Lenis Sanin, Y.; Zuluaga Cabrera, A.M.; Tarazona Morales, A.M. Adaptive Responses to Thermal Stress in Mammals. Rev. Med. Vet. 2016, 31, 121–135. [Google Scholar]
- Collin, A.; van Milgen, J.; Dubois, S.; Noblet, J. Effect of high temperature on feeding behaviour and heat production in group-housed young pigs. Br. J. Nutr. 2001, 86, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Stella, J.L.; Croney, C.C. Environmental Aspects of Domestic Cat Care and Management: Implications for Cat Welfare. Sci. World J. 2016, 2016, 6296315. [Google Scholar] [CrossRef] [PubMed]
- Shimada, A. Nutrición Animal; Editorial Trillas: Ciudad de México, México, 2007; p. 388. [Google Scholar]
- Serisier, S.; Feugier, A.; Delmotte, S.; Biourge, V.; German, A.J. Seasonal Variation in the Voluntary Food Intake of Domesticated Cats (Felis Catus). PLoS ONE 2014, 9, e96071. [Google Scholar] [CrossRef] [PubMed]
- Stornelli, M. Particularidades fisiológicas de la reproducción en felinos. Rev. Bras. Reprod. Anim. 2007, 31, 71–76. [Google Scholar]
- Heisler, L.K.; Kanarek, R.B.; Homoleski, B. Reduction of Fat and Protein Intakes But Not Carbohydrate Intake Following Acute and Chronic Fluoxetine in Female Rats. Pharmacol. Biochem. Behav. 1999, 63, 377–385. [Google Scholar] [CrossRef]
- Parker, G.C.; Bishop, C.; Coscina, D.V. Estrous cycle and food availability affect feeding induced by amygdala 5-HT receptor blockade. Pharmacol. Biochem. Behav. 2002, 71, 701–707. [Google Scholar] [CrossRef]
- Bartoshuk, L.M.; Duffy, V.B.; Miller, I.J. PTC/PROP tasting: Anatomy, psychophysics, and sex effects. Physiol. Behav. 1994, 56, 1165–1171. [Google Scholar] [CrossRef]
- Haase, L.; Green, E.; Murphy, C. Males and females show differential brain activation to taste when hungry and sated in gustatory and reward areas. Appetite 2011, 57, 421–434. [Google Scholar] [CrossRef] [Green Version]
- Becques, A.; Serra, J.; Gouat, P.; Larose, C. Effects of Pre- and Postnatal Olfactogustatory Experience on Early Preferences at Birth and Dietary Selection at Weaning in Kittens. Chem. Senses 2009, 35, 41–45. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutritional Components 1 | Principal Component Eigenvectors | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | PC8 | PC9 | PC10 | |
| DM | −0.04 | 0.29 | 0.66 | 0.24 | 0.32 | −0.15 | −0.22 | 0.05 | −0.31 | 0.38 |
| CP | −0.03 | 0.35 | −0.38 | 0.51 | 0.49 | 0.12 | −0.08 | 0.04 | −0.08 | −0.43 |
| CF | 0.27 | 0.14 | 0.13 | −0.66 | 0.59 | 0.10 | 0.26 | 0.03 | 0.09 | −0.14 |
| EE | −0.17 | 0.49 | −0.01 | −0.25 | −0.38 | −0.24 | 0.25 | 0.11 | −0.55 | −0.30 |
| NFE | −0.20 | −0.39 | 0.52 | 0.04 | −0.02 | 0.11 | −0.11 | 0.19 | 0.00 | −0.68 |
| ASH | 0.47 | 0.19 | 0.15 | 0.03 | −0.15 | −0.39 | −0.28 | −0.57 | 0.21 | −0.31 |
| Ca | 0.48 | 0.16 | 0.06 | 0.16 | −0.19 | −0.17 | 0.06 | 0.74 | 0.31 | −0.01 |
| P | 0.45 | 0.07 | 0.19 | 0.22 | −0.23 | 0.69 | 0.30 | −0.20 | −0.22 | 0.01 |
