
Thinking about the Biodiversity Loss in This Changing World


Abstract
:1. Introduction
“We live in a zoologically impoverished world, from which all the hugest, and fiercest, and strangest forms have recently disappeared.”Wallace, 1876
2. What about the Biodiversity Fluctuation during the Quaternary?
3. Are We on the Edge of a Sixth Mass Extinction?
4. Reflections on Contemporary Ecosystem Modifications and Fauna Depletion
“The more we exploit nature, the more our options are reduced, until we only have one: to fight for survival.”Morris K. Udall
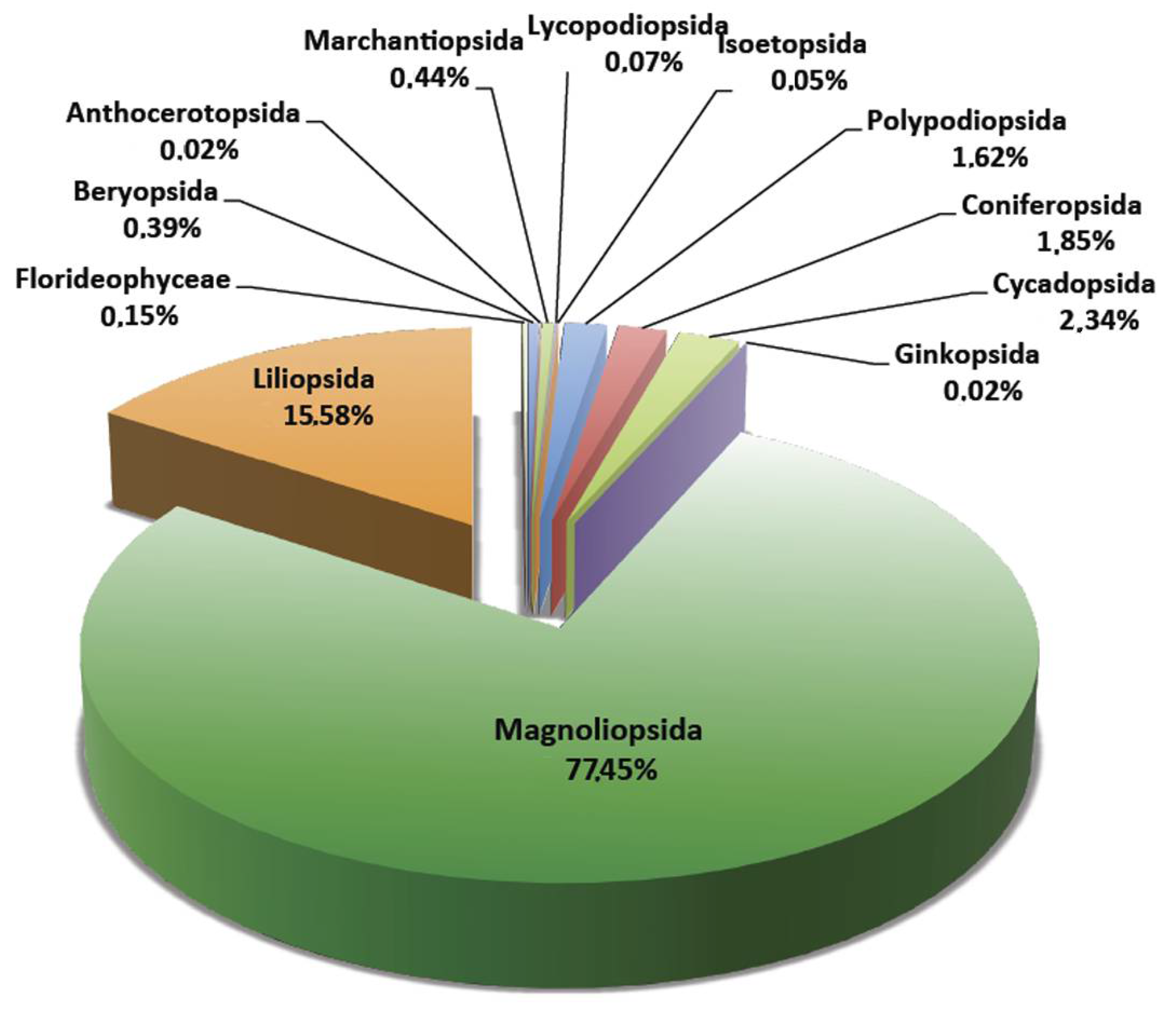
5. Remarks
“only about 1.5 species for the last 500 years and....these losses have probably been equalled or surpassed by species born (speciation) during that time”[122] (p. 243).
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leakey, R.E.; Lewin, R. The Sixth Extinction: Biodiversity and Its Survival; Phoenix Publishers: Phoenix, AZ, USA, 1995; pp. 1–271. [Google Scholar]
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.; Swartz, B.; Quental, T.B.; Marshall, C.; McGuire, J.L.; Lindsey, E.L.; Maguire, K.C.; et al. Has the Earth’s sixth mass extinction already arrived? Nature 2011, 471, 51–57. [Google Scholar] [CrossRef]
- Payne, J.L.; Bush, A.M.; Heim, N.A.; Knope, M.L.; McCauley, D.J. Ecological selectivity of the emerging mass extinction in the oceans. Science 2016, 353, 1284–1286. [Google Scholar] [CrossRef] [Green Version]
- Ceballos, G.; Ehrlich, P.R.; Raven, P.H. Vertebrates on the brink as indicators of biological annihilation and the sixth mass extinction. Proc. Natl. Acad. Sci. USA 2020, 117, 13596–13602. [Google Scholar] [CrossRef]
- Novacek, M.J. The Biodiversity Crisis: Losing What Counts; New Press: New York, NY, USA, 2001; pp. 1–223. [Google Scholar]
- Jablonski, D. Extinction: Past and present. Nature 2004, 427, 589. [Google Scholar] [CrossRef]
- Jablonski, D. Extinctions in the fossil record. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1994, 344, 11–17. [Google Scholar] [CrossRef]
- Bambach, R.K. Phanerozoic biodiversity mass extinctions. Annu. Rev. Earth Planet. Sci. 2006, 34, 127–155. [Google Scholar] [CrossRef]
- Sepkoski, J.J., Jr. Patterns of Phanerozoic Extinction: A Perspective from Global Data Bases. In Global Events and Event Stratigraphy in the Phanerozoic; Walliser, P.D.O.H., Ed.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 35–51. [Google Scholar]
- Wignall, P.B. Extinction: A Very Short Introduction; Oxford University Press: Oxford, UK, 2019; 125p. [Google Scholar]
- Elewa, A.M.T.; Abdelhady, A.A. Past, present, and future mass extinctions. J. Afr. Earth Sci. 2020, 162, 103678. [Google Scholar] [CrossRef]
- Plotnick, R.E.; Smith, F.A.; Lyons, S.K. The fossil record of the sixth extinction. Ecol. Lett. 2016, 19, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, J.W.; Weil, A. Delayed biological recovery from extinctions throughout the fossil record. Nature 2000, 404, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Erwin, D.H. Lessons from the past: Biotic recoveries from mass extinctions. Proc. Natl. Acad. Sci. USA 2001, 98, 1399–1403. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, L.W.; Alvarez, W.; Asaro, F.; Michel, H.V. Extraterrestrial cause for the Cretaceous-Tertiary extinction. Science 1980, 208, 1095–1108. [Google Scholar] [CrossRef] [Green Version]
- Raup, D.M.; Sepkoski, J.J., Jr. Periodic extinction of families and genera. Science 1986, 231, 833–836. [Google Scholar] [CrossRef]
- Erwin, D.H. The Permo–Triassic extinction. Nature 1994, 367, 231–236. [Google Scholar] [CrossRef]
- Wooldridge, S.A. Mass extinctions past and present: A unifying hypothesis. Biogeosci. Discuss. 2008, 5, 2401–2423. [Google Scholar] [CrossRef] [Green Version]
- Kaiho, K.; Kajiwara, Y.; Nakano, T.; Miura, Y.; Kawahata, H.; Tazaki, K.; Ueshima, M.; Chen, Z.; Shi, G.R. End-Permian catastrophe by a bolide impact: Evidence of a gigantic release of sulfur from the mantle. Geology 2001, 29, 815–818. [Google Scholar] [CrossRef]
- Yin, H.; Feng, Q.; Baud, A.; Xie, S.; Benton, M.J.; Lai, X.; Bottjer, D.J. The prelude of the end-Permian mass extinction predates a postulated bolide impact. Int. J. Earth Sci. 2007, 96, 903–909. [Google Scholar] [CrossRef]
- Blackburn, T.J.; Olsen, P.E.; Bowring, S.A.; McLean, N.M.; Kent, D.V.; Puffer, J.; McHone, G.; Rasbury, E.T.; Et-Touhami, M. Zircon U-Pb geochronology links the end-Triassic extinction with the Central Atlantic Magmatic Province. Science 2013, 340, 941–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoene, B.; Samperton, K.M.; Eddy, M.P.; Keller, G.; Adatte, T.; Bowring, S.A.; Khadri, S.F.R.; Gertsch, B. U-Pb geochronology of the Deccan Traps and relation to the end-Cretaceous mass extinction. Science 2015, 347, 182–184. [Google Scholar] [CrossRef] [PubMed]
- Ruban, D.A. Paleozoic–Mesozoic Eustatic Changes and Mass Extinctions: New Insights from Event Interpretation. Life 2020, 10, 281. [Google Scholar] [CrossRef]
- Bond, D.P.; Grasby, S.E. Late Ordovician mass extinction caused by volcanism, warming, and anoxia, not cooling and glaciation. Geology 2020, 48, 777–781. [Google Scholar] [CrossRef]
- Li, G.; Liao, W.; Li, S.; Wang, Y.; Lai, Z. Different triggers for the two pulses of mass extinction across the Permian and Triassic boundary. Sci. Rep. 2021, 11, 6686. [Google Scholar] [CrossRef] [PubMed]
