
A New Calcareous Nannofossil Record from the Lower Jurassic of Kermanshah, Western Iran: Implications for Biostratigraphy and Evolutionary Reconstructions


Abstract
:1. Introduction
2. Geological Context of the Studied Succession
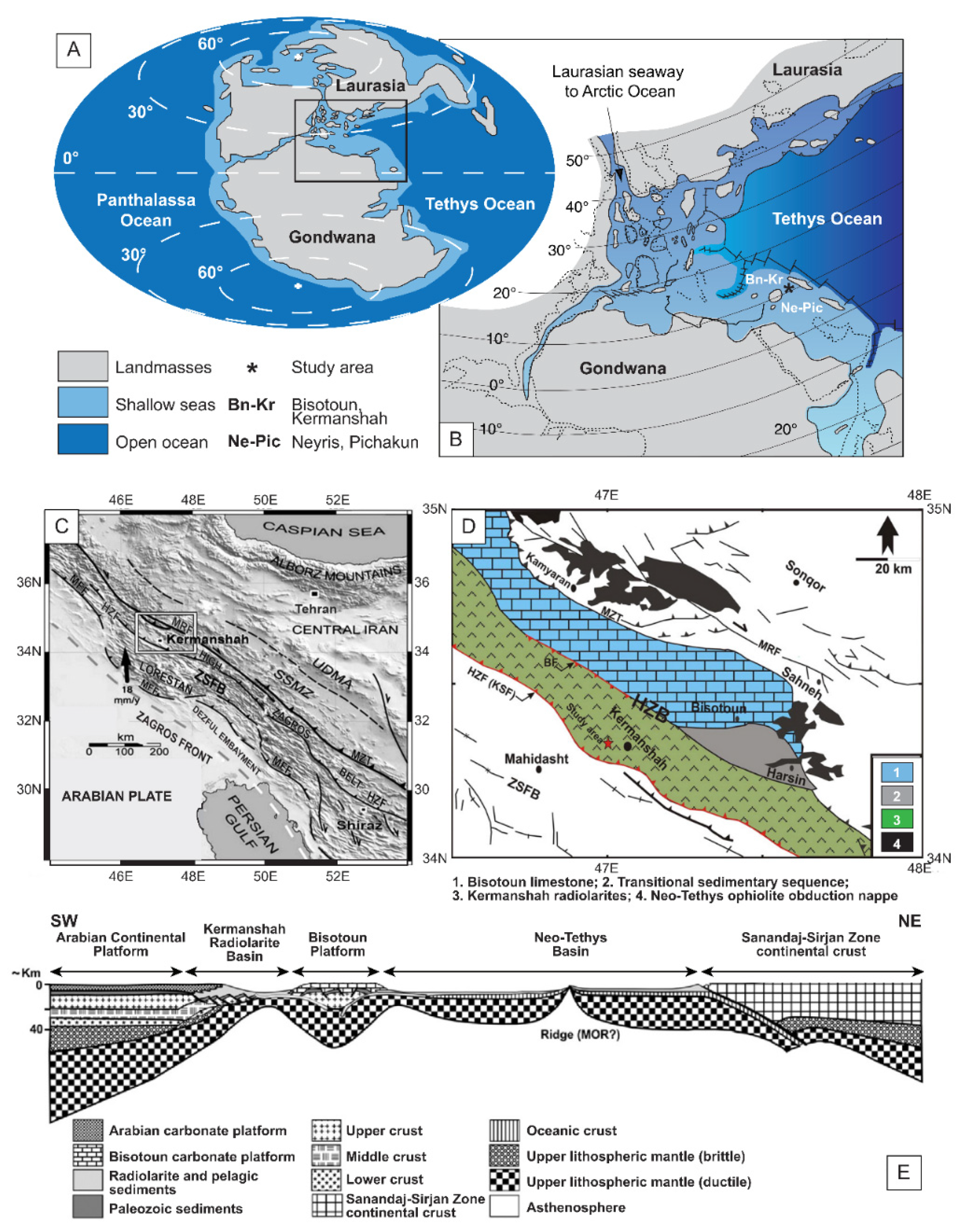
3. Nannofossil Sampling and Analysis
4. Results
4.1. Calcareous Nannofossil Biostratigraphy
4.2. Taxonomy of New Mitrolithus Species
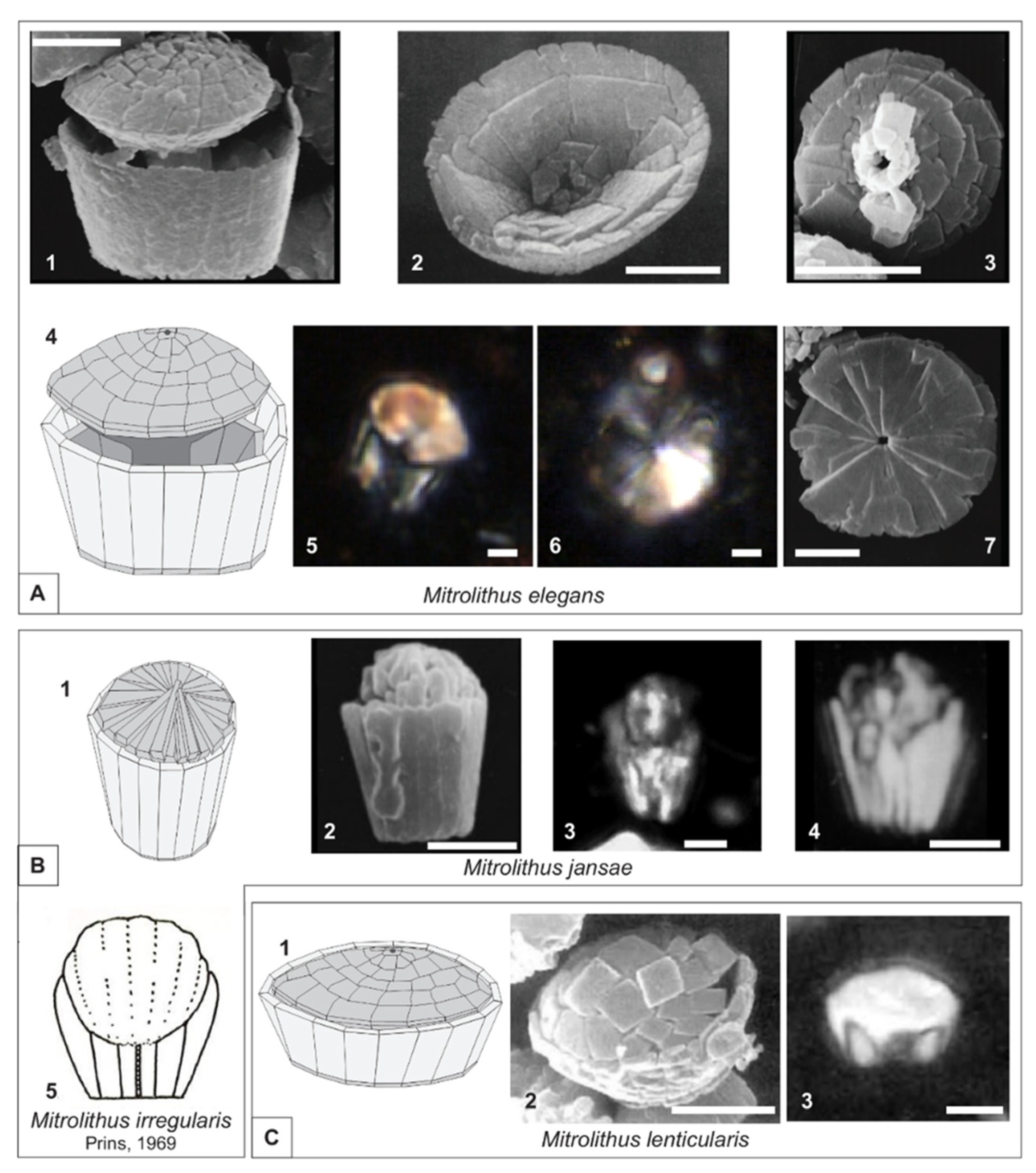
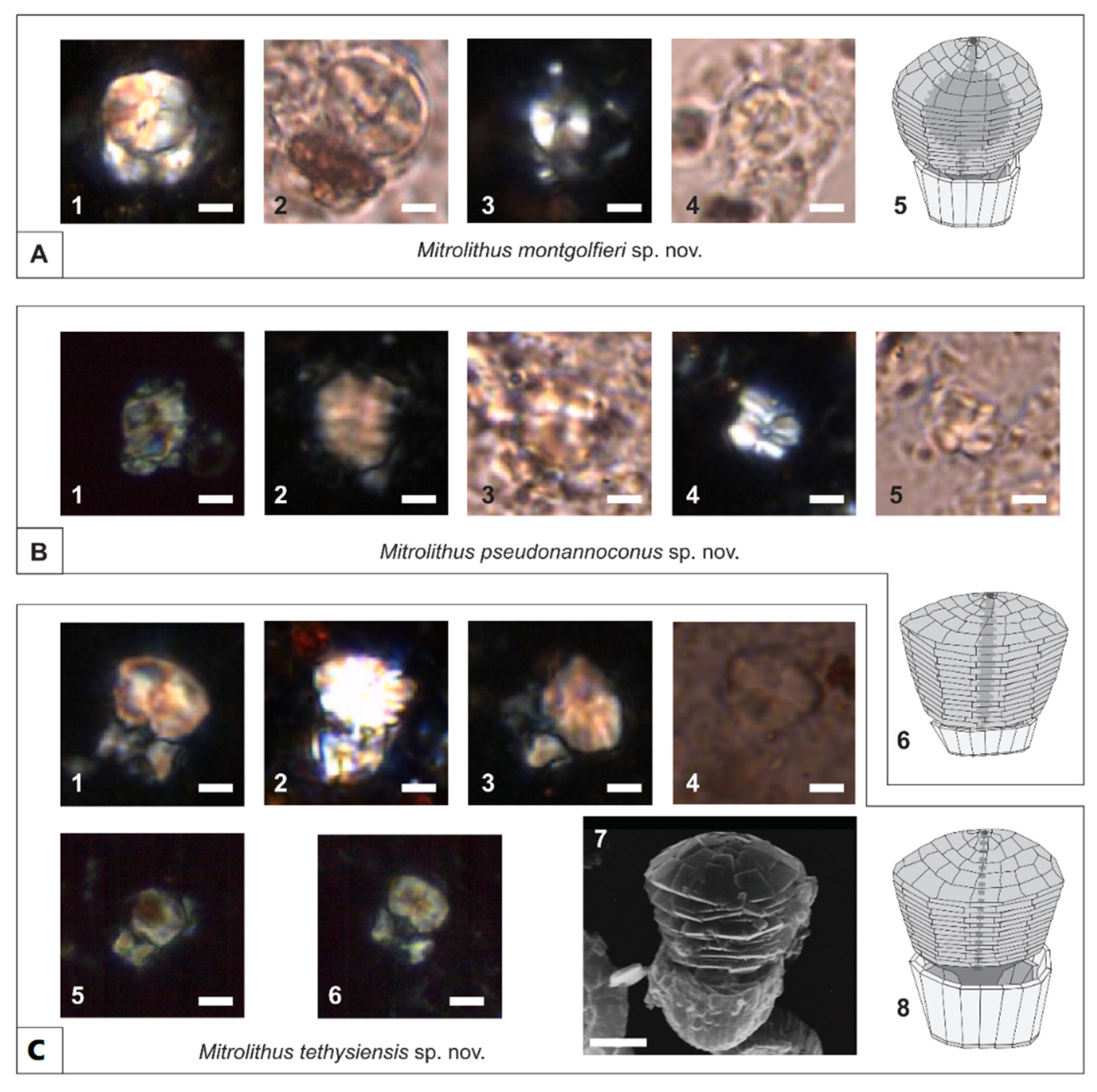
5. Discussion
5.1. Calcareous Nannofossil Biostratigraphy
5.2. Evolutionary and Paleogeographic Implications of the New Mitrolithus Species
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Alveariumdorsetense Black, 1965 |
| Crepidolithuscrassus (Deflandre in Deflandre and Fert, 1954) Noël, 1965 |
| Small-sized Crepidolithus crassus described in Suchéras-Marx et al. (2010) |
| Crepidolithuscavus Prins, 1969 |
| Crepidolithuscrucifer Rood et al. 1973 |
| Crepidolithusgranulatus Bown, 1987a |
| Crepidolithusimpontus Grün et al., 1974 |
| Crepidolithustimorensis Kristan-Tollmann, 1988 |
| Crucirhabdus aff. C. minutus Jafar, 1983 |
| Crucirhabdusprimulus Prins 1969 ex Rood et al., 1973, emended Bown, 1987a |
| Mazaganellaprotensa Bown, 1987a |
| Mazaganellapulla Bown, 1987a |
| Mitrolithuselegans Deflandre in Deflandre and Fert, 1954 |
| Mitrolithusjansae (Wiegand, 1984) Bown in Young et al., 1986 |
| Mitrolithuslenticularis Bown, 1987a |
| Mitrolithusmontgolfieri sp. nov. |
| Mitrolithuspseudonannoconus sp. nov. |
| Mitrolithustethysiensis sp. nov. |
| Parhabdolithusliasicus subsp. distinctus Bown, 1987a |
| Parhabdolithusliasicus subsp. liasicus Bown, 1987a |
| Parhabdolithusmarthae Deflandre in Deflandre and Fert, 1954 |
| Parhabdolithusrobustus Noël, 1965 |
| Schizosphaerella spp. Deflandre and Dangeard, 1938 |
| Similiscutumavitum de Kaenel and Bergen, 1993 |
| Similiscutumcruciulus de Kaenel and Bergen, 1993 |
| Similiscutumnovum (Goy in Goy et al., 1979) Mattioli et al., 2004 |
| Similiscutumorbiculus de Kaenel and Bergen, 1993 |
| Similiscutumprecarium de Kaenel and Bergen, 1993 |
