Assessing and Enhancing the Welfare of Animals with Equivocal and Reliable Cues


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
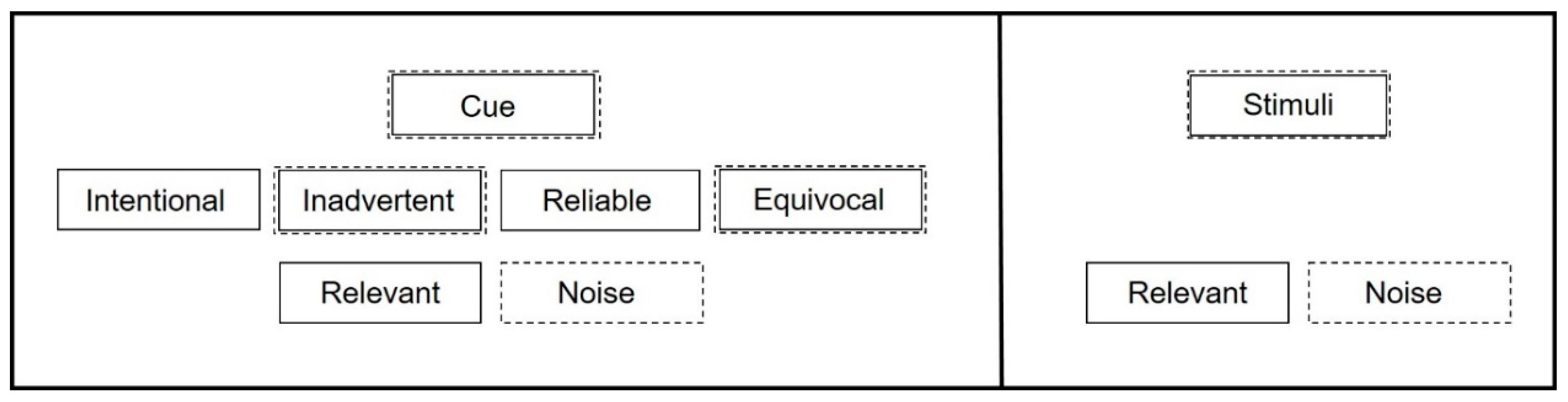
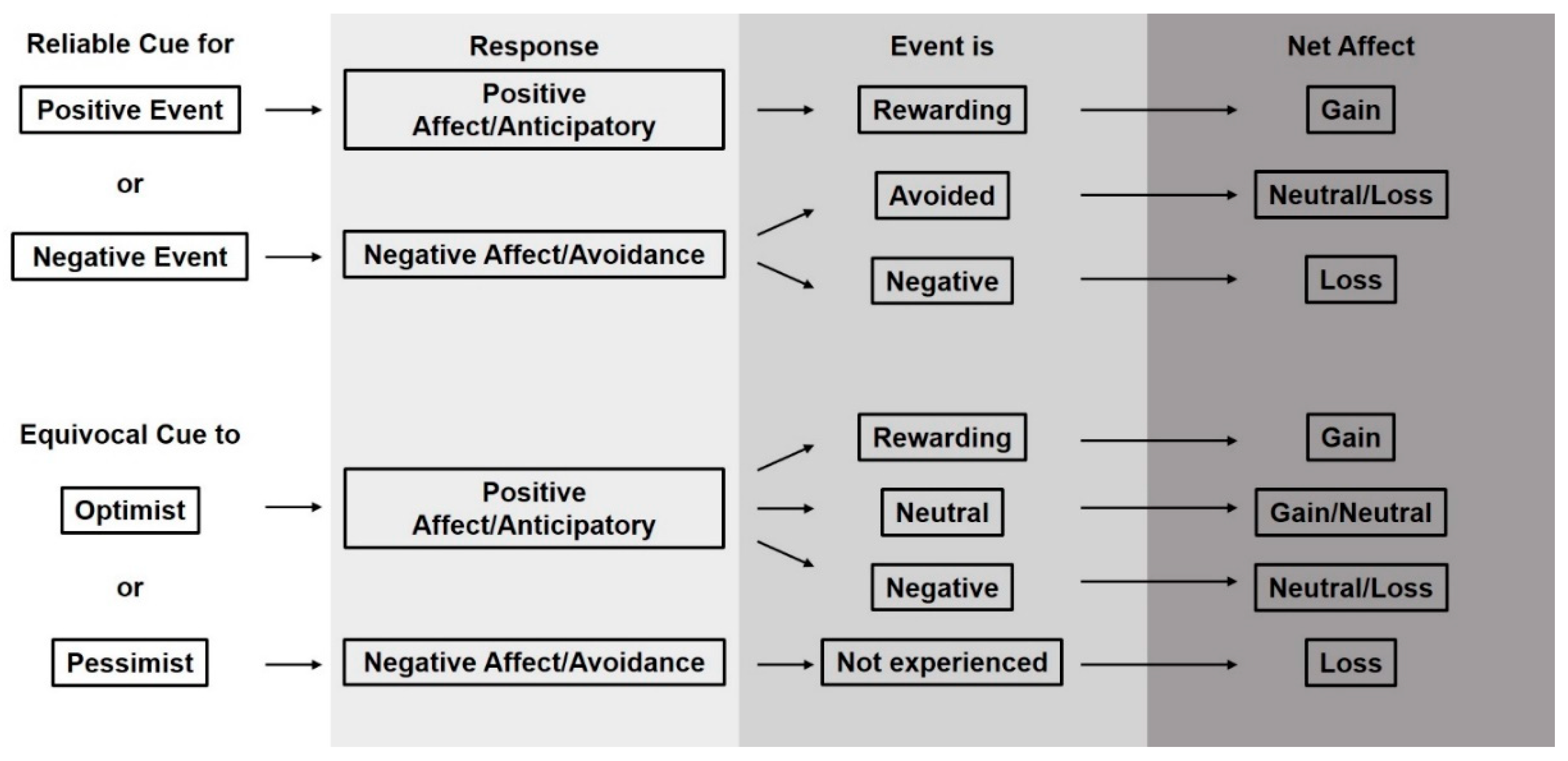
2. How Do Cues Relate to Animal Welfare?
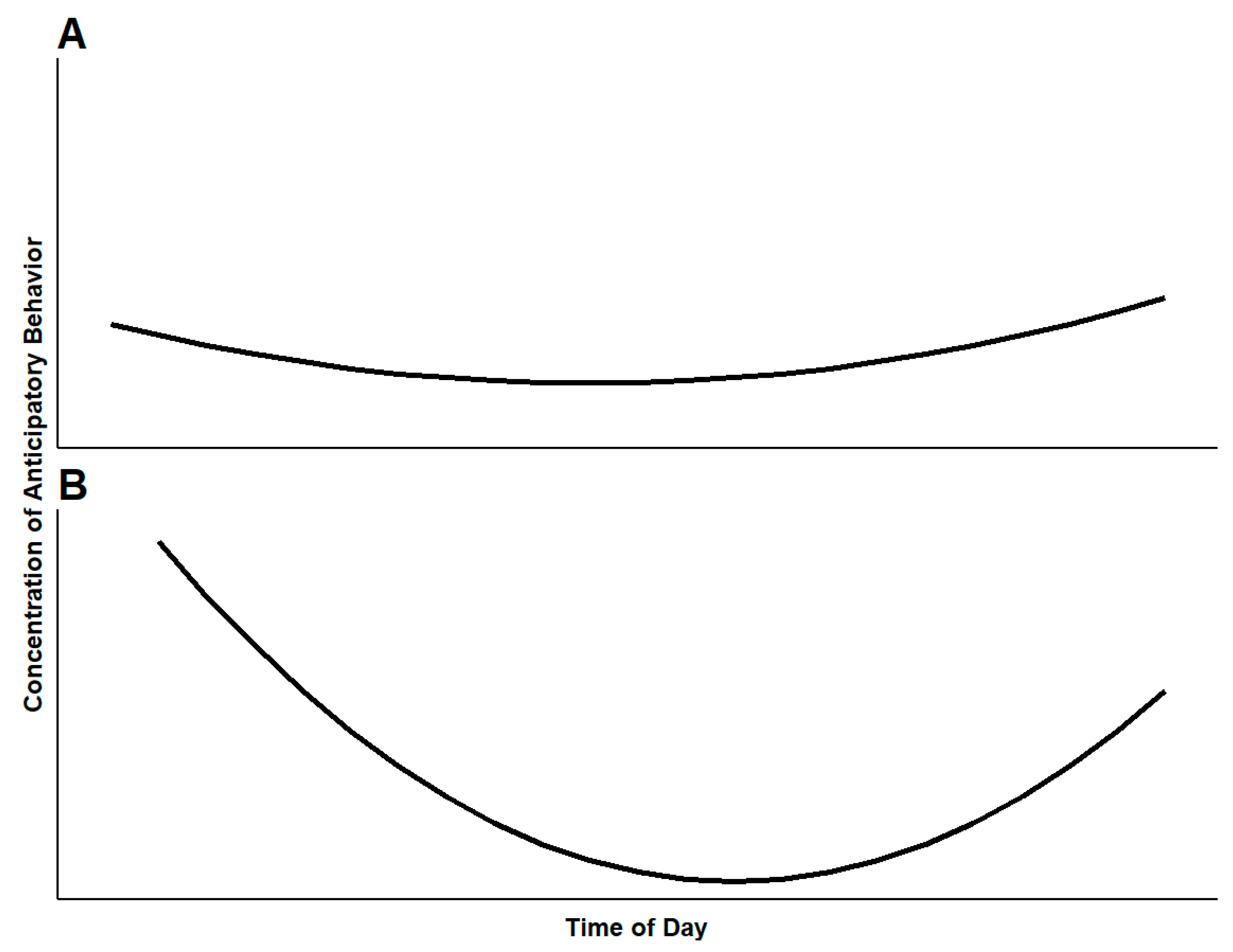
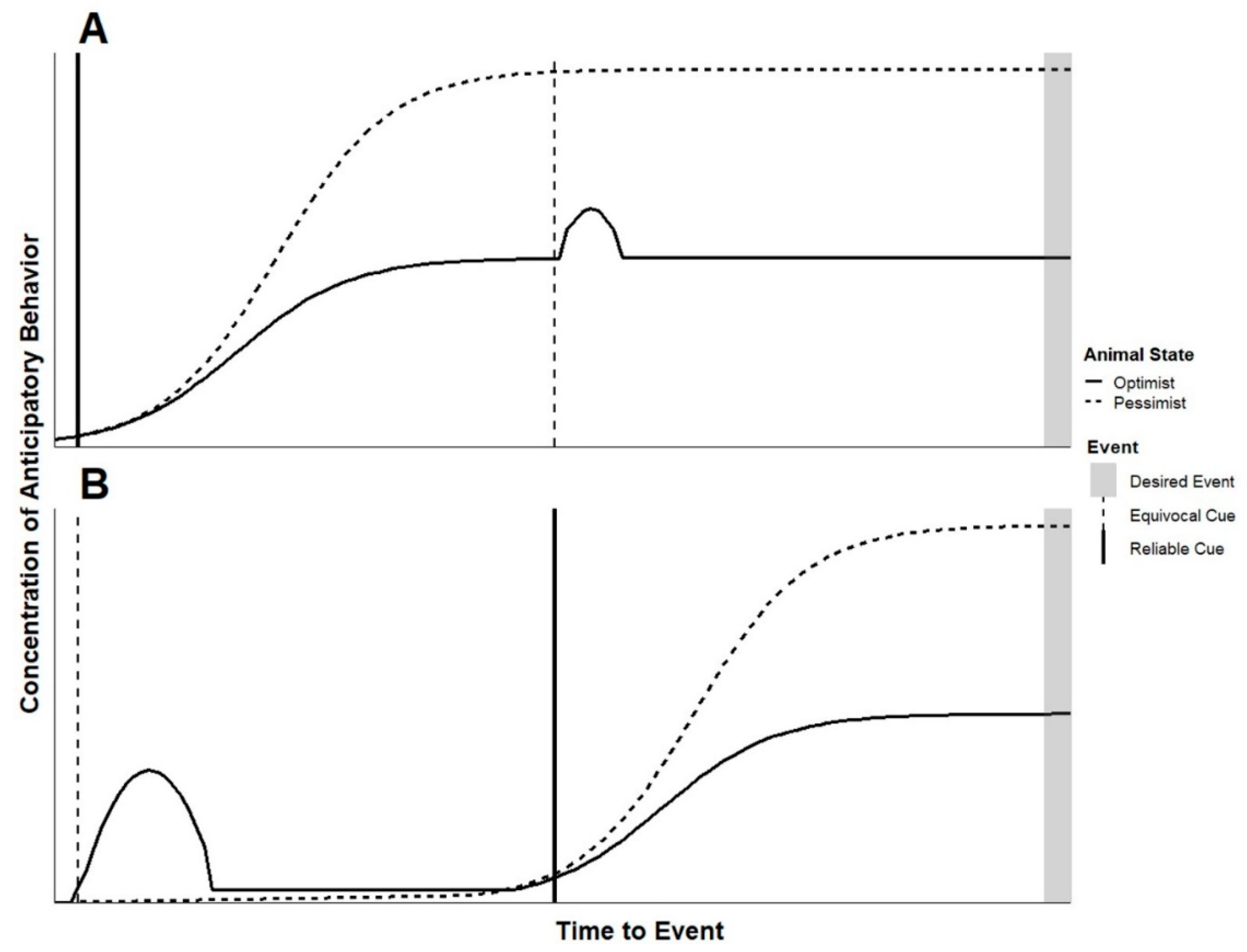
3. How Do Cues Help Us Understand Welfare?
4. How Do Cues Support Welfare?
5. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Watters, J.V. Toward A Predictive Theory for Environmental Enrichment. Zoo Biol. 2009, 28, 608–622. [Google Scholar] [CrossRef] [PubMed]
- Watters, J.V. Searching for Behavioral Indicators of Welfare in Zoos: Uncovering Anticipatory Behavior. Zoo Biol. 2014, 33, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Wielebnowski, N.C.; Fletchall, N.; Carlstead, K.; Busso, J.M.; Brown, J.L. Noninvasive assessment of adrenal activity associated with husbandry and behavioral factors in the North American clouded leopard population. Zoo Biol. 2002, 21, 77–98. [Google Scholar] [CrossRef]
- Carlstead, K.; Mench, J.A.; Meehan, C.; Brown, J.L. An Epidemiological Approach to Welfare Research in Zoos: The Elephant Welfare Project. J. Appl. Anim. Welf. Sci. 2013, 16, 319–337. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.; Torgerson-White, L.; Fripp, D.; Watters, J.; Petric, A. A Multi-Institutional Assessment of Factors Influencing Locomotion and Pacing in Captive Okapis (Okapia johnstoni). J. Appl. Anim. Welf. Sci. 2015, 18, S43–S61. [Google Scholar] [CrossRef] [PubMed]
- Krebs, B.L.; Torres, E.; Chesney, C.; Kantoniemi Moon, V.; Watters, J.V. Applying Behavioral Conditioning to Identify Anticipatory Behaviors. J. Appl. Anim. Welf. Sci. 2017, 20, 155–175. [Google Scholar] [CrossRef] [PubMed]
- Krebs, B.L.; Watters, J.V. Simple but temporally unpredictable puzzles are cognitive enrichment. Anim. Behav. Cogn. 2017, 4, 119–134. [Google Scholar] [CrossRef] [Green Version]
- Krebs, B.L.; Watters, J.V. Using Technology Driven Environments to Promote Animal Well—Being in Zoos. In Proceedings of the Human Computer Interactions, San Jose, CA, USA, 7–12 May 2016. [Google Scholar]
- Watters, J.V.; Miller, J.T.; Sullivan, T.J. Note on Optimizing Environmental Enrichment: A Study of Fennec Fox and Zoo Guests. Zoo Biol. 2011, 30, 647–654. [Google Scholar] [CrossRef]
- Meehan, C.L.; Mench, J.A. The challenge of challenge: Can problem solving opportunities enhance animal welfare? Appl. Anim. Behav. Sci. 2007, 102, 246–261. [Google Scholar] [CrossRef]