- Kaiho, K.; Aftabuzzaman, M.; Jones, D.S.; Tian, L. Pulsed volcanic combustion events coincident with the end-Permian terrestrial disturbance and the following global crisis. Geology 2021, 49, 289–293. [Google Scholar] [CrossRef]
- Chiarenza, A.A.; Farnsworth, A.; Mannion, P.D.; Lunt, D.J.; Valdes, P.J.; Morgan, J.V.; Allison, P.A. Asteroid impact, not volcanism, caused the end-Cretaceous dinosaur extinction. Proc. Natl. Acad. Sci. USA 2020, 117, 17084–17093. [Google Scholar] [CrossRef]
- Dzombak, R.M.; Sheldon, N.D.; Mohabey, D.M.; Samant, B. Stable climate in India during Deccan volcanism suggests limited influence on K–Pg extinction. Gondwana Res. 2020, 85, 19–31. [Google Scholar] [CrossRef]
- Lyons, S.L.; Karp, A.T.; Bralower, T.J.; Grice, K.; Schaefer, B.; Gulick, S.P.; Morgan, J.V.; Freeman, K.H. Organic matter from the Chicxulub crater exacerbated the K–Pg impact winter. Proc. Natl. Acad. Sci. USA 2020, 117, 25327–25334. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Li, M.; Chen, J.; Luo, Q.; Grasby, S.E.; Zhang, T.; Yuan, S.; Xu, Y.; Finney, S.C.; Sun, Y.; et al. Major volcanic eruptions linked to the Late Ordovician mass extinction: Evidence from mercury enrichment and Hg isotopes. Glob. Planet. Chang. 2021, 196, 103374. [Google Scholar] [CrossRef]
- Cooper, R.A.; Sadler, P.M.; Munnecke, A.; Crampton, J.S. Graptoloid evolutionary rates track Ordovician–Silurian global climate change. Geol. Mag. 2014, 151, 349–364. [Google Scholar] [CrossRef] [Green Version]
- Kaiho, K.; Miura, M.; Tezuka, M.; Hayashi, N.; Jones, D.S.; Oikawa, K.; Casier, J.-G.; Fujibayashi, M.; Chen, Z.-Q. Coronene, mercury, and biomarker data support a link between extinction magnitude and volcanic intensity in the Late Devonian. Glob. Planet. Chang. 2021, 199, 103452. [Google Scholar] [CrossRef]
- Rakociński, M.; Marynowski, L.; Pisarzowska, A.; Bełdowski, J.; Siedlewicz, G.; Zatoń, M.; Perri, M.C.; Spalletta, C.; Schönlaub, H.P. Volcanic related methylmercury poisoning as the possible driver of the end-Devonian Mass Extinction. Sci. Rep. 2020, 10, 7344. [Google Scholar] [CrossRef]
- Claeys, P.; Casier, J.; Margolis, S.V. Mircotektites and mass extinctions: Evidence for a Late Devonian asteroid impact. Science 1992, 257, 1102–1104. [Google Scholar] [CrossRef]
- Tobase, T.; Yoshiasa, A.; Komatsu, T.; Maekawa, T.; Hongu, H.; Okube, M.; Arima, H.; Sugiyama, K. Titanium local coordination environments in Cretaceous–Paleogene and Devonian–Carboniferous boundary sediments as a possible marker for large meteorite impact. Phys. Chem. Miner. 2019, 46, 675–685. [Google Scholar] [CrossRef]
- Marshall, J.E.; Lakin, J.; Troth, I.; Wallace-Johnson, S.M. UV-B radiation was the Devonian-Carboniferous boundary terrestrial extinction kill mechanism. Sci. Adv. 2020, 6, eaba0768. [Google Scholar] [CrossRef]
- He, T.; Dal Corso, J.; Newton, R.J.; Wignall, P.B.; Mills, B.J.; Todaro, S.; Di Stefano, P.; Turner, E.C.; Jamieson, R.A.; Randazzo, V.; et al. An enormous sulfur isotope excursion indicates marine anoxia during the end-Triassic mass extinction. Sci. Adv. 2020, 6, eabb6704. [Google Scholar] [CrossRef]
- Punekar, J.; Keller, G.; Adatte, T.; Khozyem, H.M.; Font, E.; Spangenberg, J. A multi-proxy approach to decode the end-cretaceous mass extinction. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 441, 116–136. [Google Scholar] [CrossRef] [Green Version]
- Font, E.; Adatte, T.; Sial, A.N.; Lacerda, L.D.; De Keller, G.; Punekar, J. Mercury Anomaly, Deccan Volcanism, and the end-Cretaceous mass extinction. Geology 2016, 44, 171–174. [Google Scholar] [CrossRef]
- Font, E.; Adatte, T.; Andrade, M.; Keller, G.; Bitchong, A.M.; Carvallo, C.; Ferreira, J.; Diogo, Z.; Mirão, J. Deccan volcanism induced high-stress environment during the Cretaceous–Paleogene transition at Zumaia, Spain: Evidence from magnetic, mineralogical and biostratigraphic records. Earth Planet. Sci. Lett. 2018, 484, 53–66. [Google Scholar] [CrossRef]
- Stehlin, H.G. Remarques sur les faunules de mammifères des couches éocènes etoligocènes du Bassin de Paris. Bull. Soc. Géol. Fr. 1910, 9, 488–520. [Google Scholar]
- Hooker, J.J.; Grimes, S.T.; Mattey, D.P.; Collinson, M.E.; Sheldon, N.D. Refined correlation of the UK late Eocene–early Oligocene Solent Group, and timing of its climate history. In The Late Eocene Earth: Hothouse, Icehouse and Impacts; Koeberl, C., Montanari, A., Eds.; Special Paper 452; Geological Society of America: Boulder, CO, USA, 2009; pp. 179–195. [Google Scholar]
- Van Dam, J.A.; Aziz, H.A.; Sierra, M.Á.Á.; Hilgen, F.J.; van den Hoek Ostende, L.W.; Lourens, L.J.; Laurens, L.J.; Mein, P.; van der Meulen, A.J.; Pelaez-Campomanes, P. Long-period astronomical forcing of mammal turnover. Nature 2006, 443, 687–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchetto-Zolet, A.C.; Pinheiro, F.; Salgueiro, F.; Palma-Silva, C. Phylogeographical patterns shed light on evolutionary process in South America. Mol. Ecol. 2013, 22, 1193–1213. [Google Scholar] [CrossRef] [PubMed]
- Bibi, F.; Kiessling, W. Continuous evolutionary change in Plio-Pleistocene mammals of eastern Africa. Proc. Natl. Acad. Sci. USA 2015, 112, 10623–10628. [Google Scholar] [CrossRef] [Green Version]
- Head, M.J.; Gibbard, P.L. Early–Middle Pleistocene transitions: Linking terrestrial and marine realms. Quat. Int. 2015, 389, 7–46. [Google Scholar] [CrossRef] [Green Version]
