The Relationship between Embryonic Development and the Efficiency of Target Mutations in Porcine Endogenous Retroviruses (PERVs) Pol Genes in Porcine Embryos

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. General
2.2. Oocyte Collection, in Vitro Maturation, Fertilization, and Embryo Culture
2.3. Electroporation
2.4. Analysis of Targeted Gene Sequence
2.5. Statistical Analysis
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cooper, D.K. A brief history of cross-species organ transplantation. Bayl. Univ. Med. Cent. Proc. 2012. [Google Scholar] [CrossRef]
- Magre, S.; Takeuchi, Y.; Bartosch, B. Xenotransplantation and pig endogenous retroviruses. Rev. Med. Virol. 2003, 13, 311–329. [Google Scholar] [CrossRef] [PubMed]
- Fiebig, U.; Fischer, K.; Bahr, A.; Runge, C.; Schnieke, A.; Wolf, E.; Denner, J. Porcine endogenous retroviruses: Quantification of the copy number in cell lines, pig breeds, and organs. Xenotransplantation 2018, 25, e12445. [Google Scholar] [CrossRef]
- Nelson, P.N.; Carnegie, P.; Martin, J.; Ejtehadi, H.D.; Hooley, P.; Roden, D.; Rowland-Jones, S.; Warren, P.; Astley, J.; Murray, P.G. Demystified... Human endogenous retroviruses. Mol. Pathol. 2003, 56, 11. [Google Scholar] [CrossRef] [PubMed]
- Kimsa, M.C.; Strzalka-Mrozik, B.; Kimsa, M.W.; Gola, J.; Nicholson, P.; Lopata, K.; Mazurek, U. Porcine endogenous retroviruses in xenotransplantation—Molecular aspects. Viruses 2014, 6, 2062–2083. [Google Scholar] [CrossRef] [PubMed]
- Lopata, K.; Wojdas, E.; Nowak, R.; Lopata, P.; Mazurek, U. Porcine Endogenous Retrovirus (PERV)—Molecular Structure and Replication Strategy in the Context of Retroviral Infection Risk of Human Cells. Front. Microbiol. 2018, 9, 730. [Google Scholar] [CrossRef] [PubMed]
- Ragheb, J.A.; Anderson, W.F. Uncoupled expression of Moloney murine leukemia virus envelope polypeptides SU and TM: A functional analysis of the role of TM domains in viral entry. J. Virol. 1994, 68, 3207–3219. [Google Scholar] [PubMed]
- Takeuchi, Y.; Patience, C.; Magre, S.; Weiss, R.A.; Banerjee, P.T.; Le Tissier, P.; Stoye, J.P. Host range and interference studies of three classes of pig endogenous retrovirus. J. Virol. 1998, 72, 9986–9991. [Google Scholar] [CrossRef][Green Version]
- Watanabe, R.; Miyazawa, T.; Matsuura, Y. Cell-binding properties of the envelope proteins of porcine endogenous retroviruses. Microbes Infect. 2005, 7, 658–665. [Google Scholar] [CrossRef]
- Denner, J.; Tonjes, R.R. Infection barriers to successful xenotransplantation focusing on porcine endogenous retroviruses. Clin. Microbiol. Rev. 2012, 25, 318–343. [Google Scholar]
- Mang, R.; Maas, J.; Chen, X.; Goudsmit, J.; van der Kuyl, A.C. Identification of a novel type C porcine endogenous retrovirus: Evidence that copy number of endogenous retroviruses increases during host inbreeding. J. Gen. Virol. 2001, 82, 1829–1834. [Google Scholar] [CrossRef]