- Clark, F.E. Cognitive enrichment and welfare: Current approaches and future directions. Anim. Behav. Cogn. 2017, 4, 52–71. [Google Scholar] [CrossRef] [Green Version]
- Berger, A. Activity patterns, chronobiology and the assessment of stress and welfare in zoo and wild animals. Int. Zoo Yearb. 2010, 45, 80–90. [Google Scholar] [CrossRef]
- Shepherdson, D.; Lewis, K.D.; Carlstead, K.; Bauman, J.; Perrin, N. Individual and environmental factors associated with stereotypic behavior and fecal glucocorticoid metabolite levels in zoo housed polar bears. Appl. Anim. Behav. Sci. 2013, 147, 268–277. [Google Scholar] [CrossRef]
- Eguizábal, G.V.; Palme, R.; Superina, M.; Asencio, C.J.; García Capocasa, M.C.; Busso, J.M. Characterization and correlations of behavioral and adrenocortical activities of zoo-housed lesser anteaters (Tamandua tetradactyla). Zoo Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Bethell, E.J. A “How-To” Guide for Designing Judgment Bias Studies to Assess Captive Animal Welfare. J. Appl. Anim. Welf. Sci. 2015, 18, S18–S42. [Google Scholar] [CrossRef] [PubMed]
- Ward, S.J.; Sherwen, S.; Clark, F.E. Advances in Applied Zoo Animal Welfare Science. J. Appl. Anim. Welf. Sci. 2018, 21, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clegg, L.I. Cognitive Bias in Zoo Animals: An Optimistic Outlook for Welfare Assessment. Animals 2018, 8, 104. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, S.E.; Ritvo, S. Comparative Cognition Outside the Laboratory. Comp. Cogn. Behav. Rev. 2016, 11, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, S.L.; Kwiatt, A.C.; Ross, S.R.; Cronin, K.A. The effects of cognitive testing on the welfare of zoo-housed Japanese macaques (Macaca fuscata). Appl. Anim. Behav. Sci. 2019, 212, 90–97. [Google Scholar] [CrossRef]
- Hill, S.P.; Broom, D.M. Measuring zoo animal welfare: theory and practice. Zoo Biol. 2009, 28, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Boissy, A.; Manteuffel, G.; Jensen, M.B.; Moe, R.O.; Spruijt, B.; Keeling, L.J.; Winckler, C.; Forkman, B.; Dimitrov, I.; Langbein, J.; et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 2007, 92, 375–397. [Google Scholar] [CrossRef]
- Mellor, D. Updating Animal Welfare Thinking: Moving beyond the “Five Freedoms” towards “A Life Worth Living”. Animals 2016, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Mellor, D.J.; Beausoleil, N.J. Extending the ‘Five Domains’ model for animal welfare assessment to incorporate positive welfare states. Anim. Welf. 2015, 24, 241–253. [Google Scholar] [CrossRef]