- Crampton, J.S.; Cody, R.D.; Levy, R.; Harwood, D.; McKay, R.; Naish, T.R. Southern Ocean phytoplankton turnover in response to stepwise Antarctic cooling over the past 15 million years. Proc. Natl. Acad. Sci. USA 2016, 113, 6868–6873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palombo, M.R. Faunal dynamics in SW Europe during the late Early Pleistocene: Palaeobiogeographical insights and biochronological issues. Comptes Rendus Palevol 2018, 17, 247–261. [Google Scholar] [CrossRef]
- Rossi, V.; Azzarone, M.; Capraro, L.; Faranda, C.; Ferretti, P.; Macrì, P.; Scarponi, D. Dynamics of benthic marine communities across the Early-Middle Pleistocene boundary in the Mediterranean region (Valle di Manche, Southern Italy): Biotic and stratigraphic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 495, 127–138. [Google Scholar] [CrossRef]
- Suc, J.P.; Popescu, S.M.; Fauquette, S.; Bessedik, M.; Jiménez-Moreno, G.; Taoufiq, B.; Zheng, Z.; Médal, F.; Klotz, S. Reconstruction of Mediterranean flora, vegetation and climate for the last 23 million years based on an extensive pollen dataset. Ecol. Medit. 2018, 44, 53–85. [Google Scholar] [CrossRef]
- Zhou, X.; Yang, J.; Wang, S.; Xiao, G.; Zhao, K.; Zheng, Y.; Shen, H.; Li, X. Vegetation change and evolutionary response of large mammal fauna during the Mid-Pleistocene Transition in temperate northern East Asia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 505, 287–294. [Google Scholar] [CrossRef]
- Rawlence, N.J.; Scofield, R.P.; McGlone, M.S.; Knapp, M. History repeats: Large scale synchronous biological turnover in avifauna from the Plio-Pleistocene and Late Holocene of New Zealand. Front. Ecol. Evol. 2019, 7, 158. [Google Scholar] [CrossRef] [Green Version]
- Baker, P.A.; Fritz, S.C.; Battisti, D.S.; Dick, C.W.; Vargas, O.M.; Asner, G.P.; Martin, R.E.; Wheatley, A.; Prates, I. Beyond Refugia: New insights on Quaternary climate variation and the evolution of biotic diversity in tropical South America. In Neotropical Diversification: Patterns and Processes; Rull, V., Carnaval, A.C., Eds.; Springer: Cham, Switzerland, 2020; pp. 51–70. [Google Scholar]
- Diz, P.; Peñalver-Clavel, I.; Hernández-Almeida, I.; Bernasconi, S.M. Environmental changes in the East Equatorial Pacific during the Mid Pleistocene Transition and implications for the Last Global Extinction of benthic foraminifera. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 539, 109487. [Google Scholar] [CrossRef]
- Magri, D.; Parra, I.; Di Rita, F.; Ni, J.; Shichi, K.; Worth, J.R. Linking worldwide past and present conifer vulnerability. Quat. Sci. Rev. 2020, 250, 106640. [Google Scholar] [CrossRef]
- O’Brien, K.; Patterson, D.B.; Biernat, M.D.; Braun, D.R.; Cerling, T.E.; McGrosky, A.; Faith, J.T. Ungulate turnover in the Koobi Fora Formation: Spatial and temporal variation in the Early Pleistocene. J. Afr. Earth Sci. 2020, 161, 103658. [Google Scholar] [CrossRef]
- Pérez, L.; Correa-Metrio, A.; Cohuo, S.; González, L.M.; Echeverría-Galindo, P.; Brenner, M.; Curtis, J.; Kutterofl, S.; Stockhecke, M.; Schenk, F.; et al. Ecological turnover in Neotropical freshwater and terrestrial communities during episodes of abrupt climate change. Quat. Res. 2020, 101, 26–36. [Google Scholar] [CrossRef]
- Pinsky, M.L.; Selden, R.L.; Kitchel, Z.J. Climate-driven shifts in marine species ranges: Scaling from organisms to communities. Ann. Rev. Mar. Sci. 2020, 12, 153–179. [Google Scholar] [CrossRef] [PubMed]
- Couvreur, T.L.; Dauby, G.; Blach-Overgaard, A.; Deblauwe, V.; Dessein, S.; Droissart, V.; Hardy, O.J.; Harris, D.J.; Janssens, S.B.; Ley, A.C.; et al. Tectonics, climate and the diversification of the tropical African terrestrial flora and fauna. Biol. Rev. 2021, 96, 16–51. [Google Scholar] [CrossRef] [PubMed]
- Koch, P.L.; Barnosky, A.D. Late Quaternary extinctions: State of the debate. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 215–250. [Google Scholar] [CrossRef] [Green Version]
- Surovell, T.A.; Grund, B.S. The associational critique of Quaternary overkill and why it is largely irrelevant to the extinction debate. Am. Antiq. 2012, 77, 672–687. [Google Scholar] [CrossRef] [Green Version]
- Saltré, F.; Rodríguez-Rey, M.; Brook, B.W.; Johnson, C.N.; Turney, C.S.; Alroy, J.; Cooper, A.; Beeton, N.; Bird, M.I.; Fordham, D.A.; et al. Climate change not to blame for late Quaternary megafauna extinctions in Australia. Nat. Commun. 2016, 7, 10511. [Google Scholar] [CrossRef] [Green Version]
- Lima-Ribeiro, M.S.; Diniz-Filho, J.A.F. Climate change, human overkill, and the extinction of megafauna: A macroecological approach based on pattern-oriented modelling. Evol. Ecol. Res. 2017, 18, 97–121. [Google Scholar]
- Van Der Kaars, S.; Miller, G.H.; Turney, C.S.; Cook, E.J.; Nürnberg, D.; Schönfeld, J.; Kershaw, A.P.; Lehman, S.J. Humans rather than climate the primary cause of Pleistocene megafaunal extinction in Australia. Nat. Commun. 2017, 8, 14142. [Google Scholar] [CrossRef] [Green Version]
- Broughton, J.M.; Weitzel, E.M. Population reconstructions for humans and megafauna suggest mixed causes for North American Pleistocene extinctions. Nat. Commun. 2018, 9, 5441. [Google Scholar] [CrossRef]
- Nagaoka, L.; Rick, T.; Wolverton, S. The overkill model and its impact on environmental research. Ecol. Evol. 2018, 8, 9683–9696. [Google Scholar] [CrossRef]