| Timorellacypella Bown, 1987a |
| Tubirhabduspatulus Rood, Hay and Barnard, 1973 ex Prins, 1969 |
References
- Abdi, A.; Gharaie, M.H.M.; Bádenas, B. Internal wave deposits in Jurassic Kermanshah pelagic carbonates and radiolarites (Kermanshah area, West Iran). Sediment. Geol. 2014, 314, 47–59. [Google Scholar] [CrossRef]
- Carter, E.S.; Goričan, Š.; Guex, J.; O’Dogherty, L.; De Wever, P.; Dumitrica, P.; Hori, R.S.; Matsuoka, A.; Whalen, P.A. Global radiolarian zonation for the Pliensbachian, Toarcian and Aalenian. Palaeogeogr. Palaeoclim. Palaeoecol. 2010, 297, 401–419. [Google Scholar] [CrossRef]
- O’Dogherty, L.; Carter, E.S.; Dumitrica, P.; Goriččan, Š.; Wever, P.D.; Bandini, A.N.; Baumgartner, P.O.; Matsuoka, A. Catalogue of Mesozoic Radiolarian Genera. Part 2: Jurassic-Cretaceous. Geodiversitas 2009, 31, 271–356. [Google Scholar] [CrossRef]
- Barnard, T.; Hay, W.W. On Jurassic coccoliths: A tentative zonation of the Jurassic of Southern England and North France. Eclogae Geol. Helv. 1974, 67, 563–585. [Google Scholar]
- Hamilton, G. Lower and Middle Jurassic calcareous nannofossils from Portugal. Eclogae Geol. Helv. 1979, 72, 1–17. [Google Scholar]
- Hamilton, G.B. Triassic and Jurassic Calcareous Nannofossils. In A Stratigraphical Index of Calcareous Nannofossils; Ellis Horwood: Chichester, UK, 1982; pp. 17–39. [Google Scholar]
- Medd, A.W.; Medd, A.W. Nannofossil Zonation of the English Middle and Upper Jurassic Jurassic. Mar. Micropaleontol. 1982, 7, 73–95. [Google Scholar] [CrossRef]
- Bown, P.R. Taxonomy, evolution and biostratigraphy of Late Triassic-Early Jurassic calcareous nannofossils. Spec. Pap. Palaeontol. 1987, 38, 118. [Google Scholar]
- Bown, P.; Cooper, M.K.E.; Lord, A.R. A Calcareous Nannofossil Biozonation Scheme for the early to mid Mesozoic. Newsl. Strat. 1988, 20, 91–114. [Google Scholar] [CrossRef]
- Mattioli, E.; Erba, E. Biostratigraphic Synthesis of Calcareous Nannofossil Events in Tethyan Jurassic. Riv. Ital. Paléontol. Strat. 1999, 105, 343–376. [Google Scholar] [CrossRef]