- Panksepp, J. Integrating bottom-up internalist views of emotional feelings with top-down externalist views: Might brain affective changes constitute reward and punishment effects within animal brains? Cortex 2014, 59, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Panksepp, J. Cross-Species Neuroaffective Parsing of Primal Emotional Desires and Aversions in Mammals. Emot. Rev. 2013, 5, 235–240. [Google Scholar] [CrossRef]
- Panksepp, J.; Watt, D. What is Basic about Basic Emotions? Lasting Lessons from Affective Neuroscience. Emot. Rev. 2011, 3, 387–396. [Google Scholar] [CrossRef]
- Panksepp, J. The basic emotional circuits of mammalian brains: Do animals have affective lives? Neurosci. Biobehav. Rev. 2011, 35, 1791–1804. [Google Scholar] [CrossRef] [PubMed]
- Špinka, M.; Wemelsfelder, F. Environmental Challenge and Animal Agency. In Animal Welfare, 2nd ed.; Appleby, M.C., Mench, J.A., Olson, I.A.S., Hughes, B.O., Eds.; CAB International: Cambridge, MA, USA, 2011; pp. 27–43. [Google Scholar]
- Wemelsfelder, F. The scientific validity of subjective concepts in models of animal welfare. Appl. Anim. Behav. Sci. 1997, 53, 75–88. [Google Scholar] [CrossRef] [Green Version]
- Brando, S.; Buchanan-Smith, H.M. The 24/7 approach to promoting optimal welfare for captive wild animals. Behav. Process. 2018, 156, 83–95. [Google Scholar] [CrossRef]
- Kearney, B.Y.; Fleischer, B.J. Development of an instrument to measure exercise of self-care agency. Res. Nurs. Health 1979, 2, 25–34. [Google Scholar] [CrossRef]
- Cockerham, W.C. Health Lifestyle Theory and the Convergence of Agency and Structure. J. Health Soc. Behav. 2005, 46, 51–67. [Google Scholar] [CrossRef] [Green Version]
- Rimpley, K.; Buchanan-Smith, H.M. Reliably signalling a startling husbandry event improves welfare of zoo-housed capuchins (Sapajus apella). Appl. Anim. Behav. Sci. 2013, 147, 205–213. [Google Scholar] [CrossRef]
- Ward, S.J.; Melfi, V. Keeper-animal interactions: Differences between the behaviour of zoo animals affect stockmanship. PLoS ONE 2015, 10, e0140237. [Google Scholar] [CrossRef] [PubMed]
- Hemsworth, P.H. Ethical stockmanship. Aust. Vet. J. 2007, 85, 194–200. [Google Scholar] [CrossRef]
- Boivin, X.; Lensink, J.; Tallet, C.; Veissier, I. Stockmanship and farm animal welfare. Anim. Welf. 2003, 12, 479–492. [Google Scholar]