- Price, G.J.; Louys, J.; Faith, J.T.; Lorenzen, E.; Westaway, M.C. Big data little help in megafauna mysteries. Nature 2018, 558, 23–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hocknull, S.A.; Lewis, R.; Arnold, L.J.; Pietsch, T.; Joannes-Boyau, R.; Price, G.J.; Moss, P.; Wood, R.; Dosseto, A.; Louys, J.; et al. Extinction of eastern Sahul megafauna coincides with sustained environmental deterioration. Nat. Comm. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jukar, A.M.; Lyons, S.K.; Wagner, P.J.; Uhen, M.D. Late Quaternary extinctions in the Indian Subcontinent. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 562, 110137. [Google Scholar] [CrossRef]
- Lopes, R.P.; Pereira, J.C.; Kerber, L.; Dillenburg, S.R. The extinction of the Pleistocene megafauna in the Pampa of southern Brazil. Quat. Sc. Rev. 2020, 242, 106428. [Google Scholar] [CrossRef]
- Seersholm, F.V.; Werndly, D.J.; Grealy, A.; Johnson, T.; Early, E.M.K.; Lundelius, E.L.; Winsborough, B.; Early Farr, G.; Toomey, R.; Anders, A.J.; et al. Rapid range shifts and megafaunal extinctions associated with late Pleistocene climate change. Nat. Commun. 2020, 11, 2770. [Google Scholar] [CrossRef]
- Stewart, M.; Carleton, W.C.; Groucutt, H.S. Climate change, not human population growth, correlates with Late Quaternary megafauna declines in North America. Nat. Commun. 2021, 12, 965. [Google Scholar] [CrossRef] [PubMed]
- Turner, S. Progress in the Formalisation of the Anthropocene GSSP. EGU General Assembly Conference Abstracts. held online 19–30 April, 2021, id.EGU21-3226 2021; Bibcode EGU21-3226. Available online: https://ui.adsabs.harvard.edu/abs/2021EGUGA..23.3226T (accessed on 19 May 2021).
- Zalasiewicz, J.; Waters, C.N.; Ellis, E.C.; Head, M.J.; Vidas, D.; Steffen, W.; Thomas, J.A.; Horn, E.; Summerhayes, C.P.; Leinfelder, R.; et al. The Anthropocene: Comparing its meaning in geology (chronostratigraphy) with conceptual approaches arising in other disciplines. Earth’s Future 2021, 9, e2020EF001896. [Google Scholar] [CrossRef]
- Braje, T.J.; Erlandson, J.M. Human acceleration of animal and plant extinctions: A Late Pleistocene, Holocene, and Anthropocene continuum. Anthropocene 2013, 4, 14–23. [Google Scholar] [CrossRef]
- Pimm, S.L.; Raven, P. Extinction by numbers. Nature 2000, 403, 843–845. [Google Scholar] [CrossRef]
- May, R.M. The future of biological diversity in a crowded world. Curr. Sci. 2002, 82, 1325–1331. [Google Scholar]
- Mace, G.M.; Collar, N.J.; Gaston, K.J.; Hilton-Taylor, C.; Akçakaya, H.R.; Leader-Williams, N.; Milner-Gulland, E.J.; Stuart, S.N. Quantification of extinction risk: IUCN’s system for classifying threatened species. Conserv. Biol. 2008, 22, 1424–1442. [Google Scholar] [CrossRef] [PubMed]
- Vié, J.C.; Hilton-Taylor, C.; Pollock, C.; Ragle, J.; Smart, J.; Stuart, S.N.; Tong, R. The IUCN Red List: A Key Conservation Tool. Wildlife in a Changing World: An Analysis of the 2008 IUCN Red List of Threatened Species; IUNC: Gland, Switzerland, 2009; pp. 1–159. [Google Scholar]
- Newell, N.D. Adequacy of the fossil record. J. Paleontol. 1959, 33, 488–499. [Google Scholar]
- Naimark, E.; Kirpotin, D.; Boeva, N.; Gmoshinskiy, V.; Kalinina, M.; Lyupina, Y.; Markov, A.; Nikitin, M.; Shokurov, A.; Volkov, D. Taphonomic experiments imply a possible link between the evolution of multicellularity and the fossilization potential of soft-bodied organisms. Ecol. Evol. 2020, 11, 1037–1056. [Google Scholar] [CrossRef]
- Foote, M. Origination and extinction components of taxonomic diversity: General problems. Paleobiology 2000, 26, 74–102. [Google Scholar] [CrossRef]
- Behrensmeyer, A.K.; Hill, A.P. (Eds.) Fossils in the Making: Vertebrate Taphonomy and Paleoecology (No. 69); University of Chicago Press: Chicago, IL, USA, 1988; pp. 1–345. [Google Scholar]
- Lyman, R.L. Vertebrate Taphonomy; Cambridge University Press: Cambridge, UK, 1994; p. xxvi + 524. [Google Scholar]
- Pimm, S.L.; Russell, G.J.; Gittleman, J.L.; Brooks, T.M. The future of biodiversity. Science 1995, 269, 347–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeVos, J.M.; Joppa, L.N.; Gittleman, J.L.; Stephens, P.R.; Pimm, S.L. Estimating the normal background rate of species extinction. Conserv. Biol. 2014, 29, 452–462. [Google Scholar] [CrossRef] [Green Version]
- Plotnick, R.E.; Koy, K.A. The Anthropocene fossil record of terrestrial mammals. Anthropocene 2020, 29, 100233. [Google Scholar] [CrossRef]
- MA (Millennium Ecosystem Assessment). Millennium Ecosystem Assessment Synthesis Report; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- IPBES. Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; IPBES Secretariat: Bonn, Germany, 2019. [Google Scholar]
- Lamkin, M.; Miller, A.I. On the challenge of comparing contemporary and deep-time biological-extinction rates. BioScience 2016, 66, 785–789. [Google Scholar] [CrossRef]
- Pimm, S.L.; Brooks, T.M. The sixth extinction: How large, where and when? In Nature and Human Society: The Quest for a Sustainable World; Raven, P.H., Williams, T., Eds.; National Academy Press: Washington, DC, USA, 2000; pp. 46–62. [Google Scholar]
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef] [Green Version]
- IUCN. Summary Statistics. 2020. Available online: https://www.iucnredlist.org/resources/summary-statistics#Summary (accessed on 19 May 2021).
