
The Large Dendritic Morphologies in the Antoniadi Crater (Mars) and Their Potential Astrobiological Significance


Abstract
:1. Introduction
2. The Antoniadi Crater
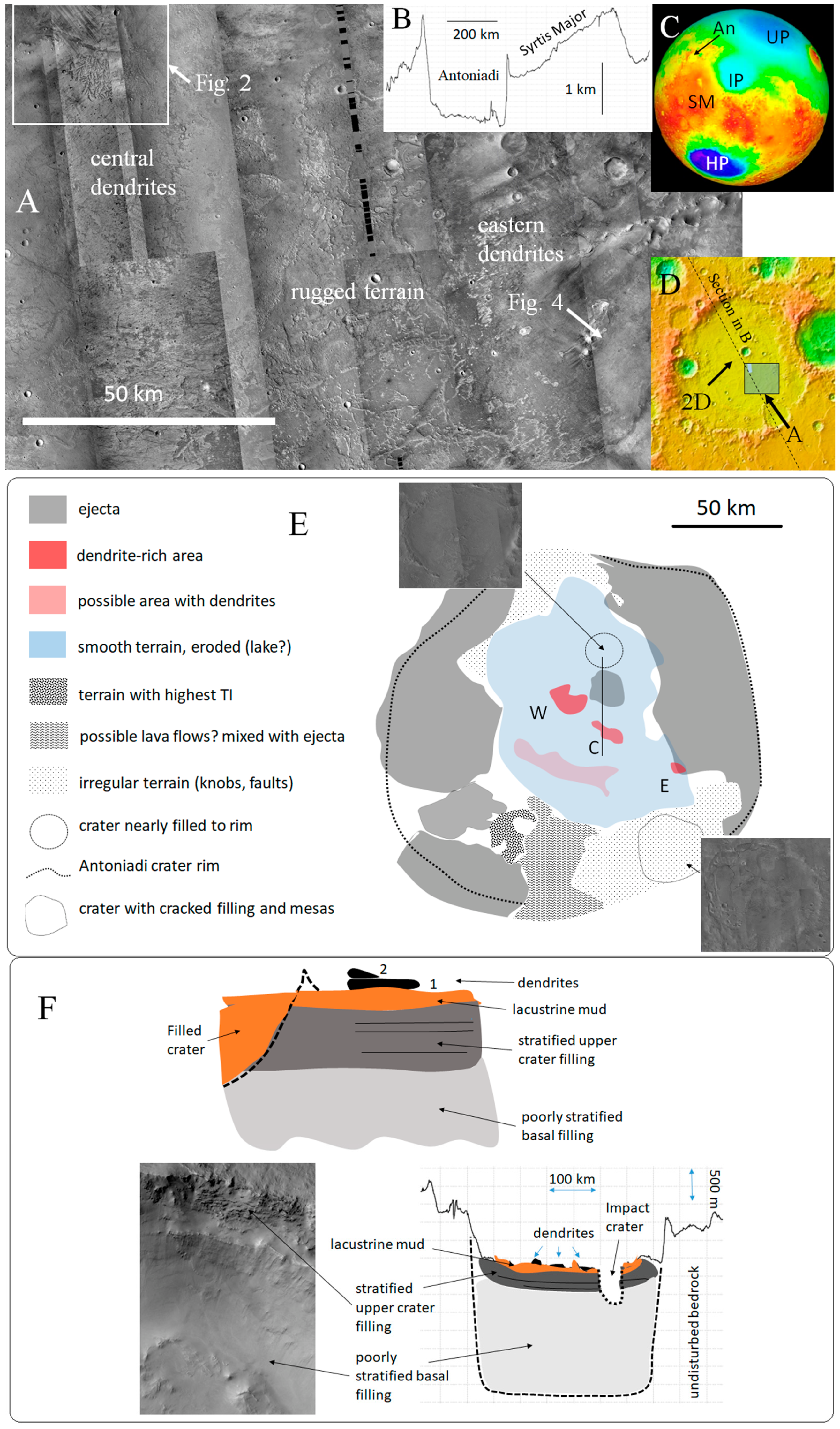
2.1. General Description
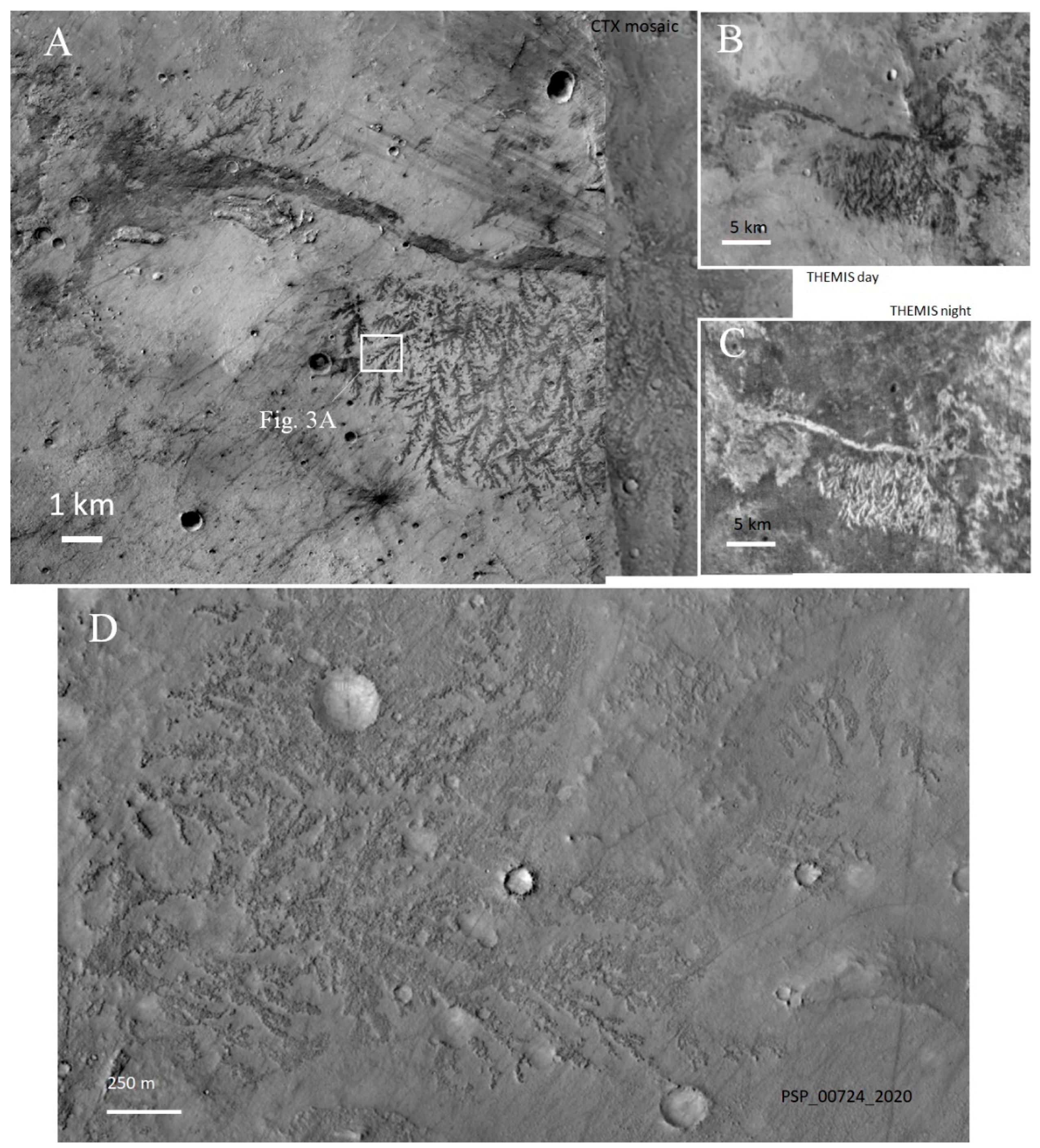
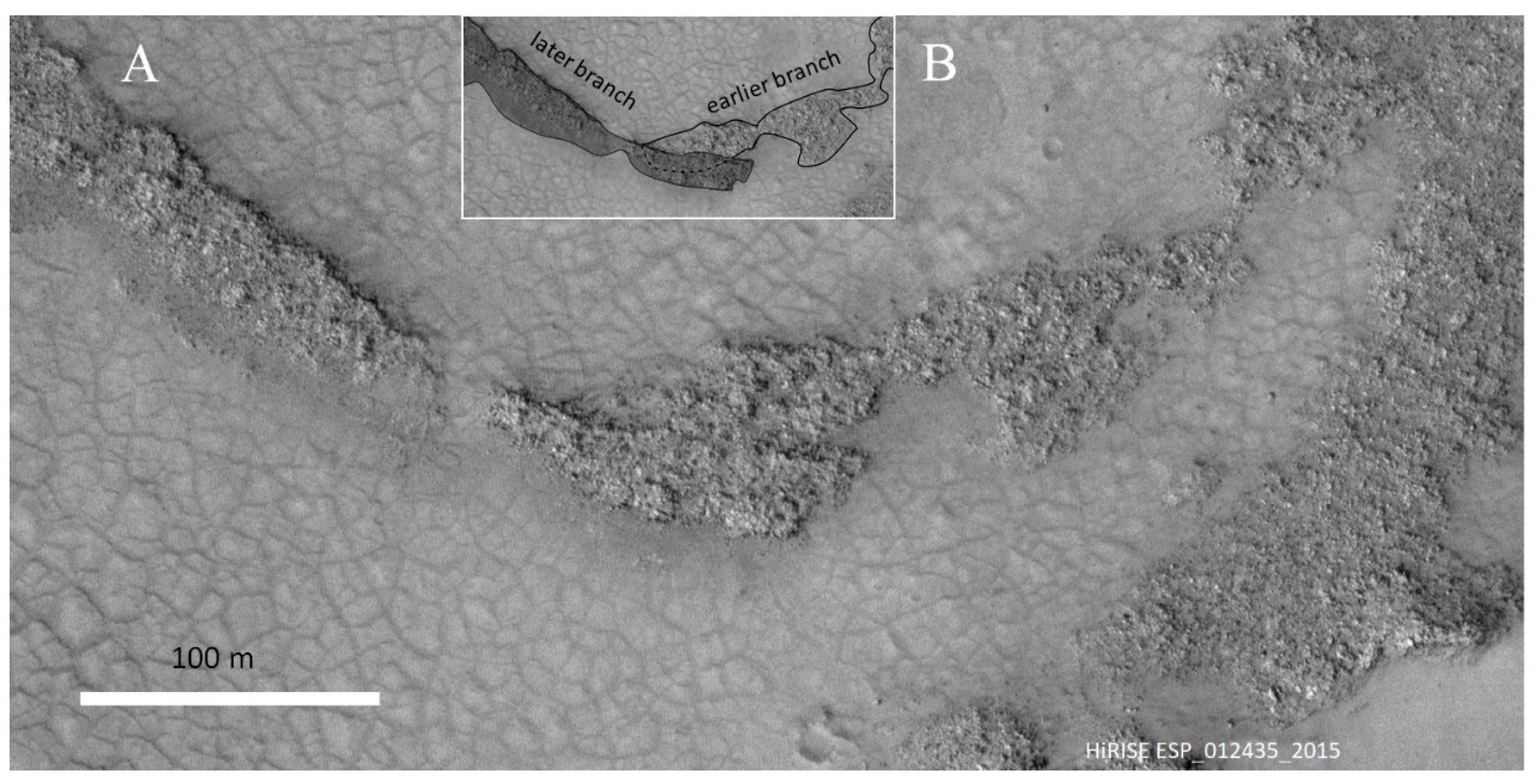
2.2. The Dendrites
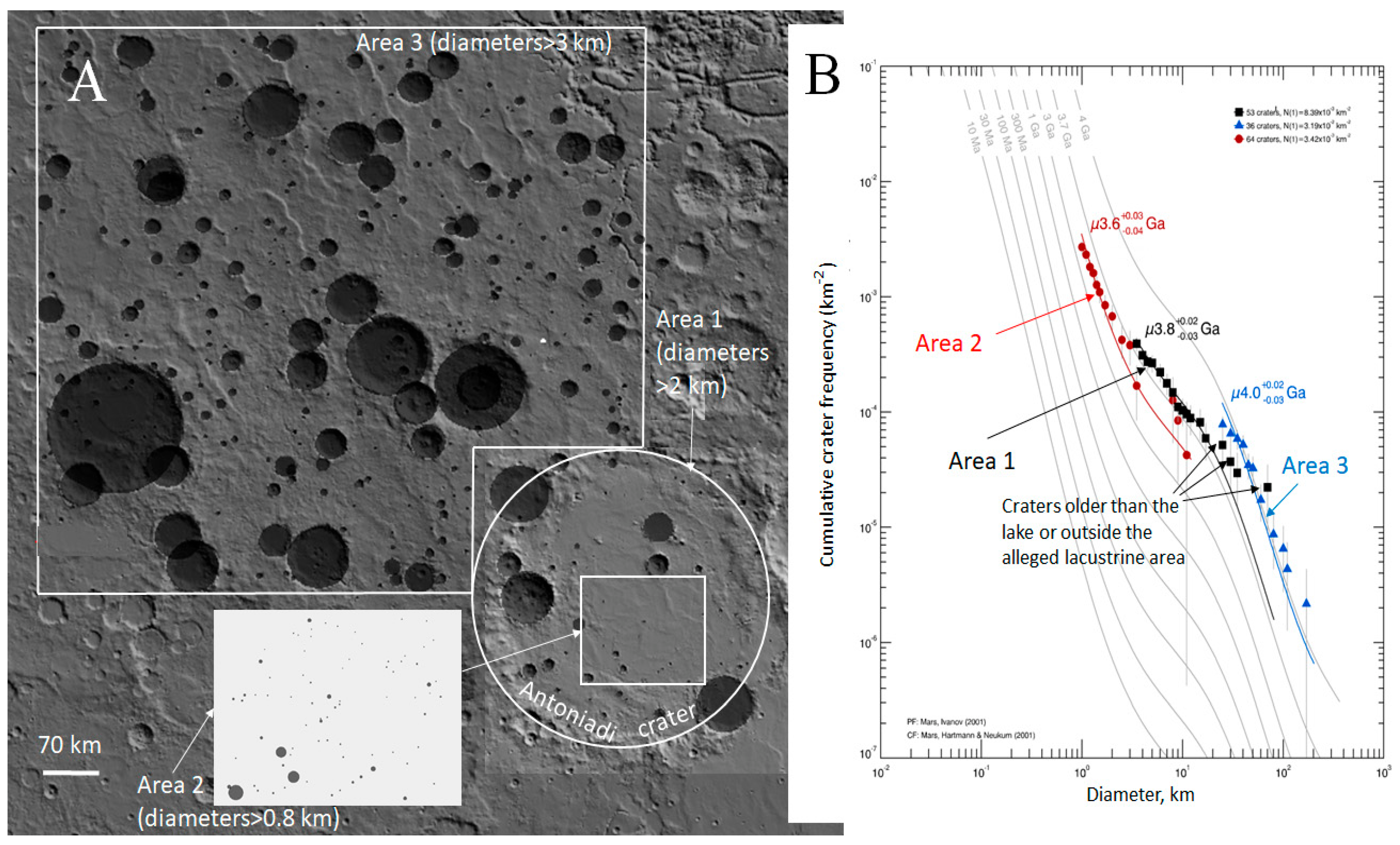
2.3. Age of the Antoniadi Crater Bottom
2.4. Standard Interpretation of the Dendrites and Its Possible Difficulties

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Example (Earth) | Example (Mars) | Superposition between Branches | Characteristic Slope (m/m) | Branches in Relief? | Inferred Direction of Gravity |
|---|---|---|---|---|---|---|
| Riverine surface precipitation–runoff in drainage basins | Most river drainages | Some examples on ancient terrains Warrego Valles (Figure 7C) | Absent | Typically, 1/50 on the large-catchment areas on Mars. On Earth, up to 1/1 in the small catchment areas at high altitude down to 1/50 in the largest and lowest ones | Possible if the water-carried sediment is compacted to a deposit less erodible than the floor (e.g., Figure 7A) | Branches to stem |
| Delta or alluvial fan | Many large and small rivers | Numerous examples (e.g., Figure 7B) | Common (braided) | 1/20 to 1/10 (up to ½ for the submerged part in Gilbert-type deltas); On Earth, 1/40 to 1/2 | Possible if carried sediment harder than floor (e.g., Figure 7B) | Stem to branches |
| Groundwater sapping | Grand Canyon | Numerous examples in Valles Marineris | Mostly absent | 1/50 to 1/8 (Valles Marineris); up to 1/10 on Earth (e.g., Grand Canyon) | Not known | Branches to stem |
| Biotic (Earth) | Stromatolite reefs | Unknown | Possible on dead branches | Virtually null (parallel to shore), to 1/20 (perpendicular to shore) | Occurs naturally | Growth possible in both ways |
| Antoniadi | Possible microbialite reefs? | Observed, but uncommon | Less than 1/200 | Yes | Growth possible in both ways |

