Copy Number Variation of the SHE Gene in Sheep and Its Association with Economic Traits

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Welfare
2.2. Animals and Growth Traits Measurements
2.3. Preparation of Sample and Genomic DNA
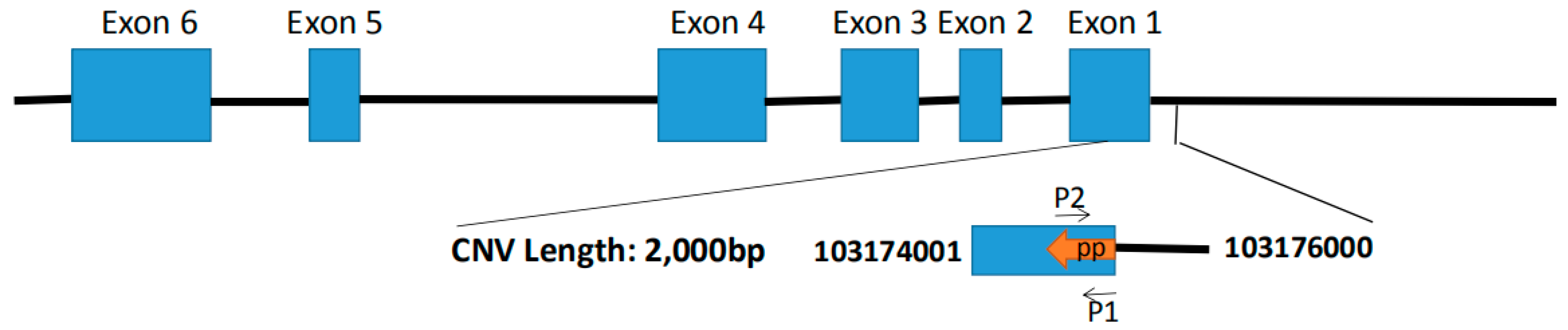
2.4. Candidate Gene Identification and Primer Design
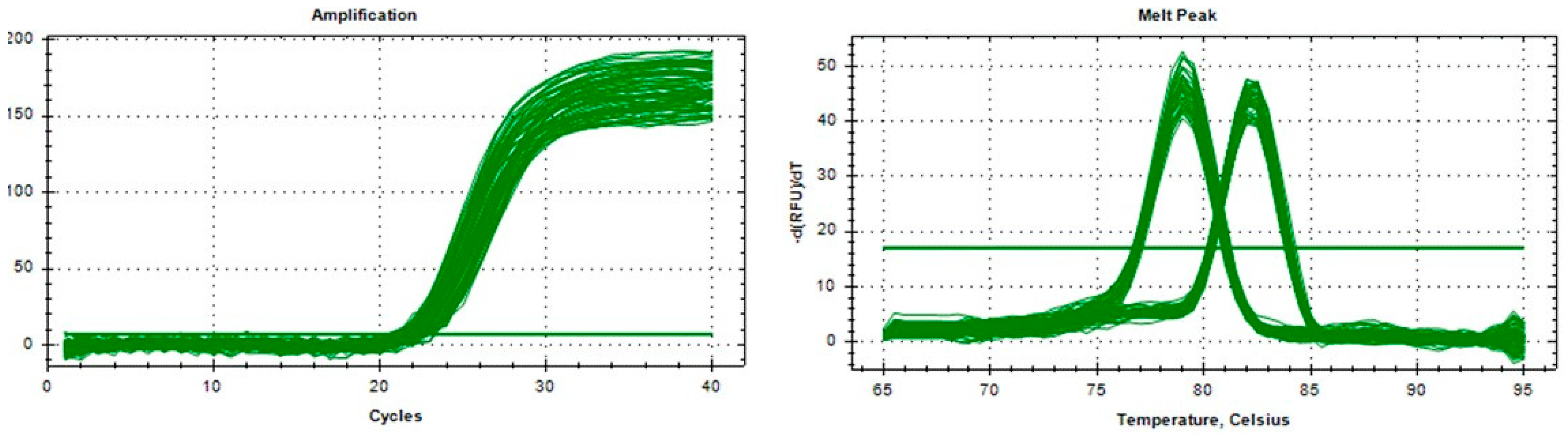
2.5. Copy Number Variation and Gene Expression Analysis of the SHE Gene
2.6. Statistical Analysis
3. Results
3.1. The Distribution of SHE Gene’s CNV in Four Sheep Breeds
3.2. Differences in CNV Distribution of the SHE Gene among Four Sheep Breeds
3.3. Association between the Copy Number Variation and Growth Traits
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mills, R.E.; Walter, K.; Stewart, C.; Handsaker, R.E.; Chen, K.; Alkan, C.; Abyzov, A.; Yoon, S.C.; Ye, K.; Cheetham, R.K.; et al. Mapping copy number variation by population-scale genome sequencing. Nature 2011, 470, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Henrichsen, C.N.; Vinckenbosch, N.; Zöllner, S.; Chaignat, E.; Pradervand, S.; Schütz, F.; Ruedi, M.; Kaessmann, H.; Reymond, A. Segmental copy number variation shapes tissue transcriptomes. Nat. Genet. 2009, 41, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Liu, X.; Pan, J.; Ma, L.; Ma, Y.; He, X.; Zhao, Q.; Pu, Y.; Li, Y.; Jiang, L. Genome-wide detection of copy number variation in Chinese indigenous sheep using an ovine high-density 600 K SNP array. Sci. Rep. 2017, 7, 912. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Fan, H.; Yuan, Z.; Hu, S.; Ma, X.; Xuan, J.; Wang, H.; Zhang, L.; Wei, C.; Zhang, Q.; et al. Genome-wide detection of CNVs in Chinese indigenous sheep with different types of tails using ovine high-density 600K SNP arrays. Sci. Rep. 2016, 6, 27822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Xu, L.; Zhou, Y.; Liu, M.; Wang, L.; Kijas, J.W.; Zhang, H.; Li, L.; Liu, G.E. Diversity of copy number variation in a worldwide population of sheep. Genomics 2018, 110, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Sadowski, I.; Stone, J.C.; Pawson, T. A noncatalytic domain conserved among cytoplasmic protein-tyrosine kinases modifies the kinase function and transforming activity of Fujinami sarcoma virus P130gag-fps. Mol. Cell. Biol. 1986, 6, 4396–4408. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.B.; Breed, J.; Barton, G.J. Conservation analysis and structure prediction of the SH2 family of phosphotyrosine binding domains. FEBS Lett. 1992, 304, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Hosoe, Y.; Numoto, N.; Inaba, S.; Ogawa, S.; Morii, H.; Abe, R.; Ito, N.; Oda, M. Structural and functional properties of Grb2 SH2 dimer in CD28 binding. Biophys. Physicobiol. 