- Ferreira, J.; Mattioli, E.; Suchéras-Marx, B.; Giraud, F.; Duarte, L.; Pittet, B.; Suan, G.; Hassler, A.; Spangenberg, J.E. Western Tethys Early and Middle Jurassic calcareous nannofossil biostratigraphy. Earth-Sci. Rev. 2019, 197, 102908. [Google Scholar] [CrossRef]
- Cornée, J.-J.; Martini, R.; Villeneuve, R.; Zaninetti, L.; Mattioli, E.; Rettori, R.; Atrops, F.; Gunawan, W. Jurassic Pelagic Deposits of East Sulawesi (Kolonodale Area, Indonesia): New Biostratigraphic Data Based on Calcareous Nannofossils. Geoitalia 1999, 32, 385–394. [Google Scholar]
- Shafik, S.; Organisation, A.G.S. Significance of Calcareous Nannofossil-Bearing Jurassic and Cretaceous Sediments on the Rowley Terrace, Offshore Northwest Australia. Available online: https://researchdata.edu.au/significance-calcareous-nannofossil-northwest-australia/688200 (accessed on 9 December 2021).
- Kazmin, V.; Ricou, L.-E.; Sbortshikov, I.M. Structure and Evolution of the Passive Margin of the Eastern Tethys. Tectonophysics 1986, 123, 153–179. [Google Scholar] [CrossRef]
- Bassoulet, J.P.; Elmi, S.; Poisson, A.; Cecca, F.; Bellion, Y.; Guiraud, R.; Baudin, F. Mid Toarcian. In Atlas Tethys Palaeoenvironmental Maps; Dercourt, J., Ricou, L.E., Vrielynck, B., Eds.; BEICIP-FRANLAB: Rueil-Malmaison, France, 1993; pp. 63–84. [Google Scholar]
- Robertson, A.H.F. Overview of tectonic settings related to the rifting and opening of Mesozoic ocean basins in the Eastern Tethys: Oman, Himalayas and Eastern Mediterranean regions. In Imaging, Mapping and Modelling Continental Lithosphere Extension and Breakup; Karner, G., Manatschal, G., Pinheiro, L.M., Eds.; Geological Society London Special Publications: Bath, UK, 2007; Volume 282, pp. 325–388. [Google Scholar] [CrossRef]
- Gharib, F.; De Wever, P. Radiolaires mésozoïques de la formation de Kermanshah (Iran). C. R. Palevol. 2010, 9, 209–219. [Google Scholar] [CrossRef]
- Navabpour, P.; Angelier, J.; Barrier, E. Brittle tectonic reconstruction of palaeo-extension inherited from Mesozoic rifting in West Zagros (Kermanshah, Iran). J. Geol. Soc. 2011, 168, 979–994. [Google Scholar] [CrossRef]