3. Possible Alternate View for Dendrites within the Antoniadi Crater
3.1. Microbialite Interpretation
3.2. Further Morphological Evidence in Favor of the Microbialite Interpretation
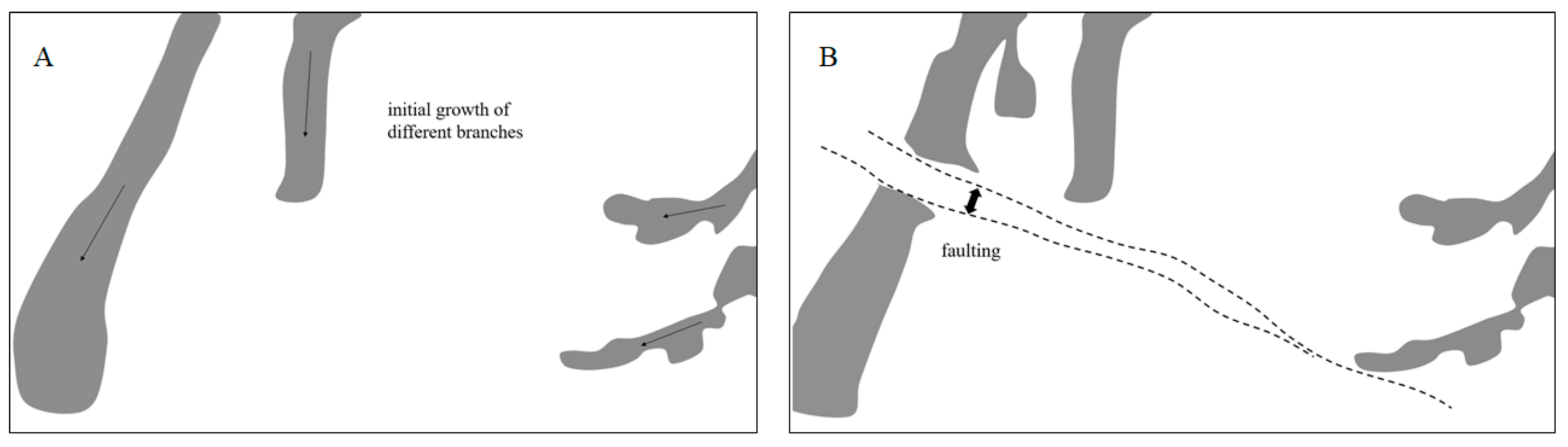
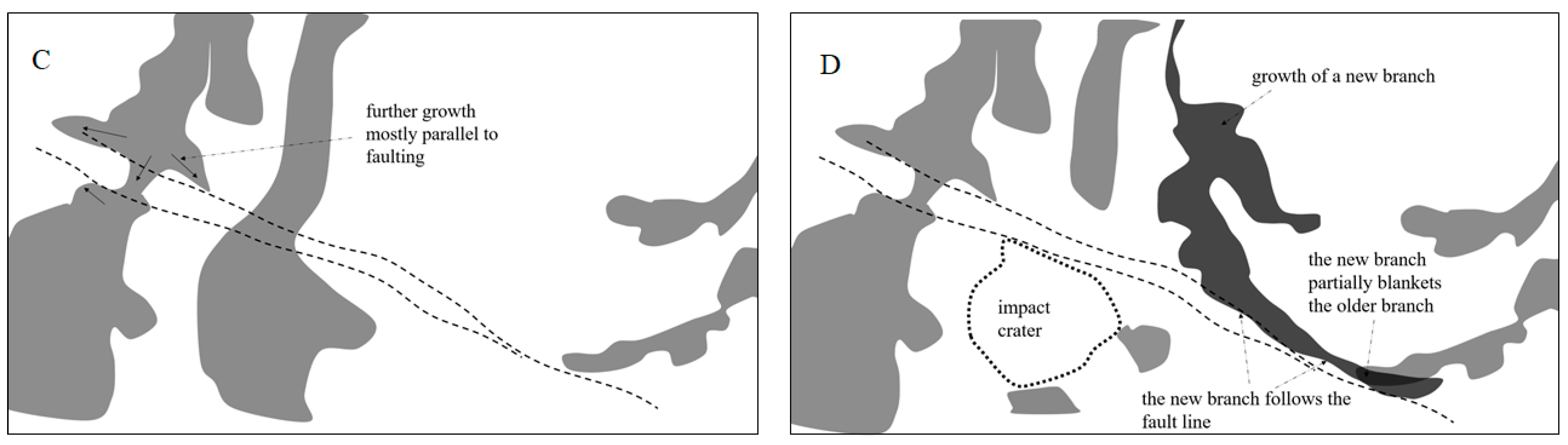
3.3. Dendrites on Kilometer-Size Scale and Small-Scale Faults
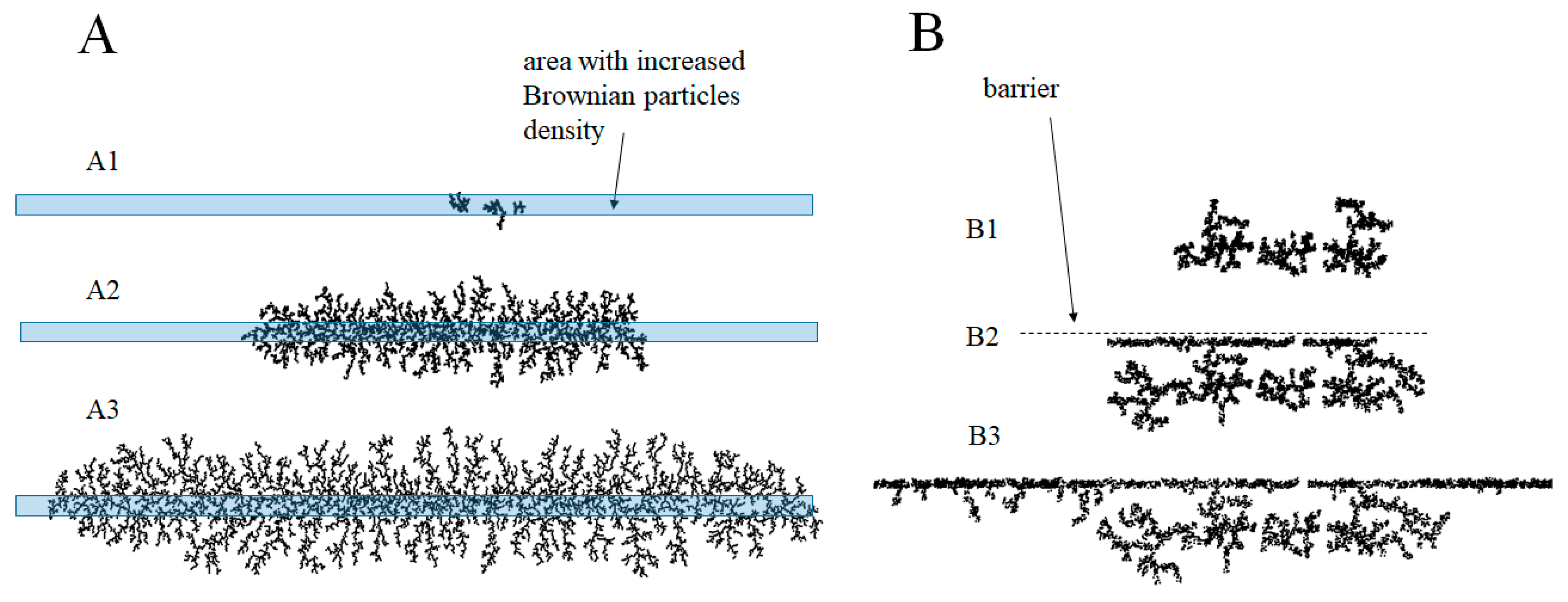
3.4. Similarity to Diffusion-Controlled Aggregation in Mineral Growth
3.5. Numerical Simulations
4. Discussion
4.1. The Privileged Condition in the Interior of the Antoniadi Crater
4.2. Microbialites and Stromatolites on Earth and Mars
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Transfer of Rocky Material between Earth and Mars
Appendix B. Data Sources
References
- Kereszturi, A.; Petrik, A. Age determination for valley networks on Mars using tectonic-fluvial interaction. Planet. Space Sci. 2020, 180, 104754. [Google Scholar] [CrossRef]
- Irwin, L.N.; Schulze-Makuch, D. The astrobiology of alien worlds: Known and unknown forms of life. Universe 2020, 6, 130. [Google Scholar] [CrossRef]
- Orosei, R.; Ding, C.; Fa, W.; Giannopoulos, A.; Hérique, A.; Kofman, W.; Lauro, S.E.; Li, C.; Pettinelli, E.; Su, Y.; et al. The Global Search for Liquid Water on Mars from Orbit: Current and Future Perspectives. Life 2020, 10, 120. [Google Scholar] [CrossRef]
- Tarnas, J.D.; Mustard, J.F.; Sherwood Lollar, B.; Stamenković, V.; Cannon, K.M.; Lorand, J.P.; Onstott, T.C.; Michalski, J.R.; Warr, O.; Palumbo, A.M.; et al. Earth-like Habitable Environments in the Subsurface of Mars. Astrobiology 2021, 21, 741–756. [Google Scholar] [CrossRef] [PubMed]
- Baker, V.R.; Strom, R.G.; Gulick, V.C.; Kargel, J.S.; Komatsu, G.; Kale, V.S. Ancient oceans, ice sheets and the hydrological cycle on Mars. Nature 1991, 352, 589–594. [Google Scholar] [CrossRef]
- Carr, M.H. Water on Mars; Oxford University Press: New York, NY, USA, 1996. [Google Scholar]