- Hemsworth, P.H.; Barnett, J.L.; Coleman, G.J. The Human-Animal Relationship in Agriculture and its Consequences for the Animal. Anim. Welf. 1993, 2, 33–51. [Google Scholar]
- Hemsworth, P.H.; Barnett, J.L.; Coleman, G.J. The integration of human-animal relations into animal welfare monitoring schemes. Anim. Welf. 2009, 18, 335–345. [Google Scholar]
- Grandin, T. Animal handling. Vet. Clin. N. Am. Food Anim. Pract. 1987, 3, 323–338. [Google Scholar] [CrossRef]
- Barker, S.B.; Wolen, A.R. The Benefits of Human–Companion Animal Interaction: A Review. J. Vet. Med. Educ. 2008, 35, 487–495. [Google Scholar] [CrossRef]
- Walsh, F. Human-Animal bonds II: The role of pets in family systems and family therapy. Fam. Process 2009, 48, 481–499. [Google Scholar] [CrossRef]
- Chelluri, G.I.; Ross, S.R.; Wagner, K.E. Behavioral correlates and welfare implications of informal interactions between caretakers and zoo-housed chimpanzees and gorillas. Appl. Anim. Behav. Sci. 2013, 147, 306–315. [Google Scholar] [CrossRef]
- Clegg, I.L.K.; Rödel, H.G.; Boivin, X.; Delfour, F. Looking forward to interacting with their caretakers: Dolphins’ anticipatory behaviour indicates motivation to participate in specific events. Appl. Anim. Behav. Sci. 2018, 202, 85–93. [Google Scholar] [CrossRef]
- Carlstead, K. A comparative approach to the study of keeper–animal relationships in the zoo. Zoo Biol. 2009, 28, 589–608. [Google Scholar] [CrossRef] [PubMed]
- Carlstead, K.; Paris, S.; Brown, J.L. Good keeper-elephant relationships in North American zoos are mutually beneficial to welfare. Appl. Anim. Behav. Sci. 2019, 211, 103–111. [Google Scholar] [CrossRef]
- Laidre, M.E.; Johnstone, R.A. Animal signals. Curr. Biol. 2013, 23, R829–R833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weary, D.M.; Fraser, D. Calling by domestic piglets: reliable signals of need? Anim. Behav. 1995, 50, 1047–1055. [Google Scholar] [CrossRef] [Green Version]
- Jensen, A.L.M.; Delfour, F.; Carter, T. Anticipatory behavior in captive bottlenose dolphins (Tursiops truncatus): A preliminary study. Zoo Biol. 2013, 32, 436–444. [Google Scholar] [CrossRef]
- Keen, H.A.; Nelson, O.L.; Robbins, C.T.; Evans, M.; Shepherdson, D.J.; Newberry, R.C. Validation of a novel cognitive bias task based on difference in quantity of reinforcement for assessing environmental enrichment. Anim. Cogn. 2014, 17, 529–541. [Google Scholar] [CrossRef]