- Berra, F.; Angiolini, L.; Muttoni, G. The Evolution of the Tethys Region throughout the Phanerozoic: A Brief Tectonic Reconstruction. In Memoir 106: Petroleum Systems of the Tethyan Region; Marlow, L., Kendall, C.C.G., Rose, L.A., Eds.; AAPG: Tulsa, OK, USA, 2014; pp. 1–27. [Google Scholar]
- Mohajjel, M.; Fergusson, C.L.; Sahandi, M.R. Cretaceous–Tertiary Convergence and Continental Collision, Sanandaj–Sirjan Zone, Western Iran. J. Asian Earth Sci. 2003, 21, 397–412. [Google Scholar] [CrossRef]
- Abdi, A.; Bádenas, B.; Gharaie, M.H.M.; Gorican, S.; Toodekesht, S.; Mattioli, E. The Story of Kermanshah Radiolarite Basin from Early Jurassic to Late Cretaceous. In Proceedings of the 3rd Trigger International Conference, Zanjan, Iran, 5–6 October 2019; Institute for Advanced Studies in Basic Sciences: Zanjan, Iran, 2019. [Google Scholar]
- Wrobel-Daveau, J.-C.; Ringenbach, J.-C.; Tavakoli, S.; Ruiz, G.M.H.; Masse, P.; Frizon de Lamotte, D. Evidence for Mantle Exhumation along the Arabian Margin in the Zagros (Kermanshah Area, Iran). Arab. J. Geosci. 2010, 3, 499–513. [Google Scholar] [CrossRef]
- Abdi, A. Sedimentology and Geochemistry of Kermanshah Radiolarite Complex with Special Reference to Depositional Environment; Doctoral-Ferdowsi University of Mashhad: Mashhad, Iran, 2016. [Google Scholar]
- Abdi, A.; Gharaie, M.H.M.; Bádenas, B. Radiolarian productivity linked to climate conditions during the Pliensbachian–Aalenian in the Kermanshah Basin (West Iran). Facies 2016, 62, 30. [Google Scholar] [CrossRef]
- Alavi, M.; Mahdavi, M.A. Stratigraphy and structures of the Nahavand region in western Iran, and their implications for the Zagros tectonics. Geol. Mag. 1994, 131, 43–47. [Google Scholar] [CrossRef]
- Agard, P.; Omrani, J.; Jolivet, L.; Mouthereau, F. Convergence history across Zagros (Iran): Constraints from collisional and earlier deformation. Geol. Rundsch. 2005, 94, 401–419. [Google Scholar] [CrossRef]
- Braud, J. La Suture du Zagros au Niveau de Kermanshah (Kurdistan Iranien): Reconstitution Paléogéographique: Évolution Géodynamique, Magmatique et Structurale; Université de Paris-Sud: Paris, France, 1987; p. 11. [Google Scholar]