- Baker, V. Water and the Martian landscape. Nature 2001, 412, 228–236. [Google Scholar] [CrossRef] [Green Version]
- Carr, M.H.; Head, J.W., III. Oceans on Mars: An assessment of the observational evidence and possible fate. J. Geophys. Res. Planets 2003, 108. [Google Scholar] [CrossRef]
- Hynek, B.M.; Beach, M.; Hoke, M.R. Updated global map of Martian valley networks and implications for climate and hydrologic processes. J. Geophys. Res. Planets 2010, 115. [Google Scholar] [CrossRef]
- Luo, W.; Cang, X.; Howard, A.D. New Martian valley network volume estimate consistent with ancient ocean and warm and wet climate. Nat. Commun. 2017, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Parker, T.J.; Gorsline, D.S.; Saunders, R.S.; Pieri, D.C.; Schneeberger, D.M. Coastal geomorphology of the Martian northern plains. J. Geophys. Res. Planets 1993, 98, 11061–11078. [Google Scholar] [CrossRef]
- Parker, T.J.; Grant, J.A.; Franklin, B.J. The northern plain: A Martian oceanic basin? In Lakes on Mars; Cabrol, N.A., Grin, E.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2010. [Google Scholar]
- Bishop, J.L. Remote detection of phyllosilicates on Mars and implications for climate and habitability. In From Habitability to Life on Mars; Elsevier: Amsterdam, The Netherlands, 2018; pp. 37–75. [Google Scholar]
- Lucchitta, B.K.; McEwen, A.S.; Clow, G.D.; Geissler, P.E.; Singer, R.B.; Schultz, R.A.; Squyres, S.W. The Canyon System on Mars; Mars, Kieffer, H.H., Eds.; University of Arizona Press: Tucson, AZ, USA, 1992; pp. 453–492. [Google Scholar]
- Di Achille, G.; Hynek, B.M. Ancient ocean on Mars supported by global distribution of deltas and valleys. Nat. Geosci. 2010, 3, 459–463. [Google Scholar] [CrossRef]
- DiBiase, R.A.; Limaye, A.B.; Scheingross, J.S.; Fischer, W.W.; Lamb, M.P. Deltaic deposits at Aeolis Dorsa: Sedimentary evidence for a standing body of water on the northern plains of Mars. J. Geophys. Res. Planets 2013, 118, 1285–1302. [Google Scholar] [CrossRef] [Green Version]