- Bateson, M.; Matheson, S.M. Performance on a categorisation task suggests that removal of environmental enrichment induces ‘pessimism’ in captive European starlings (Sturnus vulgaris). Anim. Welf. 2007, 16, 33–36. [Google Scholar]
- Puppe, B.; Ernst, K.; Schon, P.C.; Manteuffel, G. Cognitive enrichment affects behavioural reactivity in domestic pigs. Appl. Anim. Behav. Sci. 2007, 105, 75–86. [Google Scholar] [CrossRef]
- Wood-Gush, D.G.M.; Vestergaard, K. Exploratory behavior and the welfare of intensively kept animals. J. Agric. Ethics 1989, 2, 161–169. [Google Scholar] [CrossRef]
- Kuczaj, S.; Lacinak, T.; Fad, O.; Trone, M.; Solangi, M.; Ramos, J. Keeping Environmental Enrichment Enriching. Int. J. Comp. Psychol. 2002, 15, 127–137. [Google Scholar]
- Markel, A.L.; Galaktionov, Y.K.; Efimov, V.M. Factor analysis of rat behavior in an open field test. Neurosci. Behav. Physiol. 1989, 19, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Boivin, X.; Le Neindre, P.; Chupin, J.M.; Garel, J.P.; Trillat, G. Influence of breed and early management on ease of handling and open-field behaviour of cattle. Appl. Anim. Behav. Sci. 1992, 32, 313–323. [Google Scholar] [CrossRef]
- Pare, W.P. Relationship of Various Behaviors in the Open-Field Test of Emotionality. Psychol. Rep. 1964, 14, 19–22. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 2015, e52434. [Google Scholar] [CrossRef] [PubMed]
- Zeligs, J. Animal Training 101: The Complete and Practical Guide to the Art and Science of Behavior Modification; Mill City Press: Minneapolis, MN, USA, 2014; pp. 33–94. [Google Scholar]
- Lawrence, D.H. Acquired distinctiveness of cues: I. Transfer between discriminations on the basis of familiarity with the stimulus. J. Exp. Psychol. 1949, 39, 770–784. [Google Scholar] [CrossRef]
- Franks, B.; Higgins, E.T. Effectiveness in Humans and Other Animals: A Common Basis for Well-Being and Welfare. In Advances in Experimental Social Psychology; Olson, J.M., Zanna, M.P., Eds.; Elsevier Academic Press Inc.: San Diego, CA, USA, 2012; Volume 46, pp. 285–346. [Google Scholar]
- Inglis, I.R.; Forkman, B.; Lazarus, J. Free food or earned food? A review and fuzzy model of contrafreeloading. Anim. Behav. 1997, 53, 1171–1191. [Google Scholar] [CrossRef] [Green Version]
- De Jonge, F.H.; Tilly, S.L.; Baars, A.M.; Spruijt, B.M. On the rewarding nature of appetitive feeding behaviour in pigs (Sus scrofa): Do domesticated pigs contrafreeload? Appl. Anim. Behav. Sci. 2008, 114, 359–372. [Google Scholar] [CrossRef]