- Liu, G.; Li, Z.; Pan, M.; Ge, M.; Wang, Y.; Gao, Y. Genetic prevalence of porcine endogenous retrovirus in chinese experimental miniature pigs. Transplant. Proc. 2011. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Sakuma, T.; Yamamoto, T.; Mashimo, T. Simple knockout by electroporation of engineered endonucleases into intact rat embryos. Sci. Rep. 2014, 4, 6382. [Google Scholar] [CrossRef] [PubMed]
- Shan, Q.; Wang, Y.; Li, J.; Zhang, Y.; Chen, K.; Liang, Z.; Zhang, K.; Liu, J.; Xi, J.J.; Qiu, J.-L. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 686. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Güell, M.; Niu, D.; George, H.; Lesha, E.; Grishin, D.; Aach, J.; Shrock, E.; Xu, W.; Poci, J. Genome-wide inactivation of porcine endogenous retroviruses (PERVs). Science 2015, 350, 1101–1104. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Wei, H.-J.; Lin, L.; George, H.; Wang, T.; Lee, I.-H.; Zhao, H.-Y.; Wang, Y.; Kan, Y.; Shrock, E. Inactivation of porcine endogenous retrovirus in pigs using CRISPR-Cas9. Science 2017, 357, 1303–1307. [Google Scholar] [CrossRef] [PubMed]
- Tanihara, F.; Hirata, M.; Nguyen, N.T.; Le, Q.A.; Hirano, T.; Takemoto, T.; Nakai, M.; Fuchimoto, D.I.; Otoi, T. Generation of PDX-1 mutant porcine blastocysts by introducing CRISPR/Cas9-system into porcine zygotes via electroporation. Anim. Sci. J. 2019, 90, 55–61. [Google Scholar] [CrossRef]
- Naito, Y.; Hino, K.; Bono, H.; Ui-Tei, K. CRISPRdirect: Software for designing CRISPR/Cas guide RNA with reduced off-target sites. Bioinformatics 2015, 31, 1120–1123. [Google Scholar] [CrossRef]
- Patience, C.; Takeuchi, Y.; Weiss, R.A. Infection of human cells by an endogenous retrovirus of pigs. Nat. Med. 1997, 3, 282–286. [Google Scholar] [CrossRef]
- Wilson, C.A.; Wong, S.; Muller, J.; Davidson, C.E.; Rose, T.M.; Burd, P. Type C retrovirus released from porcine primary peripheral blood mononuclear cells infects human cells. J. Virol. 1998, 72, 3082–3087. [Google Scholar]
- Anderson, E.M.; Haupt, A.; Schiel, J.A.; Chou, E.; Machado, H.B.; Strezoska, Z.; Lenger, S.; McClelland, S.; Birmingham, A.; Vermeulen, A.; et al. Systematic analysis of CRISPR-Cas9 mismatch tolerance reveals low levels of off-target activity. J. Biotechnol. 2015, 211, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wei, J.J.; Sabatini, D.M.; Lander, E.S. Genetic screens in human cells using the CRISPR-Cas9 system. Science 2014, 343, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Cradick, T.J.; Qiu, P.; Lee, C.M.; Fine, E.J.; Bao, G. COSMID: A Web-Based Tool for Identifying and Validating CRISPR/Cas Off-Target Sites. Mol. Ther.-Nucleic Acids 2014, 3, e214. [Google Scholar] [CrossRef] [PubMed]