2019, 16, 80–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherif, N.; D’Hermies, F.; Barraco, P.; Elmaleh, C.; Renard, G.; Pouliquen, Y. Meibomian adenocarcinoma in its blepharo-conjunctival form. Apropos of a case. J. Fr. Ophtalmol. 1997, 20, 293–296. [Google Scholar]
- Sonstegard, T.S.; Garrett, W.M.; Ashwell, M.S.; Bennett, G.L.; Kappes, S.M.; Van Tassell, C.P. Comparative map alignment of BTA27 and HSA4 and 8 to identify conserved segments of genome containing fat deposition QTL. Mamm. Genome 2000, 11, 682–688. [Google Scholar] [CrossRef]
- Bae, J.S.; Cheong, H.S.; Kim, L.H.; NamGung, S.; Park, T.J.; Chun, J.Y.; Kim, J.Y.; Pasaje, C.F.; Lee, J.S.; Shin, H.D. Identification of copy number variations and common deletion polymorphisms in cattle. BMC Genomics 2010, 11, 232. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, X.; Ma, L.; Xu, H.; Cao, X.; Luo, R.; Chen, H.; Sun, X.; Cai, Y.; Lan, X. Detection of a new 20-bp insertion/deletion (indel) within sheep PRND gene using mathematical expectation (ME) method. Prion 2017, 11, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate—A practical and powerful approach to multiple testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Falconer, D.S.; Mackay, T.F.C. Introduction to quantitative genetics, 4th ed. Am. J. Hum. Genet. 1996, 46, 1231. [Google Scholar]
- Riollet, C.; Rainard, P.; Poutrel, B. Differential induction of complement fragment C5a and inflammatory cytokines during intramammary infections with Escherichia coli and Staphylococcus aureus. Clin. Diagn. Lab. Immunol. 2000, 7, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Feuk, L.; Carson, A.R.; Scherer, S.W. Structural variation in the human genome. Nat. Rev. Genet. 2006, 7, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Redon, R.; Ishikawa, S.; Fitch, K.R.; Feuk, L.; Perry, G.H.; Andrews, T.D.; Fiegler, H.; Shapero, M.H.; Carson, A.R.; Chen, W.; et al. Global variation in copy number in the human genome. Nature 2006, 444, 444–454. [Google Scholar] [CrossRef] [Green Version]
- Conrad, D.F.; Pinto, D.; Redon, R.; Feuk, L.; Gokcumen, O.; Zhang, Y.; Aerts, J.; Andrews, T.D.; Barnes, C.; Campbell, P.; et al. Origins and functional impact of copy number variation in the human genome. Nature 2010, 464, 704–712. [Google Scholar] [CrossRef]