- WWF. Living Planet Report WWF (2020). In Living Planet Report 2020—Bending the Curve of Biodiversity Loss; Almond, R.E.A., Grooten, M., Petersen, T., Eds.; WWF: Gland, Switzerland, 2020; pp. 1–158. [Google Scholar]
- Mora, C.; Tittensor, D.P.; Adl, S.; Simpson, A.G.; Worm, B. How many species are there on Earth and in the ocean? PLoS Biol. 2019, 9, e1001127. [Google Scholar] [CrossRef] [Green Version]
- IUNC Sampled Red List Index for Plants. 2021. Available online: http://threatenedplants.myspecies.info (accessed on 19 May 2021).
- Brummitt, N.; Bachman, S.P.; Moat, J. Applications of the IUCN Red List: Towards a global barometer for plant diversity. Endanger. Species Res. 2008, 6, 127–135. [Google Scholar] [CrossRef]
- Bachman, S.P.; Field, R.; Reader, T.; Raimondo, D.; Donaldson, J.; Schatz, G.E.; Lughadha, E.N. Progress, challenges and opportunities for Red Listing. Biol. Conserv. 2019, 234, 45–55. [Google Scholar] [CrossRef]
- Pitman, N.C.; Jorgensen, P.M. Estimating the size of the world’s threatened Flora. Science 2002, 298, 986–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivers, M.; Beech, E.; Bazos, I.; Bogunić, F.; Buira, A.; Caković, D.; Carapeto, A.; Carta, A.; Cornier, B.; Fenu, G.; et al. European Red List of Trees; IUCN Red List of Threatened Species: Regional Assessment; International Union for Conservation of Nature and Natural Resources (IUCN): Cambridge, UK; Brussels, Belgium, 2019. [Google Scholar] [CrossRef] [Green Version]
- IUNC. Table 4b: Plant Species (Kingdom: Plantae) by Class and Family. 2020. Available online: https://www.iucnredlist.org/statistics (accessed on 19 May 2021).
- Brooks, D.R.; Hoberg, E.P.; Boeger, W.A. The Stockholm Paradigm: Climate Change and Emerging Disease; University of Chicago Press: Chicago, IL, USA, 2019; pp. 1–409. [Google Scholar]
- Rothman, D.H. Thresholds of catastrophe in the Earth system. Sci. Adv. 2017, 3, e1700906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botkin, D.B.; Saxe, H.; Araujo, M.B.; Betts, R.; Bradshaw, R.H.; Cedhagen, T.; Chesson, P.; Dawson, T.P.; Etterson, J.R.; Faith, D.P.; et al. Forecasting the effects of global warming on biodiversity. Bioscience 2007, 57, 227–236. [Google Scholar] [CrossRef]