- Tanihara, F.; Takemoto, T.; Kitagawa, E.; Rao, S.; Do, L.T.K.; Onishi, A.; Yamashita, Y.; Kosugi, C.; Suzuki, H.; Sembon, S. Somatic cell reprogramming-free generation of genetically modified pigs. Sci. Adv. 2016, 2, e1600803. [Google Scholar] [CrossRef] [PubMed]
- Nishio, K.; Tanihara, F.; Nguyen, T.V.; Kunihara, T.; Nii, M.; Hirata, M.; Takemoto, T.; Otoi, T. Effects of voltage strength during electroporation on the development and quality of In Vitro-produced porcine embryos. Reprod. Domest. Anim. 2018, 53, 313–318. [Google Scholar] [CrossRef]
- Brinkman, E.K.; Chen, T.; Amendola, M.; van Steensel, B. Easy quantitative assessment of genome editing by sequence trace decomposition. Nucleic Acids Res. 2014, 42, e168. [Google Scholar] [CrossRef]
- Yonezawa, K.; Imanishi, S.; Sasaki, Y.; Fujii, H. Biological Statistics; Asakura Publishing: Tokyo, Japan, 1988. (In Japanese) [Google Scholar]
- Ross, M.J.; Coates, P.T. Using CRISPR to inactivate endogenous retroviruses in pigs: An important step toward safe xenotransplantation? Kidney Int. 2018, 93, 4–6. [Google Scholar] [CrossRef]
- Arav, A.; Aroyo, A.; Yavin, S.; Roth, Z. Prediction of embryonic developmental competence by time-lapse observation and ‘shortest-half’ analysis. Reprod. BioMed. Online 2008, 17, 669–675. [Google Scholar] [CrossRef]
- Isom, S.C.; Li, R.F.; Whitworth, K.M.; Prather, R.S. Timing of first embryonic cleavage is a positive indicator of the In Vitro developmental potential of porcine embryos derived from In Vitro fertilization, somatic cell nuclear transfer and parthenogenesis. Mol. Reprod. Dev. 2012, 79, 197–207. [Google Scholar] [CrossRef]
- Doench, J.G.; Hartenian, E.; Graham, D.B.; Tothova, Z.; Hegde, M.; Smith, I.; Sullender, M.; Ebert, B.L.; Xavier, R.J.; Root, D.E. Rational design of highly active sgRNAs for CRISPR-Cas9-mediated gene inactivation. Nat. Biotechnol. 2014, 32, 1262–1267. [Google Scholar] [CrossRef]
- Quereda, J.J.; Herrero-Medrano, J.M.; Abellaneda, J.M.; Garcia-Nicolas, O.; Martinez-Alarcon, L.; Pallares, F.J.; Ramirez, P.; Munoz, A.; Ramis, G. Porcine endogenous retrovirus copy number in different pig breeds is not related to genetic diversity. Zoonoses Public Health 2012, 59, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Lee, J.; Yoon, J.K.; Kim, N.Y.; Kim, G.W.; Park, C.; Oh, Y.K.; Kim, Y.B. Rapid determination of perv copy number from porcine genomic DNA by real-time polymerase chain reaction. Anim. Biotechnol. 2011, 22, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Patience, C.; Switzer, W.M.; Takeuchi, Y.; Griffiths, D.J.; Goward, M.E.; Heneine, W.; Stoye, J.P.; Weiss, R.A. Multiple groups of novel retroviral genomes in pigs and related species. J. Virol. 2001, 75, 2771–2775. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
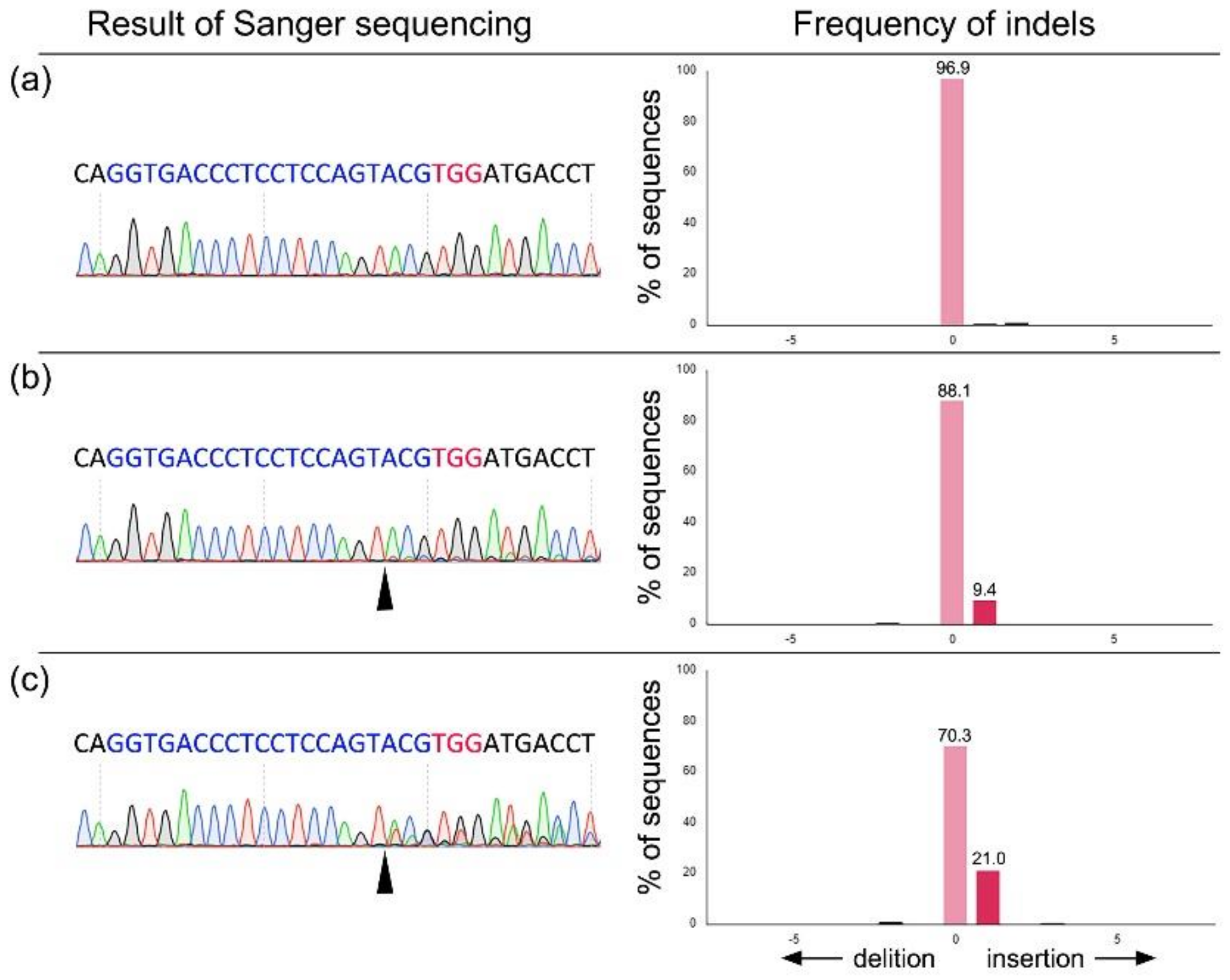{kind=link}
| gRNA †† | No. of Oocytes Examined | No. (Mean ± SEM) of Embryos | No. of Blastocysts Examined | No. (Mean) of Mutated Blastocysts ††† | Total Mutation Efficiency (Mean ± SEM) †††† | |