- Zhang, F.; Gu, W.; Hurles, M.E.; Lupski, J.R. Copy Number Variation in Human Health, Disease, and Evolution. Annu. Rev. Genom. Hum. Genet. 2009, 10, 451–481. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.E.; Hou, Y.; Zhu, B.; Cardone, M.F.; Jiang, L.; Cellamare, A.; Mitra, A.; Alexander, L.J.; Coutinho, L.L.; Dell’Aquila, M.E.; et al. Analysis of copy number variations among diverse cattle breeds. Genome Res. 2010, 20, 693–703. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Nahashon, S.; Feaster, T.K.; Bohannon-Stewart, A.; Adefope, N. An initial map of chromosomal segmental copy number variations in the chicken. BMC Genomics 2010, 11, 351. [Google Scholar] [CrossRef] [PubMed]
- Fadista, J.; Nygaard, M.; Holm, L.-E.; Thomsen, B.; Bendixen, C. A Snapshot of CNVs in the Pig Genome. PLoS ONE 2008, 3, e3916. [Google Scholar] [CrossRef] [PubMed]
- Fontanesi, L.; Martelli, P.L.; Beretti, F.; Riggio, V.; Dall’Olio, S.; Colombo, M.; Casadio, R.; Russo, V.; Portolano, B.L. An initial comparative map of copy number variations in the goat (Capra hircus) genome. BMC Genomics 2010, 11, 639. [Google Scholar] [CrossRef] [PubMed]
- Fontanesi, L.; Beretti, F.; Martelli, P.; Colombo, M.; Dall’Olio, S.; Occidente, M.; Portolano, B.; Casadio, R.; Matassino, D.; Russo, V. A first comparative map of copy number variations in the sheep genome. Genomics 2011, 97, 158–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontanesi, L.; Martelli, P.L.; Scotti, E.; Russo, V.; Rogel-Gaillard, C.; Casadio, R.; Vernesi, C. Exploring copy number variation in the rabbit (Oryctolagus cuniculus) genome by array comparative genome hybridization. Genomics 2012, 100, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Geldermann, H. Investigations on inheritance of quantitative characters in animals by gene markers I. Methods. Theor. Appl. Genet. 1975, 46, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.K.; Huang, Y.Z.; Ma, Y.L.; Cheng, J.; Qu, Z.X.; Ma, Y.; Bai, Y.Y.; Tian, F.; Lin, F.P.; Ma, Y.L.; et al. Integrating CNVs into meta-QTL identified GBP4 as positional candidate for adult cattle stature. Funct. Integr. Genomics 2018, 18, 559–567. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, X.; Feng, C.; Song, C.; Hu, X.; Li, N. A genome-wide survey of copy number variation regions in various chicken breeds by array comparative genomic hybridization method. Anim. Genet. 2012, 43, 282–289. [Google Scholar] [CrossRef]
- Shi, T.; Xu, Y.; Yang, M.; Huang, Y.; Lan, X.; Lei, C.; Qi, X.; Yang, X.; Chen, H. Copy number variations at LEPR gene locus associated with gene expression and phenotypic traits in Chinese cattle. Anim. Sci. J. 2016, 87, 336–343. [Google Scholar] [CrossRef]