- Goudge, T.A.; Milliken, R.E.; Head, J.W.; Mustard, J.F.; Fassett, C.I. Sedimentological evidence for a deltaic origin of the western fan deposit in Jezero crater, Mars and implications for future exploration. Earth Planet. Sci. Lett. 2017, 458, 357–365. [Google Scholar] [CrossRef]
- De Blasio, F.V. Possible erosion marks of bottom oceanic currents in the northern lowlands of Mars. Planet. Space Sci. 2014, 93, 10–21. [Google Scholar] [CrossRef]
- Moscardelli, L. Boulders of the Vastitas Borealis Formation: Potential origin and implications for an ancient martian ocean. GSA Today 2014, 24, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, J.A.P.; Fairén, A.G.; Tanaka, K.L.; Zarroca, M.; Linares, R.; Platz, T.; Komatsu, G.; Miyamoto, H.; Kargel, J.S.; Yan, J.; et al. Tsunami waves extensively resurfaced the shorelines of an early Martian ocean. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- De Blasio, F.V. Frontal Aureole Deposit on Acheron Fossae ridge as evidence for landslide-generated tsunami on Mars. Planet. Space Sci. 2020, 187, 104911. [Google Scholar] [CrossRef]
- Wharton, R.A.; Crosby, J.M.; McKay, C.P.; Rice, J.W. Paleolakes on Mars. J. Paleolimnol. 1995, 13, 267–283. [Google Scholar] [CrossRef]
- De Hon, R.A. Hydrologic provinces of Mars: Physiographic controls on drainage and ponding. In Lakes on Mars; Elsevier: Amsterdam, The Netherlands, 2010; pp. 68–89. [Google Scholar]
- Lucchitta, B.K. Lakes in Valles Marineris. In Lakes on Mars; Elsevier: Amsterdam, The Netherlands, 2010; pp. 111–161. [Google Scholar]
- Harrison, K.P.; Chapman, M.G. Evidence for ponding and catastrophic floods in central Valles Marineris, Mars. Icarus 2008, 198, 351–364. [Google Scholar] [CrossRef]
- Cabrol, N.A.; Grin, E.A. Searching for lakes on Mars: Four decades of exploration. In Lakes on Mars; Elsevier: Amsterdam, The Netherlands, 2010; pp. 1–29. [Google Scholar]