| LIP | −0.22 | 0.45 | 0.03 | −0.27 | −0.20 | 0.46 | −0.57 | 0.08 | 0.29 | 0.00 |
| ME | −0.39 | 0.30 | 0.25 | 0.19 | −0.05 | −0.04 | 0.56 | −0.17 | 0.55 | 0.01 |
| Principal Component Eigenvalues | ||||||||||
| SD | 1.87 | 1.72 | 1.18 | 1.02 | 0.74 | 0.54 | 0.39 | 0.29 | 0.21 | 0.08 |
| % of Variance | 34.98 | 29.70 | 13.87 | 10.37 | 5.45 | 2.89 | 1.49 | 0.83 | 0.43 | 0.00 |
| Cumulative % | 34.98 | 64.68 | 78.55 | 88.92 | 94.37 | 97.25 | 98.74 | 99.57 | 99.99 | 100.0 |
| Variable | OR 1 | SE 2 | p-Value |
|---|---|---|---|
| (Intercept) | 74.3906 | 1.3737 | <2 × 10−16 |
| PC1 | −1.4203 | 0.7403 | 0.0598 |
| PC2 | −1.5687 | 0.8034 | 0.0555 |
| PC4 | 2.6764 | 1.3598 | 0.0537 |
| Variable | OR 1 | SE 2 | p-Value |
|---|---|---|---|
| (Intercept) | 46.90 | 5.21 | <0.001 |
| Weight | −5.49 | 0.82 | <0.001 |
| Sex | |||
| Male | 11.76 | 4.72 | 0.014 |
| Season | |||
| Autumn | 7.86 | 3.43 | 0.022 |
| Spring | −5.31 | 3.32 | 0.110 |
| Summer | 2.66 | 4.02 | 0.508 |
| Weight: Sex | |||
| Male | −2.19 | 1.05 | 0.038 |
| Weight: Season | |||
| Autumn | −1.68 | 0.99 | 0.089 |
| Spring | 0.52 | 0.86 | 0.548 |
| Summer | 0.50 | 1.11 | 0.652 |
| Sex: Season | |||
| Male: Autumn | 4.20 | 2.52 | 0.096 |
| Male: Spring | 0.97 | 2.40 | 0.686 |
| Male: Summer | 4.18 | 2.67 | 0.119 |
| Variable | OR 1 | SE 2 | p-Value |
|---|---|---|---|
| (Intercept) | 76.22 | 2.95 | <0.001 |
| Weight | −2.92 | 1.46 | 0.046 |
| Weight 2* | −0.29 | 0.65 | 0.661 |
| Weight 3* | 1.16 | 0.37 | 0.002 |
| Sex | |||
| Male | −8.42 | 3.60 | 0.023 |
| Season | |||
| Autumn | −1.59 | 2.31 | 0.493 |
| Spring | −0.76 | 2.25 | 0.736 |
| Summer | −10.03 | 2.72 | <0.001 |
| Sex: Season | |||
| Male: Autumn | 7.62 | 4.32 | 0.078 |
| Male: Spring | 7.78 | 4.16 | 0.062 |
| Male: Summer | 10.22 | 4.65 | 0.028 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alegría-Morán, R.A.; Guzmán-Pino, S.A.; Egaña, J.I.; Sotomayor, V.; Figueroa, J. Food Preferences in Cats: Effect of Dietary Composition and Intrinsic Variables on Diet Selection. Animals 2019, 9, 372. https://doi.org/10.3390/ani9060372
Alegría-Morán RA, Guzmán-Pino SA, Egaña JI, Sotomayor V, Figueroa J. Food Preferences in Cats: Effect of Dietary Composition and Intrinsic Variables on Diet Selection. Animals. 2019; 9(6):372. https://doi.org/10.3390/ani9060372
Chicago/Turabian StyleAlegría-Morán, Raúl A., Sergio A. Guzmán-Pino, Juan I. Egaña, Valeria Sotomayor, and Jaime Figueroa. 2019. "Food Preferences in Cats: Effect of Dietary Composition and Intrinsic Variables on Diet Selection" Animals 9, no. 6: 372. https://doi.org/10.3390/ani9060372
APA StyleAlegría-Morán, R. A., Guzmán-Pino, S. A., Egaña, J. I., Sotomayor, V., & Figueroa, J. (2019). Food Preferences in Cats: Effect of Dietary Composition and Intrinsic Variables on Diet Selection. Animals, 9(6), 372. https://doi.org/10.3390/ani9060372