- Langbein, J.; Siebert, K.; Nürnberg, G. On the use of an automated learning device by group-housed dwarf goats: Do goats seek cognitive challenges? Appl. Anim. Behav. Sci. 2009, 120, 150–158. [Google Scholar] [CrossRef]
- Horvath, K.; Angeletti, D.; Nascetti, G.; Carere, C. Invertebrate welfare: an overlooked issue. Annali Dell’istituto Superiore di Sanità 2013, 49, 9–17. [Google Scholar]
- Burghardt Gordon, M.; Ward, B.; Rosscoe, R. Problem of reptile play: Environmental enrichment and play behavior in a captive Nile soft-shelled turtle, Trionyx triunguis. Zoo Biol. 1996, 15, 223–238. [Google Scholar] [CrossRef]
- Bassett, L.; Buchanan-Smith, H.M. Effects of predictability on the welfare of captive animals. Appl. Anim. Behav. Sci. 2007, 102, 223–245. [Google Scholar] [CrossRef] [Green Version]
- Panksepp, J.; Burgdorf, J. 50-kHz chirping (laughter?) in response to conditioned and unconditioned tickle-induced reward in rats: effects of social housing and genetic variables. Behav. Brain Res. 2000, 115, 25–38. [Google Scholar] [CrossRef]
- Reimert, I.; Bolhuis, J.E.; Kemp, B.; Rodenburg, T.B. Indicators of positive and negative emotions and emotional contagion in pigs. Physiol. Behav. 2013, 109, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Castles, D.L.; Whiten, A.; Aureli, F. Social anxiety, relationships and self-directed behaviour among wild female olive baboons. Anim. Behav. 1999, 58, 1207–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troisi, A. Displacement Activities as a Behavioral Measure of Stress in Nonhuman Primates and Human Subjects. Stress 2002, 5, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Spruijt, B.M.; van den Bos, R.; Pijlman, F.T.A. A concept of welfare based on reward evaluating mechanisms in the brain: anticipatory behaviour as an indicator for the state of reward systems. Appl. Anim. Behav. Sci. 2001, 72, 145–171. [Google Scholar] [CrossRef]
- Van Der Harst, J.E.; Fermont, P.C.J.; Bilstra, A.E.; Spruijt, B.M. Access to enriched housing is rewarding to rats as reflected by their anticipatory behaviour. Anim. Behav. 2003, 66, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Dudink, S.; Simonse, H.; Marks, I.; De Jonge, F.H.; Spruijt, B.M. Announcing the arrival of enrichment increases play behaviour and reduces weaning-stress-induced behaviours of piglets directly after weaning. Appl. Anim. Behav. Sci. 2006, 101, 86–101. [Google Scholar] [CrossRef]