- Newsom, H.E.; Brittelle, G.E.; Hibbitts, C.A.; Crossey, L.J.; Kudo, A.M. Impact crater lakes on Mars. J. Geophys. Res. Planets 1996, 101, 14951–14955. [Google Scholar] [CrossRef]
- Arvidson, R.E.; Catalano, J.G. Martian Habitability as Inferred From Landed Mission Observations. In From Habitability to Life on Mars; Elsevier: Amsterdam, The Netherlands, 2018; pp. 77–126. [Google Scholar]
- Ori, G.G.; Marinangeli, L.; Baliva, A. Terraces and Gilbert-type deltas in crater lakes in Ismenius Lacus and Memnonia (Mars). J. Geophys. Res. Planets 2000, 105, 17629–17641. [Google Scholar] [CrossRef]
- Flannery, D.T.; Summons, R.E.; Walter, M.R. Archean Lakes as analogues for habitable Martian paleoenvironments. In From Habitability to Life on Mars; Elsevier: Amsterdam, The Netherlands, 2018; pp. 127–152. [Google Scholar]
- Smith, M.R.; Bandfield, J.L.; Gillespie, A.R. Felsic and altered mineral suite in Antoniadi Crater, Mars as a future rover landing site. In Proceedings of the 42nd Lunar and Planetary Science Conference, Woodlands, TX, USA, 7–11 March 2011; p. 1671. [Google Scholar]
- Smith, M.R.; Bandfield, J.L. Geology of quartz and hydrated silica-bearing deposits near Antoniadi Crater, Mars. J. Geophys. Res. Planets 2012, 117. [Google Scholar] [CrossRef]
- Garvin, J.B.; Sakimoto, S.E.H.; Frawley, J.J. Craters on Mars: Global geometric properties from gridded MOLA. In Proceedings of the Sixth International Conference on Mars, Pasadena, CA, USA, 20–25 July 2003; Volume 3. [Google Scholar]
- Zegers, T.E.; Oosthoek, J.H.; Rossi, A.P.; Blom, J.K.; Schumacher, S. Melt and collapse of buried water ice: An alternative hypothesis for the formation of chaotic terrains on Mars. Earth Planet. Sci. Lett. 2010, 297, 496–504. [Google Scholar] [CrossRef]
- McEwen, A. Branched Features on the Floor of Antoniadi Crater. 2009. Available online: https://www.uahirise.org/ESP_012435_2015 (accessed on 31 July 2020).