- Lehnert, S.A.; Reverter, A.; Byrne, K.A.; Wang, Y.; Nattrass, G.S.; Hudson, N.J.; Greenwood, P.L. Gene expression studies of developing bovine longissimus muscle from two different beef cattle breeds. BMC Dev. Biol. 2007, 7, 95. [Google Scholar] [CrossRef]
- Peng, Z.; Yuan, Y.; Li, Y.J.; Wang, H.X.; Shi, J.; Cao, W.X.; Luo, H.W.; Deng, J.R.; Feng, W.L. Targeting BCR tyrosine177 site with novel SH2-DED causes selective leukemia cell death in vitro and in vivo. Int. J. Biochem. Cell Biol. 2012, 44, 861–868. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Pairs Sequences (5′–3′) | Amplification Length (bp) | Tm (°C) |
|---|---|---|---|
| SHE | 1F: AACAAAGCGCATTTAGGGCA | 174 | 60 |
| 1R: ACGTCATGATCCAGCGATAGT | 174 | 60 | |
| ANKRD1 | 2F: TGGGCACCACGAAATTCTCA | 143 | 60 |
| 2R: TGGCAGAAATGTGCGAACG | 143 | 60 |
| Breeds | Loss | Normal | Gain |
|---|---|---|---|
| CKS | 0.56 | 0.28 | 0.16 |
| HS | 0.76 | 0.12 | 0.12 |
| STHS | 0.34 | 0.29 | 0.37 |
| LTHS | 0.10 | 0.49 | 0.41 |
| Breed | CKS | HS | STHS | LTHS |
|---|---|---|---|---|
| CKS | 22.886 (p = 1.07 × 10−5) | 45.736 (p = 1.17 × 10−10) | 33.844 (p = 4.47 × 10−8) | |
| HS | 84.897 (p = 3.67 × 10−19) | 67.481 (p = 2.22 × 10−15) | ||
| STHS | 15.809 (p = 3.69 × 10−4) | |||
| LTHS |
| Breed | Growth Traits | CNV Type (Mean ± SE) | p | ||
|---|---|---|---|---|---|
| Loss | Normal | Gain | |||
| CKS | Body length (cm) | 71.66 ± 1.60 b | 74.22 ± 1.588 a | 73.33 ± 1.59 a,b | 0.028 |
| HS | Circumference of cannon bone (cm) | 76.39 ± 0.36 c | 76.40 ± 0.89 a,b | 78.77 ± 0.89 a | 0.046 |
| STHS | Heart girth (cm) | 7.03 ± 0.04 b | 7.29 ± 0.11 a,b | 7.31 ± 0.11 a | 0.012 |
| Chest width (cm) | 19.58 ± 0.39 a,b | 20.26 ± 0.44 a | 18.60 ± 0.38 b | 0.019 | |
| High at the cross (cm) | 62.06 ± 0.51 B | 62.02 ± 0.58 B,C | 64.20 ± 0.50 A | 0.004 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, R.; Cheng, J.; Cao, X.-K.; Ma, Y.-L.; Chaogetu, B.; Huang, Y.-Z.; Lan, X.-Y.; Lei, C.-Z.; Hu, L.-Y.; Chen, H. Copy Number Variation of the SHE Gene in Sheep and Its Association with Economic Traits. Animals 2019, 9, 531. https://doi.org/10.3390/ani9080531
Jiang R, Cheng J, Cao X-K, Ma Y-L, Chaogetu B, Huang Y-Z, Lan X-Y, Lei C-Z, Hu L-Y, Chen H. Copy Number Variation of the SHE Gene in Sheep and Its Association with Economic Traits. Animals. 2019; 9(8):531. https://doi.org/10.3390/ani9080531
Chicago/Turabian StyleJiang, Rui, Jie Cheng, Xiu-Kai Cao, Yi-Lei Ma, Buren Chaogetu, Yong-Zhen Huang, Xian-Yong Lan, Chu-Zhao Lei, Lin-Yong Hu, and Hong Chen. 2019. "Copy Number Variation of the SHE Gene in Sheep and Its Association with Economic Traits" Animals 9, no. 8: 531. https://doi.org/10.3390/ani9080531