- Combourieu-Nebout, N.; Bertini, A.; Russo-Ermolli, E.; Peyron, O.; Klotz, S.; Montade, V.; Fauquette, S.; Allen, J.; Fusco, F.; Goring, S.; et al. Climate changes in the central Mediterranean and Italian vegetation dy-namics since the Pliocene. Rev. Palaeobot. Palynol. 2015, 218, 127–147. [Google Scholar] [CrossRef]
- Zhou, Z.; Ding, Y.; Shi, H.; Cai, H.; Fu, Q.; Liu, S.; Li, T. Analysis and prediction of vegetation dynamic changes in China: Past, present and future. Ecol. Indic. 2020, 117, 106642. [Google Scholar] [CrossRef]
- Elias, S.A. 4 The Value of Insects in Paleoecology. Dev. Quat. Sci. 2010, 12, 39–51. [Google Scholar] [CrossRef]
- Alter, S.E.; Meyer, M.; Post, K.; Czechowski, P.; Gravlund, P.; Gaines, C.; Rosenbaum, H.C.; Kaschner, K.; Turvey, S.T.; van der Plicht, J.; et al. Climate impacts on transocean dispersal and habitat in gray whales from the Pleistocene to 2100. Mol. Ecol. 2015, 24, 1510–1522. [Google Scholar] [CrossRef]
- Baca, M.; Nadachowski, A.; Lipecki, G.; Mackiewicz, P.; Marciszak, A.; Popović, D.; Socha, P.; Stefaniak, K.; Wojtal, P. Impact of climatic changes in the Late Pleistocene on migrations and extinction of mammals in Europe: Four case studies. Geol. Q. 2017, 61, 291–304. [Google Scholar] [CrossRef]
- Barnosky, A.D. Transforming the global energy system is required to avoid the sixth mass extinction. MRS Energy Sustain. 2015, 2, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Schloss, C.A.; Nuñez, T.A.; Lawler, J.J. Dispersal will limit ability of mammals to track climate change in the Western Hemisphere. Proc. Natl. Acad. Sci. USA 2012, 109, 8606–8611. [Google Scholar] [CrossRef] [Green Version]
- Pinsky, M.L.; Eikeset, A.M.; McCauley, D.J.; Payne, J.L.; Sunday, J.M. Greater vulnerability to warming of marine versus terrestrial ectotherms. Nature 2019, 569, 108–111. [Google Scholar] [CrossRef]
- Young, H.S.; McCauley, D.J.; Galetti, M.; Dirzo, R. Patterns, Causes, and Consequences of Anthropocene Defaunation. Ann. Rev. Ecol. Evol. Syst. 2016, 47, 333–358. [Google Scholar] [CrossRef] [Green Version]
- Borja, A.; Andersen, J.H.; Arvanitidis, C.D.; Basset, A.; Buhl-Mortensen, L.; Carvalho, S.; Dafforn, C.D.; Devlin, M.J.; Escobar-Briones, E.G.; Grenz, C.; et al. Past and future grand challenges in marine ecosystem ecology. Front. Mar. Sci. 2020, 7, 362. [Google Scholar] [CrossRef]