- Zaki, A.S.; Pain, C.F.; Edgett, K.S.; Giegengack, R. Inverted stream channels in the Western Desert of Egypt: Synergistic remote, field observations and laboratory analysis on Earth with applications to Mars. Icarus 2018, 309, 105–124. [Google Scholar] [CrossRef]
- Brooker, L.M.; Balme, M.R.; Conway, S.J.; Hagermann, A.; Barrett, A.M.; Collins, G.S.; Soare, R.J. Clastic polygonal networks around Lyot crater, Mars: Possible formation mechanisms from morphometric analysis. Icarus 2018, 302, 386–406. [Google Scholar] [CrossRef]
- Harris, R.C. Giant Desiccation Cracks in Arizona; Arizona Geological Survey: Tucson, AZ, USA, 2004; 93p. [Google Scholar]
- El-Maarry, M.R.; Watters, W.; McKeown, N.K.; Carter, J.; Dobrea, E.N.; Bishop, J.L.; Pommerol, A.; Thomas, N. Potential desiccation cracks on Mars: A synthesis from modeling, analogue-field studies, and global observations. Icarus 2014, 241, 248–268. [Google Scholar] [CrossRef]
- Michael, G.G. Planetary surface dating from crater size–frequency distribution measurements: Multiple resurfacing episodes and differential isochron fitting. Icarus 2013, 226, 885–890. [Google Scholar] [CrossRef]
- Christensen, P.R.; Jakosky, B.M.; Kieffer, H.H.; Malin, M.C.; McSween, H.Y.; Nealson, K.; Mehall, G.L.; Silverman, S.H.; Ferry, S.; Caplinger, M.; et al. The thermal emission imaging system (THEMIS) for the Mars 2001 Odyssey Mission. Space Sci. Rev. 2004, 110, 85–130. [Google Scholar] [CrossRef]
- Michael, G.G.; Neukum, G. Planetary surface dating from crater size–frequency distribution measurements: Partial resurfacing events and statistical age uncertainty. Earth Planet. Sci. Lett. 2010, 294, 223–229. [Google Scholar] [CrossRef]
- Tanaka, K.L.; Skinner, J.A., Jr.; Dohm, J.M.; Irwin, R.P., III; Kolb, E.J.; Fortezzo, C.M.; Platz, T.; Michael, G.G.; Hare, T.M. Geologic Map of Mars; USGS: Reston, VA, USA, 2014.
- Soare, R.J.; Burr, D.M.; Tseung, J.M.W.B. Possible pingos and a periglacial landscape in northwest Utopia Planitia. Icarus 2005, 174, 373–382. [Google Scholar] [CrossRef]
- Orgel, C.; Hauber, E.; van Gasselt, S.; Reiss, D.; Johnsson, A.; Ramsdale, J.D.; Smith, I.; Swirad, Z.M.; Séjourné, A.; Wilson, J.T.; et al. Grid mapping the northern plains of Mars: A new overview of recent water-and ice-related landforms in Acidalia Planitia. J. Geophys. Res. Planets 2019, 124, 454–482. [Google Scholar] [CrossRef] [Green Version]
- Séjourné, A.; Costard, F.; Swirad, Z.M.; Łosiak, A.; Bouley, S.; Smith, I.; Balme, M.R.; Orgel, C.; Ramsdale, J.D.; Hauber, E.; et al. Grid mapping the northern plains of Mars: Using morphotype and distribution of ice-related landforms to understand multiple ice-rich deposits in Utopia Planitia. J. Geophys. Res. Planets 2019, 124, 483–503. [Google Scholar] [CrossRef] [Green Version]
- Mellon, M.T.; Arvidson, R.E.; Marlow, J.J.; Phillips, R.J.; Asphaug, E. Periglacial landforms at the Phoenix landing site and the northern plains of Mars. J. Geophys. Res. Planets 2008, 113. [Google Scholar] [CrossRef] [Green Version]
- El Maarry, M.R.; Markiewicz, W.J.; Mellon, M.T.; Goetz, W.; Dohm, J.M.; Pack, A. Crater floor polygons: Desiccation patterns of ancient lakes on Mars? J. Geophys. Res. Planets 2010, 115. [Google Scholar] [CrossRef] [Green Version]
- Chan, M.A.; Nicoll, K.; Ormö, J.; Okubo, C.; Komatsu, G. Utah’s geologic and geomorphic analogs to Mars—An overview for planetary exploration. Geol. Soc. Am. Spec. Pap. 2011, 483, 349–375. [Google Scholar]
- Pain, C.F.; Clarke, J.D.A.; Thomas, M. Inversion of relief on Mars. Icarus 2007, 190, 478–491. [Google Scholar] [CrossRef]
- Mangold, N.; Quantin, C.; Ansan, V.; Delacourt, C.; Allemand, P. Evidence for precipitation on Mars from dendritic valleys in the Valles Marineris area. Science 2004, 305, 78–81. [Google Scholar] [CrossRef]
- Burr, D.M.; Williams, R.M.; Wendell, K.D.; Chojnacki, M.; Emery, J.P. Inverted fluvial features in the Aeolis/Zephyria Plana region, Mars: Formation mechanism and initial paleodischarge estimates. J. Geophys. Res. Planets 2010, 115. [Google Scholar] [CrossRef] [Green Version]
- Leopold, L.B.; Wolman, M.G.; Miller, J.P. Fluvial Processes in Geomorphology; Courier Dover Publications: Mineola, NY, USA, 2020. [Google Scholar]
- Isherwood, R.J.; Jozwiak, L.M.; Jansen, J.C.; Andrews-Hanna, J.C. The volcanic history of Olympus Mons from paleo-topography and flexural modeling. Earth Planet. Sci. Lett. 2013, 363, 88–96. [Google Scholar] [CrossRef]
- Lefort, A.; Burr, D.M.; Nimmo, F.; Jacobsen, R.E. Channel slope reversal near the Martian dichotomy boundary: Testing tectonic hypotheses. Geomorphology 2015, 240, 121–136. [Google Scholar] [CrossRef]
- Banerdt, W.B.; Golombek, M.P.; Tanaka, K.L. Stress and tectonics on Mars. Mars 1992, 249–297. [Google Scholar]
- Golombek, M.P.; Phillips, R.J. Mars tectonics. In Planetary Tectonics; Watters, T.R., Schultz, R.A., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 183–232. [Google Scholar]
- Kah, L.C.; Bartley, J.K.; Stagner, A.F. Reinterpreting a proterozoic enigma: Conophyton–Jacutophyton stromatolites of the Mesoproterozoic Atar group, Mauritania. Int. Assoc. Sediment. Spec. Pub. 2009, 41, 277–295. [Google Scholar]
- Noffke, N. Geobiology: Microbial Mats in Sandy Deposits from the Archean Era to Today; Springer Science and Business Media: Berlin, Germany, 2010. [Google Scholar]
- Reitner, J.; Quéric, N.-V.; Arp, G. Advances in Stromatolite Geobiology; Springer: Heidelberg, Germany, 2011; 559p. [Google Scholar]
- Riding, R. Microbialites, stromatolites, and thrombolites. In Encyclopedia of Geobiology, Encyclopedia of Earth Sciences Series; Reitner, J., Thiel, V., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2011; pp. 635–654. [Google Scholar]
- Bosak, T.; Knoll, A.H.; Petroff, A.P. The meaning of stromatolites. Annu. Rev. Earth Planet. Sci. 2013, 41, 21–44. [Google Scholar] [CrossRef] [Green Version]