- Stephan, F.K. The “other” circadian system: Food as a zeitgeber. J. Biol. Rhythm. 2002, 17, 284–292. [Google Scholar] [CrossRef]
- Solis-Salazar, T.; Martinez-Merlos, M.T.; Angeles-Castellanos, M.; Mendoza, J.; Escobar, C. Behavioral and physiological adaptations in rats during food-entrainment. Biol. Rhythm Res. 2005, 36, 99–108. [Google Scholar] [CrossRef]
- Mistlberger, R.E. Circadian food-anticipatory activity: Formal models and physiological mechanisms. Neurosci. Biobehav. Rev. 1994, 18, 171–195. [Google Scholar] [CrossRef]
- Makowska, I.J.; Weary, D.M. Differences in Anticipatory Behaviour between Rats (Rattus norvegicus) Housed in Standard versus Semi-Naturalistic Laboratory Environments. PLoS ONE 2016, 11, e0147595. [Google Scholar] [CrossRef] [PubMed]
- Van der Harst, J.E.; Baars, A.M.; Spruijt, B.M. Standard housed rats are more sensitive to rewards than enriched housed rats as reflected by their anticipatory behaviour. Behav. Brain Res. 2003, 142, 151–156. [Google Scholar] [CrossRef]
- Van der Horst, J.E.; Spruijt, B.M. Tools to measure and improve animal welfare: reward-related behaviour. Anim. Welf. 2007, 16, 67–73. [Google Scholar]
- Clegg, I.; Delfour, F. Cognitive judgement bias is associated with frequency of anticipatory behavior in bottlenose dolphins. Zoo Biol. 2018, 37, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.; Yngvesson, J.; Boissy, A.; Uvnas-Moberg, K.; Lidfors, L. Behavioural expression of positive anticipation for food or opportunity to play in lambs. Behav. Process. 2015, 113, 152–158. [Google Scholar] [CrossRef]
- Harding, E.J.; Paul, E.S.; Mendl, M. Animal behaviour: Cognitive bias and affective state. Nature 2004, 427, 312. [Google Scholar] [CrossRef]
- Mendl, M.; Burman, O.H.P.; Parker, R.M.A.; Paul, E.S. Cognitive bias as an indicator of animal emotion and welfare: Emerging evidence and underlying mechanisms. Appl. Anim. Behav. Sci. 2009, 118, 161–181. [Google Scholar] [CrossRef]
- Kloke, V.; Schreiber, R.S.; Bodden, C.; Möllers, J.; Ruhmann, H.; Kaiser, S.; Lesch, K.-P.; Sachser, N.; Lewejohann, L. Hope for the Best or Prepare for the Worst? Towards a Spatial Cognitive Bias Test for Mice. PLoS ONE 2014, 9, e105431. [Google Scholar] [CrossRef]
- Richter, S.H.; Schick, A.; Hoyer, C.; Lankisch, K.; Gass, P.; Vollmayr, B. A glass full of optimism: Enrichment effects on cognitive bias in a rat model of depression. Cogn. Affect. Behav. Neurosci. 2012, 12, 527–542. [Google Scholar] [CrossRef] [PubMed]