- Blakey, R.C. Gondwana Paleogeography from Assembly to Breakup—A 500 m.y. Odyssey. In Special Paper 441: Resolving the Late Paleozoic Ice Age in Time and Space; Geological Society of America: Boulder, CO, USA, 2008; Volume 441, pp. 1–28. [Google Scholar]
- Mattioli, E.; Pittet, B.; Young, J.R.; Bown, P.R. Biometric Analysis of Pliensbachian-Toarcian (Lower Jurassic) Coccoliths of the Family Biscutaceae: Intra- and Interspecific Variability versus Palaeoenvironmental Influence. Mar. Micropaleontol. 2004, 52, 5–27. [Google Scholar] [CrossRef]
- Bown, P.R.; Young, J.R. Techniques. In Calcareous Nannofossil Biostratigraphy; British Micropalaeontological Society Publications Series; Bown, P.R., Ed.; Chapman and Kluwer Academic: London, UK, 1998; pp. 16–28. [Google Scholar]
- Roth, P.H. Preservation of Calcareous Nannofossils and Fine-Grained Carbonate Particles in Mid-Cretaceous Sediments from the Southern Angola Basin. In Initial Reports of Deep Sea Drilling Project 75; Hay, W.W., Ed.; U.S. Government Printing Office: Washington, DC, USA, 1984; pp. 651–655. [Google Scholar]
- De Kaenel, E.; Bergen, J.A. New Early and Middle Jurassic Coccolith Taxa and Biostratigraphy from the Eastern Proto-Atlantic (Morocco, Portugal and DSDP Site 547 B). Eclogae Geol. Helv. 1993, 86, 861–907. [Google Scholar]
- Bown, P.R.; Cooper, M.K.E. Jurassic. In Calcareous Nannofossil Biostratigraphy; Bown, P.R., Ed.; British Micropalaeontological Society Publications Series: London, UK, 1998; pp. 34–85. [Google Scholar]
- Black, M. Coccoliths. Endeavour 1965, 24, 131–137. [Google Scholar]
- Deflandre, G.; Fert, C. Observations sur les Coccolithophoridés Actuels et Fossiles en Microscopie Ordinaire et Électronique. Ann. Paléontologie 1954, 40, 115–176. [Google Scholar]
- Young, J.R.; Teale, C.T.; Bown, P.R. Revision of the Stratigraphy of the Longobucco Group (Liassic, Southern Italy); Based on the New Data on Nannofossils and Ammonite. Eclogae Geol. Helv. 1986, 79, 117–135. [Google Scholar]