|---|---|---|---|---|---|---|
| Cleaved | Developed to Blastocysts | |||||
| Control | 184 | 159 (86.1± 1.5) | 41 (21.5 ± 3.9) a | - | - | - |
| gRNA1 | 267 | 227 (85.1 ± 2.6) | 22 (9.2 ± 3.4) b | 18 | 13 (72.2) | 12.4 ± 2.2 |
| gRNA2 | 263 | 220 (83.9 ± 2.4) | 11 (4.3 ± 1.6) b,c | 10 | 5 (50.0) | 11.8 ± 2.1 |
| gRNA3 | 263 | 224 (85.5 ± 3.1) | 1 (0.4 ± 0.4) c | 1 | 1 (100) | 18.1 |
| gRNA †† | No. of Oocytes Examined | No. (Mean ± SEM) of Embryos | No. of Blastocysts Examined | No. (Mean) of Mutated Blastocysts ††† | Total Mutation Efficiency (Mean ± SEM) †††† | |
|---|---|---|---|---|---|---|
| Cleaved | Developed to Blastocysts | |||||
| Control | 157 | 124 (79.4 ± 3.2) a | 28 (14.4 ± 1.5) a | - | - | - |
| gRNA1 & gRNA2 | 230 | 204 (88.9 ± 2.1) b | 11 (4.7 ± 1.0) b | 10 | 8 (80.0) | 11.0 ± 3.1 |
| gRNA1 & gRNA3 | 197 | 146 (74.0 ± 3.2) a | 2 (1.1 ± 0.6) c | 2 | 1 (50.0) | 15.9 |
| gRNA2 & gRNA3 | 222 | 174 (78.8 ± 4.6) a | 0 (0) c | - | - | - |
| gRNA †† | No. ofPooled Embryo Sample | No. (Mean) of Mutated Embryo Sample † | Total Mutation Efficiency (Mean ± SEM) †† |
|---|---|---|---|
| gRNA1 | 10 | 10 (100) | 32.9 ± 3.1 a,b |
| gRNA2 | 9 | 8 (88.9) | 17.1 ± 2.9 a |
| gRNA3 | 8 | 8 (100) | 44.0 ± 5.1 b |
| gRNA1 & gRNA2 | 15 | 12 (80.0) | 21.1 ± 4.1 a |
| gRNA1 & gRNA3 | 16 | 14 (87.5) | 59.9 ± 6.2 c |
| gRNA2 & gRNA3 | 10 | 10 (100) | 42.2 ± 5.8 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirata, M.; Wittayarat, M.; Hirano, T.; Nguyen, N.T.; Le, Q.A.; Namula, Z.; Fahrudin, M.; Tanihara, F.; Otoi, T. The Relationship between Embryonic Development and the Efficiency of Target Mutations in Porcine Endogenous Retroviruses (PERVs) Pol Genes in Porcine Embryos. Animals 2019, 9, 593. https://doi.org/10.3390/ani9090593
Hirata M, Wittayarat M, Hirano T, Nguyen NT, Le QA, Namula Z, Fahrudin M, Tanihara F, Otoi T. The Relationship between Embryonic Development and the Efficiency of Target Mutations in Porcine Endogenous Retroviruses (PERVs) Pol Genes in Porcine Embryos. Animals. 2019; 9(9):593. https://doi.org/10.3390/ani9090593
Chicago/Turabian StyleHirata, Maki, Manita Wittayarat, Takayuki Hirano, Nhien Thi Nguyen, Quynh Anh Le, Zhao Namula, Mokhamad Fahrudin, Fuminori Tanihara, and Takeshige Otoi. 2019. "The Relationship between Embryonic Development and the Efficiency of Target Mutations in Porcine Endogenous Retroviruses (PERVs) Pol Genes in Porcine Embryos" Animals 9, no. 9: 593. https://doi.org/10.3390/ani9090593
APA StyleHirata, M., Wittayarat, M., Hirano, T., Nguyen, N. T., Le, Q. A., Namula, Z., Fahrudin, M., Tanihara, F., & Otoi, T. (2019). The Relationship between Embryonic Development and the Efficiency of Target Mutations in Porcine Endogenous Retroviruses (PERVs) Pol Genes in Porcine Embryos. Animals, 9(9), 593. https://doi.org/10.3390/ani9090593