- Russell, M.J.; Ingham, J.K.; Zedef, V.; Maktav, D.; Sunar, F.; Hall, A.J.; Fallick, A.E. Search for signs of ancient life on Mars: Expectations from hydromagnesite microbialites, Salda Lake, Turkey. J. Geol. Soc. 1999, 156, 869–888. [Google Scholar] [CrossRef] [Green Version]
- Noffke, N. Ancient sedimentary structures in the <3.7b Ga Gillespie Lake Member, Mars, that Compare in macroscopic morphology, spatial associations, and temporal succession with terrestrrial microbialites. Astrobiology 2015, 15, 169–192. [Google Scholar] [CrossRef]
- Cady, S.L.; Skok, J.R.; Gulick, V.G.; Berger, J.A.; Hinman, N.W. Siliceous hot spring deposits: Why they remain key astrobiological targets. In From Habitability to Life on Mars; Elsevier: Amsterdam, The Netherlands, 2018; pp. 179–210. [Google Scholar]
- Talbot, M.R.; Allen, P.A. Lakes. In Sedimentary Environments: Processes, Facies and Stratigraphy; Reading, H.G., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 1996; pp. 83–124. [Google Scholar]
- Cockell, C.S.; Lee, P. The biology of impact craters—A review. Biol. Rev. 2002, 77, 279–310. [Google Scholar] [CrossRef]
- Altermann, W. Accretion, trapping and binding of sediment in Archean stromatolites morphological expression of the antiquity of life. Space Sci. Rev. 2008, 135, 55–79. [Google Scholar] [CrossRef]
- Awramik, S.M.; Buchheim, H.P. Giant stromatolites of the Eocene Green River Formation (Colorado, USA). Geology 2015, 43, 691–694. [Google Scholar] [CrossRef]
- Andres, M.S.; Reid, R.P. Growth morphologies of modern marine stromatolites: A case study from Highborne Cay, Bahamas. Sediment. Geol. 2006, 185, 319–328. [Google Scholar] [CrossRef]
- Ricketts, B.D.; Donaldson, J.A. Stromatolite Reef Development on a Mud-Dominated Platform in the Middle Precambrian Belcher Group of Hudson Bay; AAPG: Tulsa, OK, USA, 1988. [Google Scholar]
- Bridge, J.; Demicco, R. Earth Surface Processes, Landforms and Sediment Deposits; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Peters, S.E.; Husson, J.M.; Wilcots, J. The rise and fall of stromatolites in shallow marine environments. Geology 2017, 45, 487–490. [Google Scholar] [CrossRef]
- Horgan, B.H.; Anderson, R.B.; Dromart, G.; Amador, E.S.; Rice, M.S. The mineral diversity of Jezero crater: Evidence for possible lacustrine carbonates on Mars. Icarus 2019, 339, 113526. [Google Scholar] [CrossRef]
- Grotzinger, J.P.; Knoll, A.H. Stromatolites in Precambrian carbonates: Evolutionary mileposts or environmental dipsticks? Annu. Rev. Earth Planet. Sci. 1999, 27, 313–358. [Google Scholar] [CrossRef] [PubMed]
- Grey, K.; Planavsky, N.J. Microbialites of Lake Thetis, Cervantes, Western Australia: A Field Guide; Geological Survey of Western Australia: Perth, Australia, 2009.
- Grey, K.; Moore, L.S.; Burne, R.V.; Pierson, B.K.; Bauld, J. Lake Thetis, Western Australia: And example of saline lake sedimentation dominated by benthic microbial processes. Aust. J. Mar. Freshw. Res. 1990, 41, 275–300. [Google Scholar] [CrossRef]
- Meltzner, A.J.; Woodroffe, C.D. Coral microatolls. In Handbook of Sea-Level Research; Wiley: Hoboken, NJ, USA, 2015; pp. 125–145. [Google Scholar]
- Getzin, S.; Yizhaq, H.; Bell, B.; Erickson, T.E.; Postle, A.C.; Katra, I.; Tsuk, O.; Zelnik, Y.R.; Wiegand, K.; Wiegand, T.; et al. Discovery of fairy circles in Australia supports self-organization theory. Proc. Natl. Acad. Sci. USA 2016, 113, 3551–3556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horodyski, R.J.; Bloeser, B.; Vonder Haar, S. Laminated algal mats from a coastal lagoon, Laguna Mormona, Baja California, Mexico. J. Sediment. Res. 1977, 47, 680–696. [Google Scholar]
- Kovalchuk, O.; Owttrim, G.W.; Konhauser, K.O.; Gingras, M.K. Desiccation cracks in siliciclastic deposits: Microbial mat-related compared to abiotic sedimentary origin. Sediment. Geol. 2017, 347, 67–78. [Google Scholar] [CrossRef]
- Seibold, E.; Berger, W. The Sea Floor: An Introduction to Marine Geology; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Barabási, A.L.; Stanley, H.E. Fractal Concepts in Surface Growth; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- García-Ruiz, J.M.; Otálora, F.; Sanchez-Navas, A.; Higes-Rolando, F.J. The formation of manganese dendrites as the mineral record of flow structures. In Fractals and Dynamic Systems in Geoscience; Springer: Berlin/Heidelberg, Germany, 1994; pp. 307–318. [Google Scholar]
- Dupraz, C.; Pattisina, R.; Verrecchia, E.P. Translation of energy into morphology: Simulation of stromatolite morphospace using a stochastic model. Sediment. Geol. 2006, 185, 185–203. [Google Scholar] [CrossRef] [Green Version]
- Witten, T.A., Jr.; Sander, L.M. Diffusion-limited aggregation, a kinetic critical phenomenon. Phys. Rev. Lett. 1981, 47, 1400. [Google Scholar] [CrossRef]
- Braga, F.L.; Ribeiro, M.S. Diffusion limited aggregation: Algorithm optimization revisited. Comput. Phys. Commun. 2011, 182, 1602–1605. [Google Scholar] [CrossRef]
- Parker, B.C.; Simmons, G.M., Jr.; Love, F.G.; Wharton, R.A., Jr.; Seaburg, K.G. Modern stromatolites in Antarctic dry valley lakes. BioScience 1981, 31, 656–661. [Google Scholar] [CrossRef]
- Billi, D. Desert cyanobacteria under space and planetary simulations: A tool for searching for life beyond Earth and supporting human space exploration. Int. J. Astrobiol. 2019, 18, 483–489. [Google Scholar] [CrossRef]
- Kargel, J.S. Mars—A Warmer, Wetter Planet; Springer Science and Business Media: Berlin, Germany, 2004. [Google Scholar]
- Newsom, H.E. Heated Lakes on Mars; Elsevier: Amsterdam, The Netherlands, 2010; pp. 93–110. [Google Scholar]
- Abramov, O.; Kring, D.A. Impact-induced hydrothermal activity on early Mars. J. Geophys. Res. Planets 2005, 110. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.P.; Paige, D.A.; Manning, C.E. Layering in the wall rock of Valles Marineris: Intrusive and extrusive magmatism. Geophys. Res. Lett. 2003, 30. [Google Scholar] [CrossRef]
- Yin, A. Structural analysis of the Valles Marineris fault zone: Possible evidence for large-scale strike-slip faulting on Mars. Lithosphere 2012, 4, 286–330. [Google Scholar] [CrossRef] [Green Version]