- Kirkman, S.P.; Baliwe, N.G.; Nhleko, J.; Pfaff, M.C. Ecosystem health and human wealth—A comparison of sub-Saharan African Large Marine Ecosystems. Environ. Dev. 2020, 36, 100551. [Google Scholar] [CrossRef]
- Scholes, R.J.; Biggs, R. A biodiversity intactness index. Nature 2005, 434, 45–49. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.N.; Arnell, A.P.; Contu, S.; De Palma, A.; Ferrier, S.; Hill, S.L.L.; Hoskins, A.J.; Lysenko, I.; Phillips, H.R.; et al. Has land use pushed terrestrial biodiversity beyond the planetary boundary? A global assessment. Science 2016, 353, 288–291. [Google Scholar] [CrossRef]
- Díaz, S.; Settele, J.; Brondízio, E.S.; Ngo, H.T.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.A.; Butchart, S.H.M.; Chan, K.M.A.; et al. Pervasive human-driven decline of life on Earth points to the need for transformative change. Science 2019, 366, eaax3100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IPBES. Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Díaz, S., Settele, J., Brondízio, E., Ngo, H., Guèze, M., Agard, J., Arneth, A., Balvanera, P., Brauman, K., Butchart, S., et al., Eds.; IPBES Secretariat: Bonn, Germany, 2019; pp. 1–56. [Google Scholar]
- Briggs, J.C. Emergence of a sixth mass extinction? Biol. J. Linn. Soc. 2017, 122, 243–248. [Google Scholar] [CrossRef]
- MacLeod, N. The Great Extinctions; Firefly Books: Buffalo, NY, USA, 2013; p. 208. [Google Scholar]
- Everard, M.; Johnston, P.; Santillo, D.; Staddon, C. The role of ecosystems in mitigation and management of Covid-19 and other zoonoses. Environ. Sci. Policy 2020, 111, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shipley, B.R.; Lauer, D.A.; Pineau, R.M.; McGuire, J.L. Plant biomes demonstrate that landscape resilience today is the lowest it has been since end-Pleistocene megafaunal extinctions. Glob. Chang. Biol. 2020, 26, 5914–5927. [Google Scholar] [CrossRef] [PubMed]
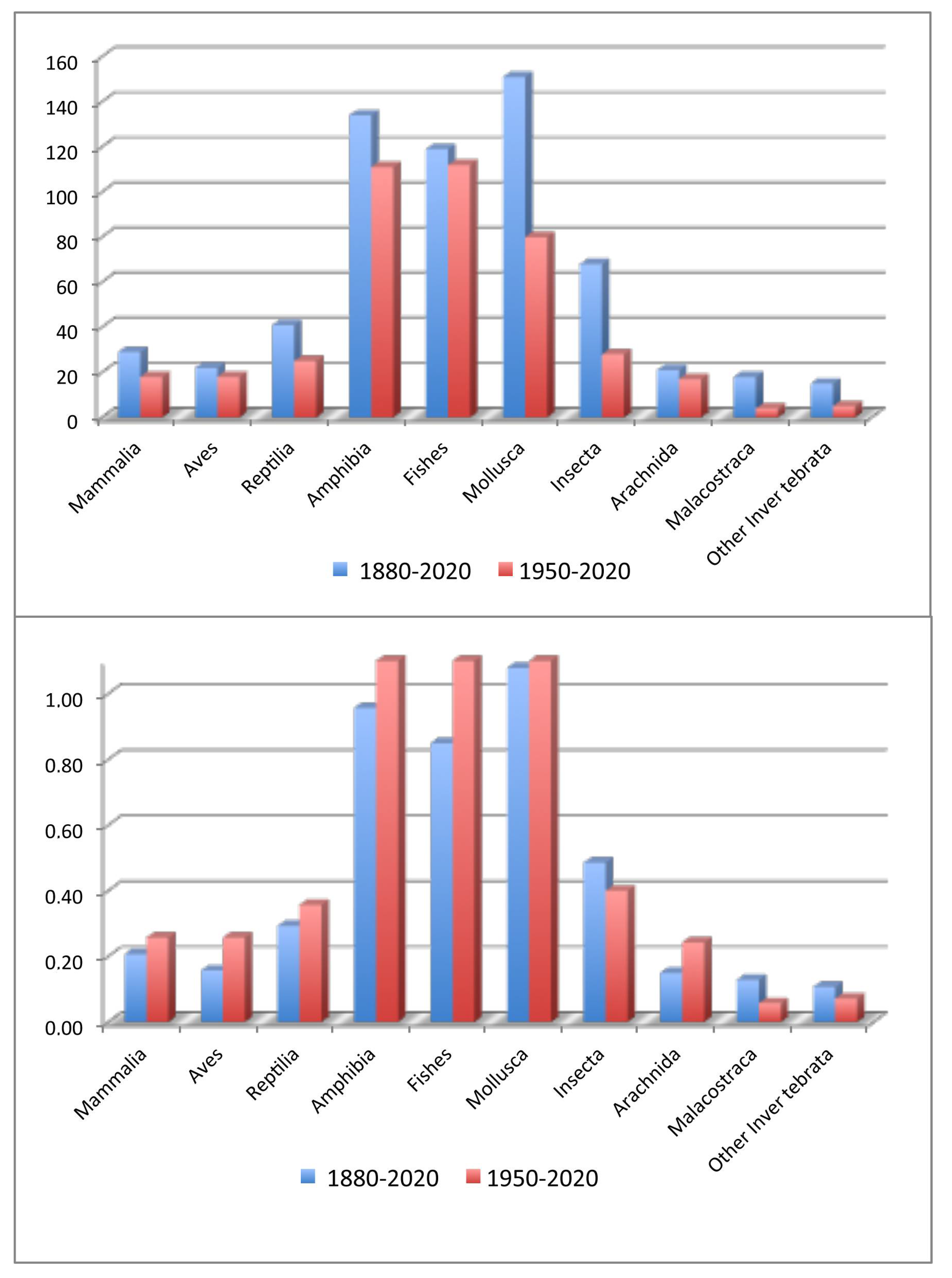
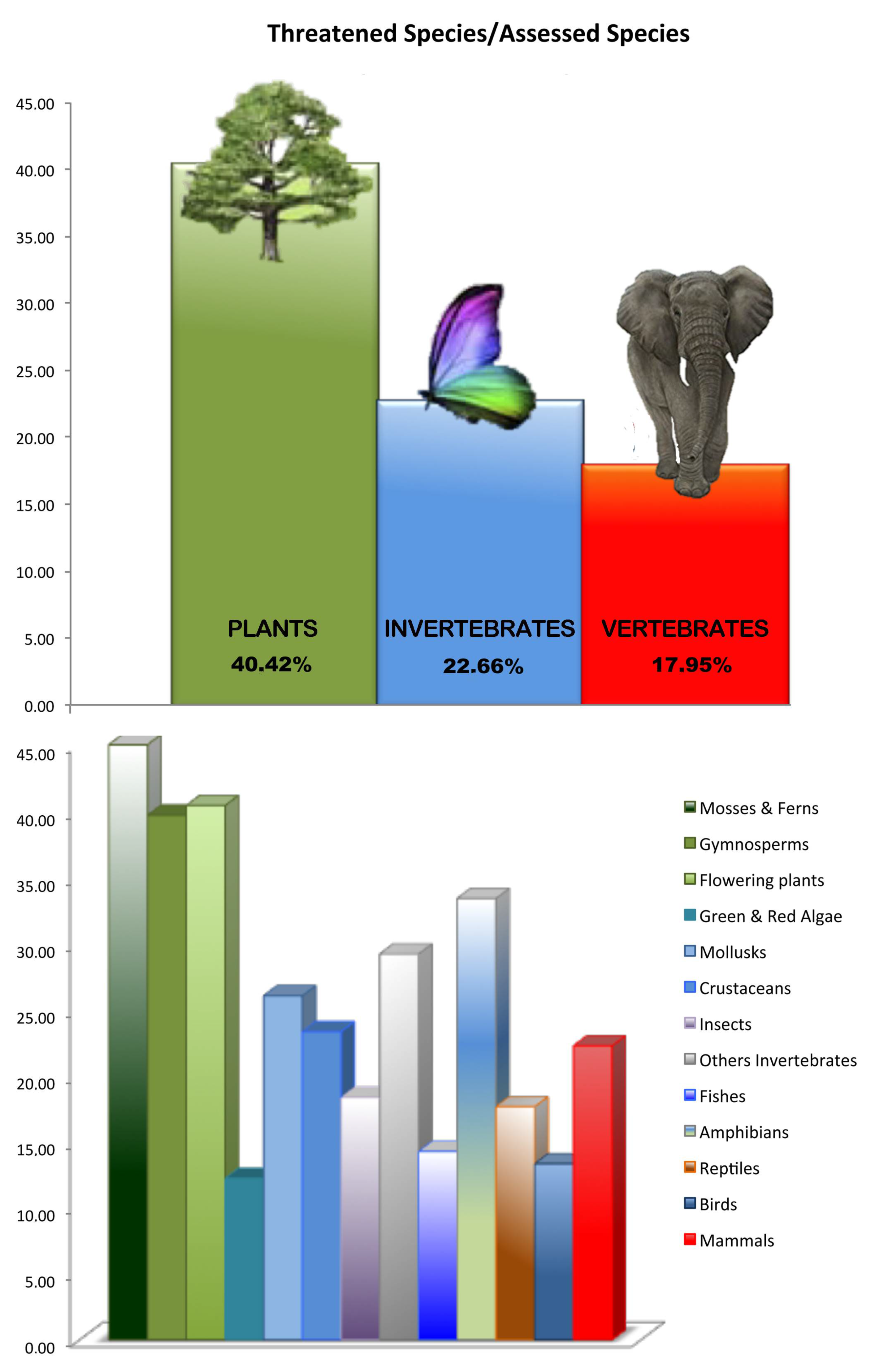

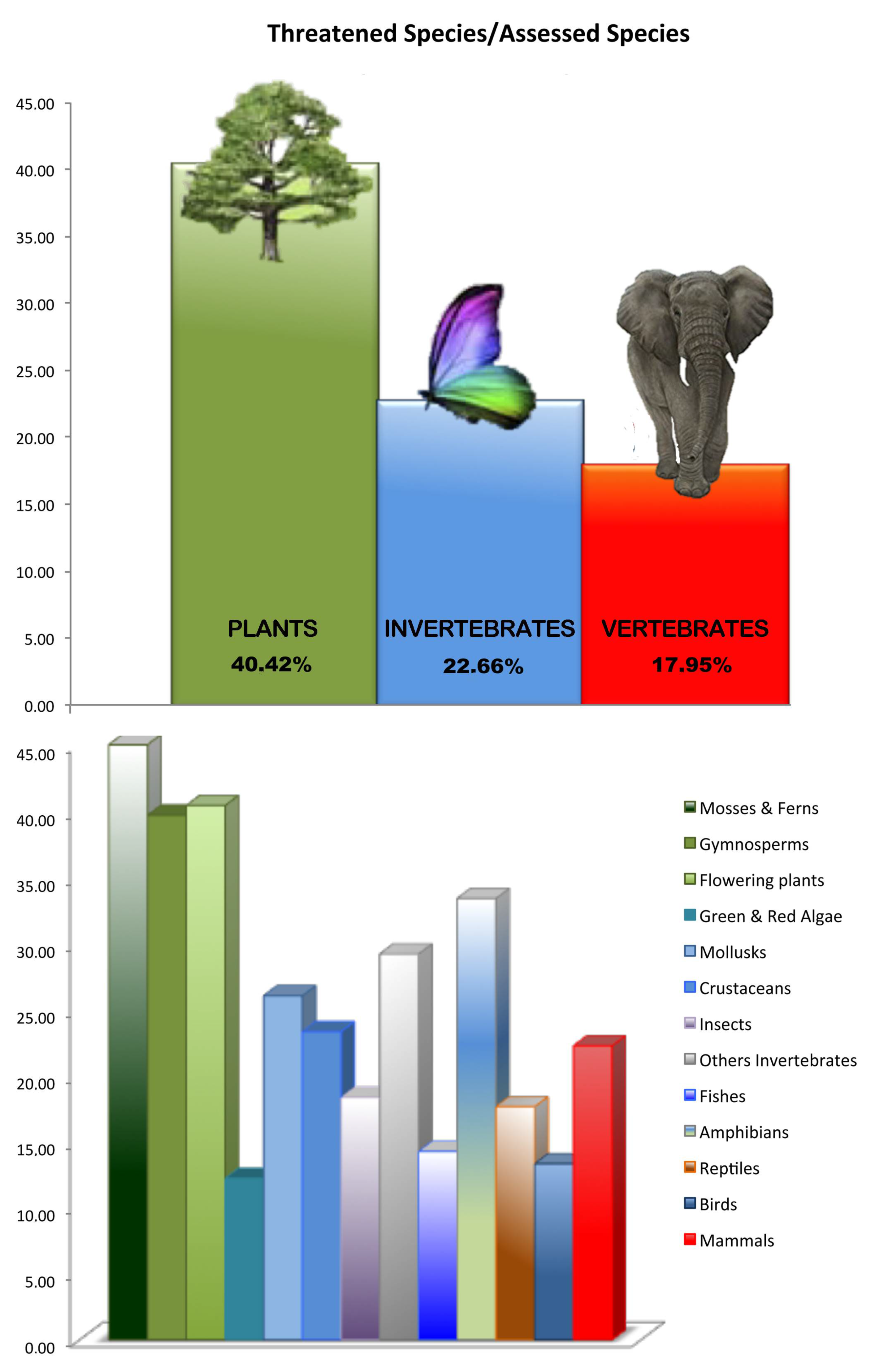
{kind=link}
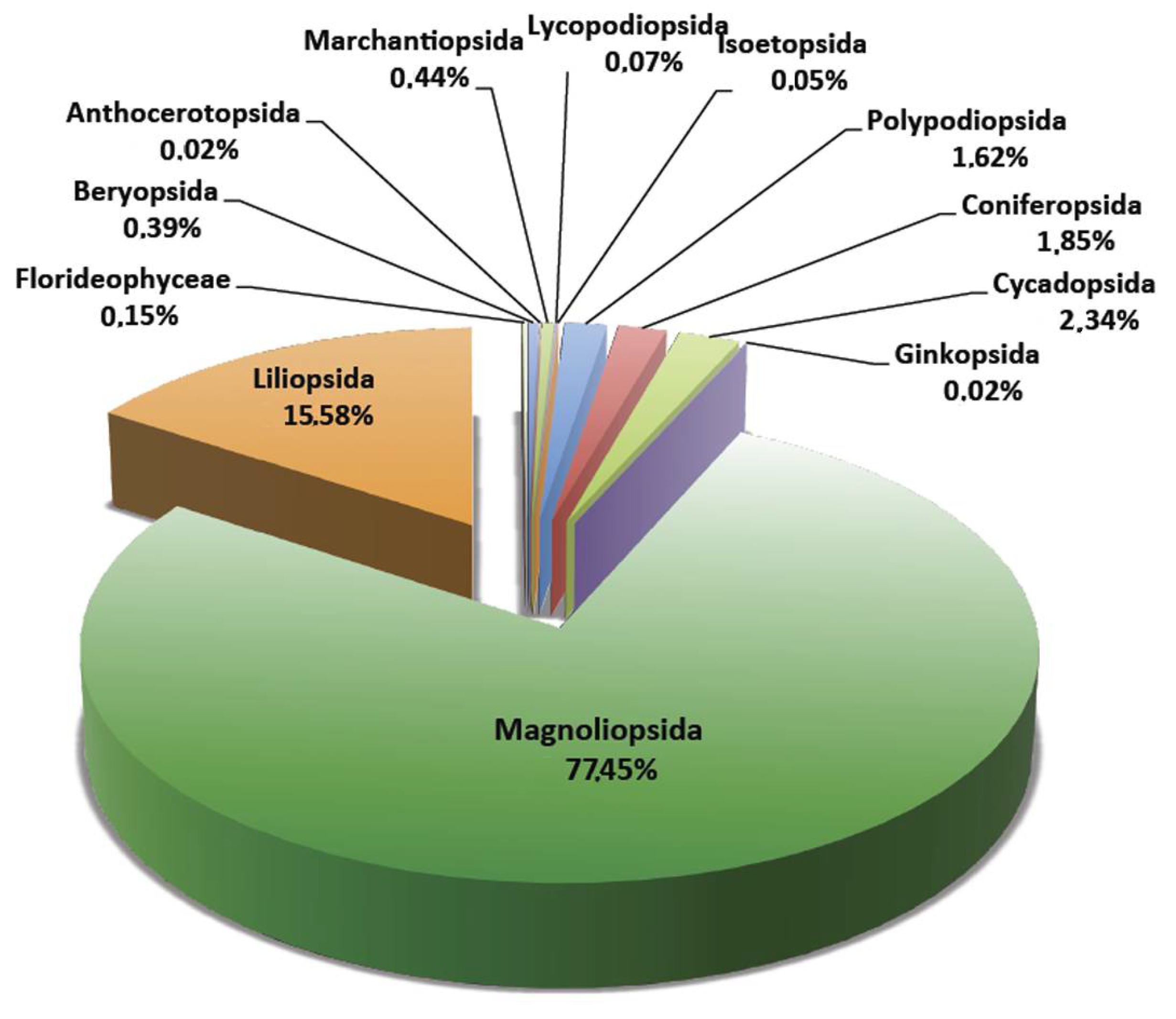
{kind=link}
{kind=link}
| Event | End (MYR Ago) | Extent * (Myr) | Genera Estimated Lost * | Species Estimated Lost * | Main Proposed Causes [2,10,11,16,17,18,21,22,23] | Other Proposed Causes |
|---|---|---|---|---|---|---|
