

Calcareous Nannofossils Biostratigraphy of Late Cretaceous–Paleocene Successions from Northern Jordan and Their Implications for Basin Analysis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Study Area
2.1. Location Area and Geological Setting
2.2. Biostratigraphy
3. Materials and Methods
3.1. Sampling
3.2. Nannofossils Examinations
3.2.1. Taxonomy
3.2.2. Preservation/Dissolution Indices
3.2.3. Statistical Analysis
3.2.4. Calcareous Nannofossil Nutrient and Temperature Indices
3.2.5. Sedimentation Rate
4. Results
4.1. Calcareous Nannofossil Assemblages
4.2. Biostratigraphy
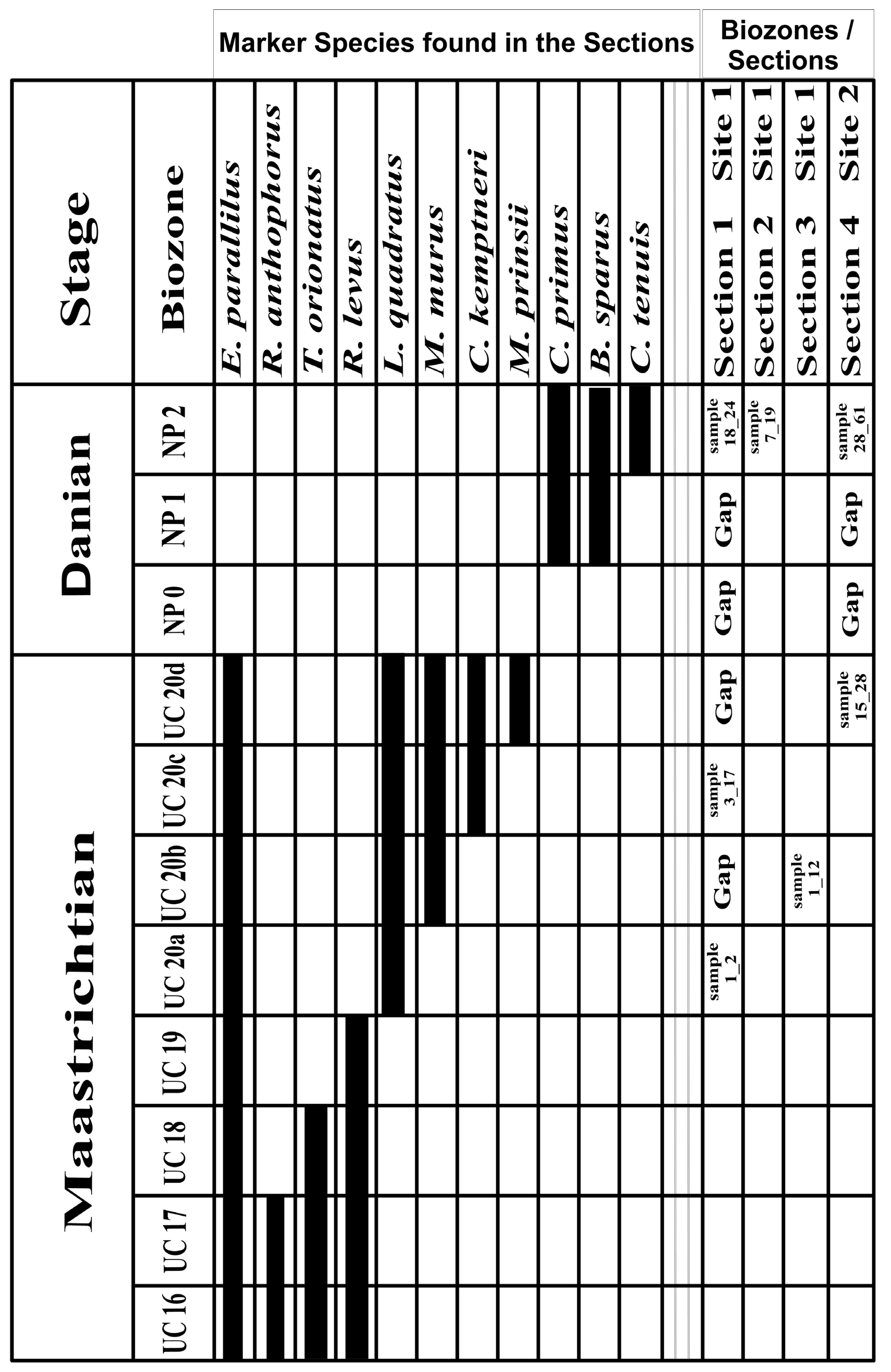
4.2.1. Biozonation