- Prins, B. Evolution and Stratigraphy of Coccolithinids from the Lower and Middle Lias. In Proceedings of the First International Conference on Planktonic Microfossils, Genève, Switzerland, 27 September–3 October 1969; pp. 547–558. [Google Scholar]
- Wiegand, G. Two Genera of Calcareous Nannofossils from the Lower Jurassic. J. Palaeontol. 1954, 58, 1151–1155. [Google Scholar]
- Casellato, C.E.; Erba, E. Calcareous Nannofossil Biostratigraphy and Paleoceanography of the Toarcian Oceanic Anoxic Event at Colle di Sogno Section (Southern Alps, Italy). Riv. Ital. Paleontol. E Stratigr. 2015, 121, 297–327. [Google Scholar] [CrossRef]
- Bown, P.R. The Structural Development of Early Mesozoic Coccoliths and Its Evolutionary and Taxonomic Significance. Abh. Geol. Bundesanst. 1987, 39, 33–49. [Google Scholar]
- Bown, P.R.; Cooper, M.K.E. Conical Calcareous Nannofossils in the Mesozoic. In Nannofossils and Their Applications; Crux, J.A., van Heck, S.E., Eds.; British Micropalaeontological Society Series; Ellis Horwood Ltd.: Chichester, UK, 1989; pp. 98–106. [Google Scholar]
- Menini, A.; Mattioli, E.; Spangenberg, J.E.; Pittet, B.; Suan, G. New Calcareous Nannofossil and Carbon Isotope Data for the Pliensbachian/Toarcian Boundary (Early Jurassic) in the Western Tethys and Their Paleoenvironmental Implications. Newsl. Stratigr. 2019, 52, 173–196. [Google Scholar] [CrossRef]
- Mattioli, E.; Pittet, B.; Suan, G.; Mailliot, S. Calcareous Nannoplankton Changes across the Early Toarcian Oceanic Anoxic Event in the Western Tethys: Implications for Paleoceanography within the Western Tethys. Paleoceanography 2008, 23, 3208. [Google Scholar] [CrossRef]
- Bucefalo Palliani, R.; Mattioli, E. Ecology of dinoflagellate cysts and calcareous nannofossils from bituminous facies of the Early Toarcian, central Italy. In Proceedings of the III EPA Workshop: Black Shales Models, Dotternhausen, Germany, 6–8 July 1995; pp. 60–62. [Google Scholar]