| End-Ordovician | ~443 | 3.3–1.9 | 57% | 86% | Climate (alternating glacial and interglacial episodes) but [24]; sea level changes, regression followed by transgression phases, triggering ocean anoxia. | Volcanism revealed by high concentration of mercury traces, e.g., [30]; changes in atmospheric and oceanic geochemistry, e.g., [31];. |
| Late-Devonian | ~359 | 2.9–2 | 35% | 75% | Global cooling followed by global warming; atmospheric carbon dioxide decrease; sea level rise associated with the spreas of anoxic waters from depth shelf into shallow waters. | Volcanism revealed by high mercury concentration, e.g., [32], methylmercury poisoning [33]; meteorite impact, e.g., [34,35]; UV-B radiation [36]. |
| End-Permian | ~251 | 2.8–0.16 | 56% | 96% | Siberian volcanism leading to high hydrogen sulphide, carbon dioxide levels, and halogenous levels in the oceans and atmosphere, but see [25,26]; ocean acidification; global greenhouse warming. | Meteorite impact but [19,20] |
| end-Triassic (P-Tr) | ~200 | 8.3–0.6 | 47% | 80% | Increase in temperature likely caused by magmatic activity (Central Atlantic Magmatic Province) and resulting augment of atmospheric carbon dioxide; release of greenhouse gases causing deep-ocean acidification; ocean calcification crisis. | Ocean anoxia, e.g., [37] |
| End-Cretaceous (K-Pg) | ~65 | 2.5–less than 1 Myr | 40% | 76% | Long-term events: Deccan volcanism; carbon dioxide, inducing warming; tectonic uplift, accelerating erosion and contributing to ocean eutrophication and anoxic episodes. Rapid event: Chicxulub asteroid impact (Yucatàn Peninsula Mexico), causing sudden cooling and a prolonged cold winter. see also [27,28,29] | Ocean acidification and Hg toxicity, e.g., [38,39,40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palombo, M.R. Thinking about the Biodiversity Loss in This Changing World. Geosciences 2021, 11, 370. https://doi.org/10.3390/geosciences11090370
Palombo MR. Thinking about the Biodiversity Loss in This Changing World. Geosciences. 2021; 11(9):370. https://doi.org/10.3390/geosciences11090370
Chicago/Turabian StylePalombo, Maria Rita. 2021. "Thinking about the Biodiversity Loss in This Changing World" Geosciences 11, no. 9: 370. https://doi.org/10.3390/geosciences11090370
APA StylePalombo, M. R. (2021). Thinking about the Biodiversity Loss in This Changing World. Geosciences, 11(9), 370. https://doi.org/10.3390/geosciences11090370