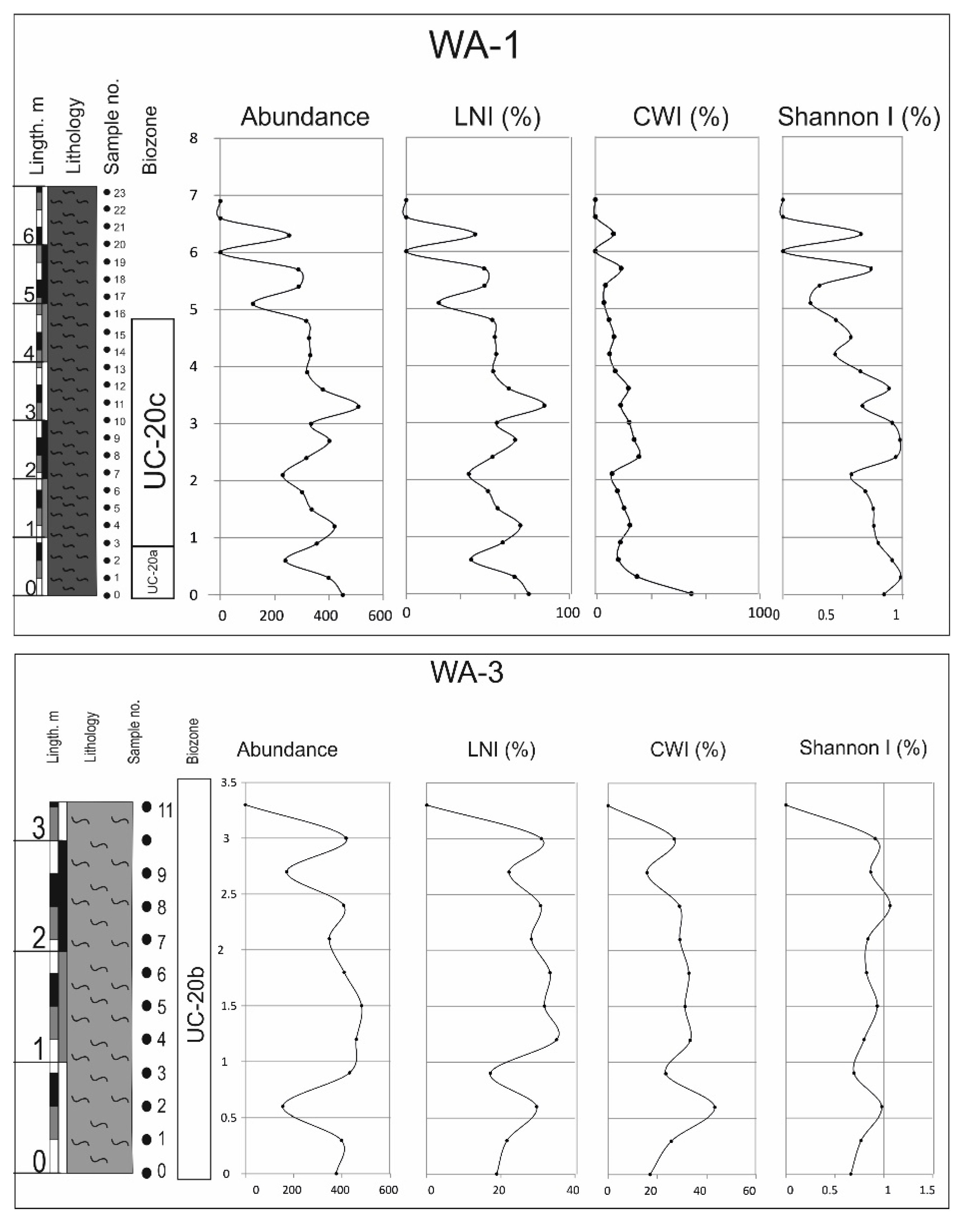
4.2.2. Thicknesses and Preservation
4.3. Continuity of Stratigraphic Sections
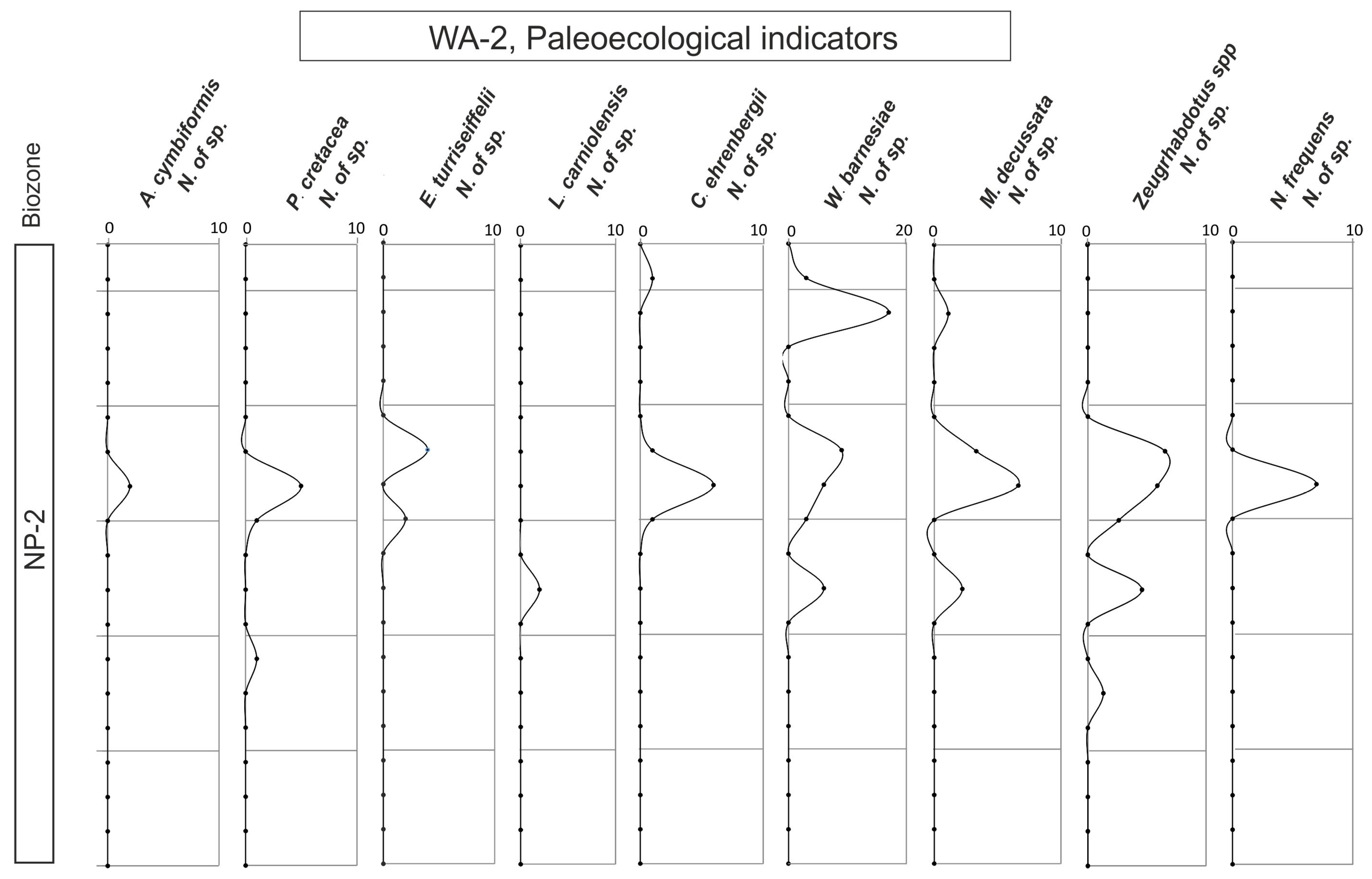
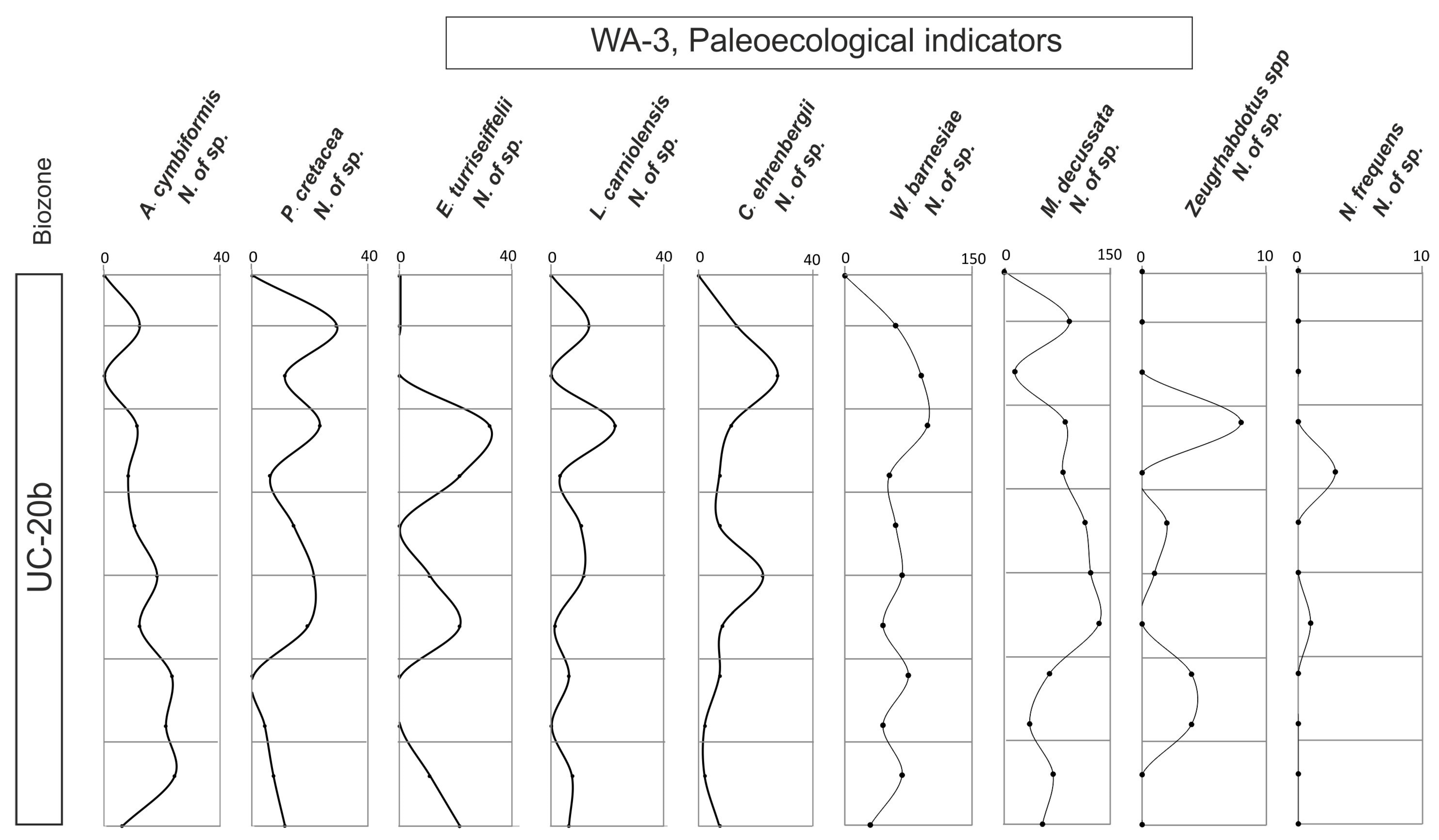