- Mattioli, E.; Pittet, B. Spatial and temporal distribution of calcareous nannofossils along a proximal–distal transect in the Lower Jurassic of the Umbria–Marche Basin (central Italy). Palaeogeogr. Palaeoclim. Palaeoecol. 2004, 205, 295–316. [Google Scholar] [CrossRef]
- Erba, E. Calcareous nannofossils and Mesozoic oceanic anoxic events. Mar. Micropaleontol. 2004, 52, 85–106. [Google Scholar] [CrossRef]
- Okada, H.; Honjo, S. The distribution of oceanic coccolithophorids in the Pacific. Deep Sea Res. Oceanogr. Abstr. 1973, 20, 355–374. [Google Scholar] [CrossRef]
- Bour, I.; Mattioli, E.; Pittet, B. Nannofacies analysis as a tool to reconstruct paleoenvironmental changes during the Early Toarcian anoxic event. Palaeogeogr. Palaeoclim. Palaeoecol. 2007, 249, 58–79. [Google Scholar] [CrossRef]
- Fraguas, Á.; Erba, E. Biometric analyses as a tool for the differentiation of two coccolith species of the genus Crepidolithus (Pliensbachian, Lower Jurassic) in the Basque-Cantabrian Basin (Northern Spain). Mar. Micropaleontol. 2010, 77, 125–136. [Google Scholar] [CrossRef]
- Reggiani, L.; Mattioli, E.; Pittet, B.; Duarte, L.; de Oliveira, L.V.; Comas-Rengifo, M. Pliensbachian (Early Jurassic) calcareous nannofossils from the Peniche section (Lusitanian Basin, Portugal): A clue for palaeoenvironmental reconstructions. Mar. Micropaleontol. 2010, 75, 1–16. [Google Scholar] [CrossRef]
- Suchéras-Marx, B.; Mattioli, E.; Pittet, B.; Escarguel, G.; Suan, G. Astronomically-paced coccolith size variations during the early Pliensbachian (Early Jurassic). Palaeogeogr. Palaeoclim. Palaeoecol. 2010, 295, 281–292. [Google Scholar] [CrossRef] [Green Version]
- Clémence, M.-E.; Gardin, S.; Bartolini, A. New insights in the pattern and timing of the Early Jurassic calcareous nannofossil crisis. Palaeogeogr. Palaeoclim. Palaeoecol. 2015, 427, 100–108. [Google Scholar] [CrossRef]
- Ferreira, J.; Mattioli, E.; Pittet, B.; Cachão, M.; Spangenberg, J.E. Palaeoecological insights on Toarcian and lower Aalenian calcareous nannofossils from the Lusitanian Basin (Portugal). Palaeogeogr. Palaeoclim. Palaeoecol. 2015, 436, 245–262. [Google Scholar] [CrossRef]