- Carter, J.; Poulet, F. Ancient plutonic processes on Mars inferred from the detection of possible anorthositic terrains. Nat. Geosci. 2013, 6, 1008–1012. [Google Scholar] [CrossRef]
- Werner, S.C. The global martian volcanic evolutionary history. Icarus 2009, 201, 44–68. [Google Scholar] [CrossRef]
- Skok, J.R.; Mustard, J.F.; Ehlmann, B.L.; Milliken, R.E.; Murchie, S.L. Silica deposits in the Nili Patera caldera on the Syrtis Major volcanic complex on Mars. Nat. Geosci. 2010, 3, 838–841. [Google Scholar] [CrossRef] [Green Version]
- Ruff, W.; Farmer, J.D. Silica deposits on Mars with features resembling hot spring biosignatures at El Tatio in Chile. Nat. Commun. 2016, 7, 13554. [Google Scholar] [CrossRef]
- Rizzo, V.; Cantasano, N. Possible organosedimentary structures on Mars. Int. J. Astrobiol. 2009, 8, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, V. Why should geological criteria used on Earth not be valid also for Mars? Evidence of possible microbialites and algae in extinct Martian lakes. Int. J. Astrobiol. 2020, 19, 283–294. [Google Scholar] [CrossRef]
- McKay, C.P.; Stoker, C.R. The early environment and its evolution on Mars: Implication for life. Rev. Geophys. 1989, 27, 189–214. [Google Scholar] [CrossRef] [Green Version]
- Tewari, V.C. Earliest Microbes on Earth and Possible Occurrence of Stromatolites on Mars. In Exobiology: Matter, Energy, and Information in the Origin and Evolution of Life in the Universe; Chela-Flores, J., Raulin, F., Eds.; Springer: Dordrecht, The Netherlands, 1998. [Google Scholar]
- Williams, S.H.; Zimbelman, J.R. “ White Rock”: An eroded Martian lacustrine deposit (?). Geology 1994, 22, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Horodyski, R.J. Lyngbya mats at Laguna Mormona, Baja California, Mexico; comparison with Proterozoic stromatolites. J. Sediment. Res. 1977, 47, 1305–1320. [Google Scholar]
- Nutman, A.P.; Bennett, V.C.; Friend, C.R.; Van Kranendonk, M.J.; Chivas, A.R. Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures. Nature 2016, 537, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Kamada, A.; Kuroda, T.; Kasaba, Y.; Terada, N.; Nakagawa, H. Global climate and river transport simulations of early Mars around the Noachian and Hesperian boundary. Icarus 2021, 368, 114618. [Google Scholar] [CrossRef]
- Guzewich, S.D.; Way, M.J.; Aleinov, I.; Wolf, E.T.; Del Genio, A.; Wordsworth, R.; Tsigaridis, K. 3D Simulations of the Early Martian Hydrological Cycle Mediated by a H2-CO2 Greenhouse. J. Geophys. Res. Planets 2021, 126, e2021JE006825. [Google Scholar] [CrossRef]
- Cockell, C.S.; Bush, T.; Bryce, C.; Direito, S.; Fox-Powell, M.; Harrison, J.P.; Lammer, H.; Landenmark, H.; Martin-Torres, J.; Nicholson, N.; et al. Habitability: A review. Astrobiology 2016, 16, 89–117. [Google Scholar] [CrossRef]
- Grotzinger, J.P.; Gupta, S.; Malin, M.C.; Rubin, D.M.; Schieber, J.; Siebach, K.; Calef, F. Deposition, exhumation, and paleoclimate of an ancient lake deposit, Gale crater, Mars. Science 2015, 350, aac7575. [Google Scholar] [CrossRef]
- Domagal-Goldman, S.D.; Wright, K.E.; Adamala, K.; De La Rubia, L.A.; Bond, J.; Dartnell, L.R.; Goldman, A.D.; Lynch, K.; Naud, M.E.; Paulino-Lima, I.G.; et al. The astrobiology primer v2.0. Astrobiology 2016, 16, 561. [Google Scholar]
- Kereszturi, A.; Bradák, B.; Chatzitheodoridis, E.; Ujvari, G. Indicators and methods to understand past environments from ExoMars rover drills. Orig. Life Evol. Biosph. 2016, 46, 435–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gladman, B. Destination: Earth. Martian meteorite delivery. Icarus 1997, 130, 228–246. [Google Scholar] [CrossRef]
- McSween, H.Y.; McSween, H.Y.; McSween, J. Meteorites and Their Parent Planets; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Mileikowsky, C.; Cucinotta, F.A.; Wilson, J.W.; Gladman, B.; Horneck, G.; Lindegren, L.; Melosh, J.; Rickman, H.; Zheng, J.Q. Natural transfer of viable microbes in space: 1. From Mars to Earth and Earth to Mars. Icarus 2000, 145, 391–427. [Google Scholar] [CrossRef] [PubMed]
- Malin, M.C.; Bell, J.F.; Cantor, B.A.; Caplinger, M.A.; Calvin, W.M.; Clancy, R.T.; Edgett, K.S.; Edwards, L.; Haberle, R.M.; James, P.B.; et al. Context camera investigation on board the Mars Reconnaissance Orbiter. J. Geophys. Res. Planets 2007, 112. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.E.; Zuber, M.T.; Solomon, S.C.; Phillips, R.J.; Head, J.W.; Garvin, J.B.; Banerdt, W.B.; Muhleman, D.O.; Pettengill, G.H.; Neumann, G.A.; et al. The global topography of Mars and implications for surface evolution. Science 1999, 284, 1495–1503. [Google Scholar] [CrossRef] [Green Version]
- McEwen, A.S.; Eliason, E.M.; Bergstrom, J.W.; Bridges, N.T.; Hansen, C.J.; Delamere, W.A.; Grant, J.A.; Gulick, V.C.; Herkenhoff, K.E.; Keszthelyi, L.; et al. Mars reconnaissance orbiter’s high resolution imaging science experiment (HiRISE). J. Geophys. Res. Planets 2007, 112. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, B.A. Mars/Moon cratering rate ratio estimates. Space Sci. Rev. 2001, 96, 87–104. [Google Scholar] [CrossRef]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Blasio, F.V. The Large Dendritic Morphologies in the Antoniadi Crater (Mars) and Their Potential Astrobiological Significance. Geosciences 2022, 12, 53. https://doi.org/10.3390/geosciences12020053
De Blasio FV. The Large Dendritic Morphologies in the Antoniadi Crater (Mars) and Their Potential Astrobiological Significance. Geosciences. 2022; 12(2):53. https://doi.org/10.3390/geosciences12020053
Chicago/Turabian StyleDe Blasio, Fabio Vittorio. 2022. "The Large Dendritic Morphologies in the Antoniadi Crater (Mars) and Their Potential Astrobiological Significance" Geosciences 12, no. 2: 53. https://doi.org/10.3390/geosciences12020053
APA StyleDe Blasio, F. V. (2022). The Large Dendritic Morphologies in the Antoniadi Crater (Mars) and Their Potential Astrobiological Significance. Geosciences, 12(2), 53. https://doi.org/10.3390/geosciences12020053