- Wichman, A.; Keeling, L.J.; Forkman, B. Cognitive bias and anticipatory behaviour of laying hens housed in basic and enriched pens. Appl. Anim. Behav. Sci. 2012, 140, 62–69. [Google Scholar] [CrossRef]
- Doyle, R.E.; Lee, C.; Deiss, V.; Fisher, A.D.; Hinch, G.N.; Boissy, A. Measuring judgement bias and emotional reactivity in sheep following long-term exposure to unpredictable and aversive events. Physiol. Behav. 2011, 102, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Alcaro, A.; Panksepp, J. The SEEKING mind: Primal neuro-affective substrates for appetitive incentive states and their pathological dynamics in addictions and depression. Neurosci. Biobehav. Rev. 2011, 35, 1805–1820. [Google Scholar] [CrossRef]
- Van der Harst, J.E.; Baars, A.M.; Spruijt, B.M. Announced rewards counteract the impairment of anticipatory behaviour in socially stressed rats. Behav. Brain Res. 2005, 161, 183–189. [Google Scholar] [CrossRef]
- Destrez, A.; Deiss, V.; Leterrier, C.; Calandreau, L.; Boissy, A. Repeated exposure to positive events induces optimistic-like judgment and enhances fearfulness in chronically stressed sheep. Appl. Anim. Behav. Sci. 2014, 154, 30–38. [Google Scholar] [CrossRef]
- Balsam, P.; Sanchez-Castillo, H.; Taylor, K.; Van Volkinburg, H.; Ward, R.D. Timing and anticipation: conceptual and methodological approaches. Eur. J. Neurosci. 2009, 30, 1749–1755. [Google Scholar] [CrossRef] [Green Version]
- Panksepp, J.; Moskal, J. Dopamine and SEEKING: Subcortical “Reward” Systems and Appetitive Urges. In Handbook of Approach and Avoidance Motivation; Elliot, A.J., Ed.; Psychology Press: New York, NY, USA, 2008; pp. 67–87. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watters, J.V.; Krebs, B.L. Assessing and Enhancing the Welfare of Animals with Equivocal and Reliable Cues. Animals 2019, 9, 680. https://doi.org/10.3390/ani9090680
Watters JV, Krebs BL. Assessing and Enhancing the Welfare of Animals with Equivocal and Reliable Cues. Animals. 2019; 9(9):680. https://doi.org/10.3390/ani9090680
Chicago/Turabian StyleWatters, Jason V., and Bethany L. Krebs. 2019. "Assessing and Enhancing the Welfare of Animals with Equivocal and Reliable Cues" Animals 9, no. 9: 680. https://doi.org/10.3390/ani9090680
APA StyleWatters, J. V., & Krebs, B. L. (2019). Assessing and Enhancing the Welfare of Animals with Equivocal and Reliable Cues. Animals, 9(9), 680. https://doi.org/10.3390/ani9090680