- Erba, E. Nannofossils and superplumes: The Early Aptian “nannoconid crisis”. Paleoceanography 1994, 9, 483–501. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kani Sad Section (Abdi et al. [1,23,24]) | Present Paper | ||||||
|---|---|---|---|---|---|---|---|
| Lithological member | Thickness and dominant lithology | Age | Kani Sad and Sad sections and nannofossil zones (present tectonic position) | Restored position and age | |||
| J.3 | 22 m, ribbon cherts | Middle Toarcian-Aalenian | Not studied | ||||
 | |||||||
| J.2 | 18 m, limestones with marl intercalations | Pliensbachian p.p.-early Toarcian | J.1 20 m, limestones with marl intercalations | J.0 | NJT 4a | Lower Pliensba-chian | |
| NJT 3b-c | NJT 3b-c | Upper Sinemurian | |||||
 | uncertain interval | ||||||
| J.1 | 3 m, cherts | J.0 up to 12 m, cherts, shales and calciturbidites | NJT 4a | ||||
| J.1 | NJT 3b | ||||||
| NJT 3c | |||||||
 ; Probable fault contact
; Probable fault contact  .
.Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdi, A.; Mattioli, E.; Bádenas, B. A New Calcareous Nannofossil Record from the Lower Jurassic of Kermanshah, Western Iran: Implications for Biostratigraphy and Evolutionary Reconstructions. Geosciences 2022, 12, 59. https://doi.org/10.3390/geosciences12020059
Abdi A, Mattioli E, Bádenas B. A New Calcareous Nannofossil Record from the Lower Jurassic of Kermanshah, Western Iran: Implications for Biostratigraphy and Evolutionary Reconstructions. Geosciences. 2022; 12(2):59. https://doi.org/10.3390/geosciences12020059
Chicago/Turabian StyleAbdi, Asad, Emanuela Mattioli, and Beatriz Bádenas. 2022. "A New Calcareous Nannofossil Record from the Lower Jurassic of Kermanshah, Western Iran: Implications for Biostratigraphy and Evolutionary Reconstructions" Geosciences 12, no. 2: 59. https://doi.org/10.3390/geosciences12020059
APA StyleAbdi, A., Mattioli, E., & Bádenas, B. (2022). A New Calcareous Nannofossil Record from the Lower Jurassic of Kermanshah, Western Iran: Implications for Biostratigraphy and Evolutionary Reconstructions. Geosciences, 12(2), 59. https://doi.org/10.3390/geosciences12020059