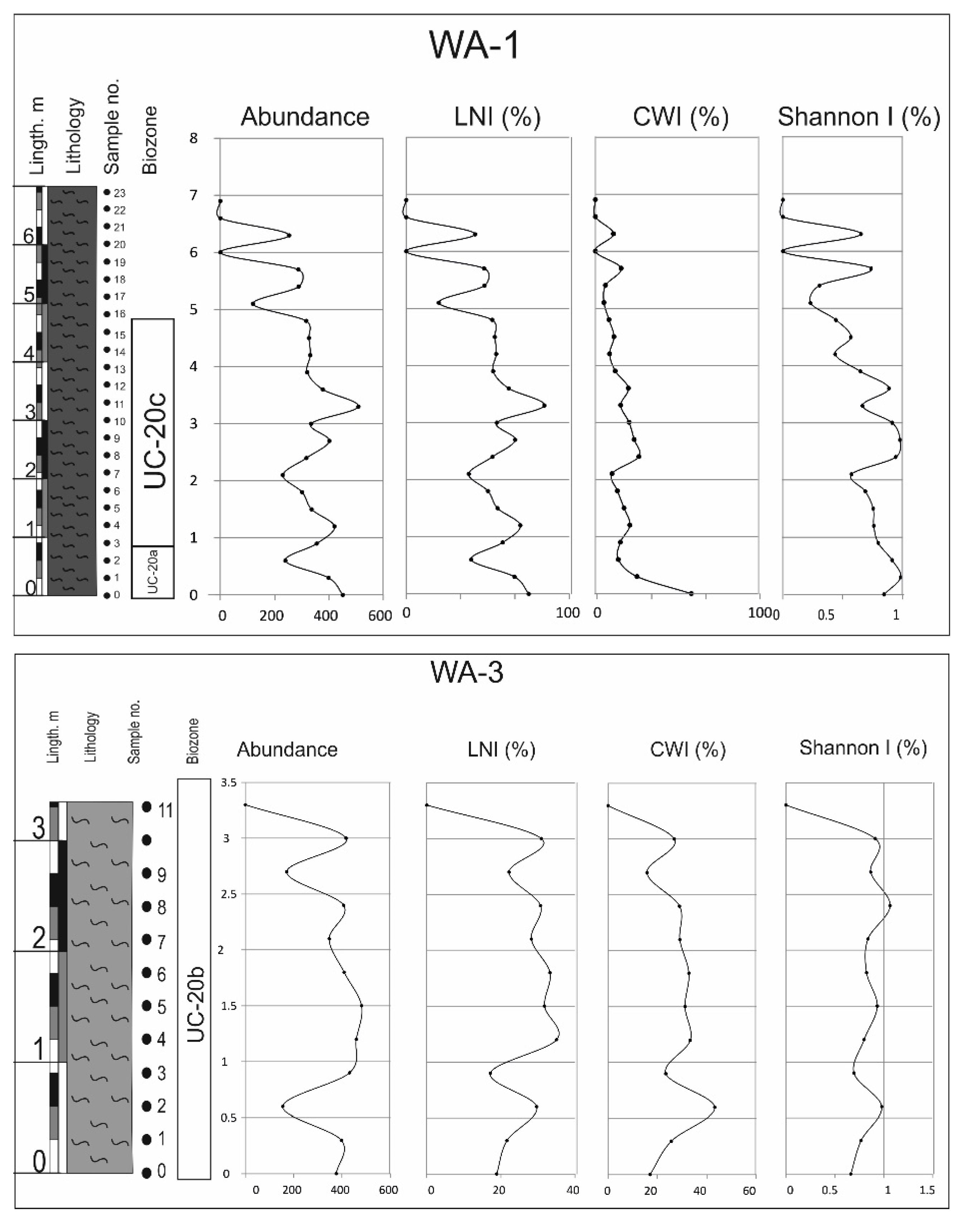
4.4. Ecological Indicators
5. Discussion
5.1. Age Framework
5.2. Sedimentation Rates
5.3. Stratigraphy and Structural Model
5.4. Ecological Model and Oceanographic Implication
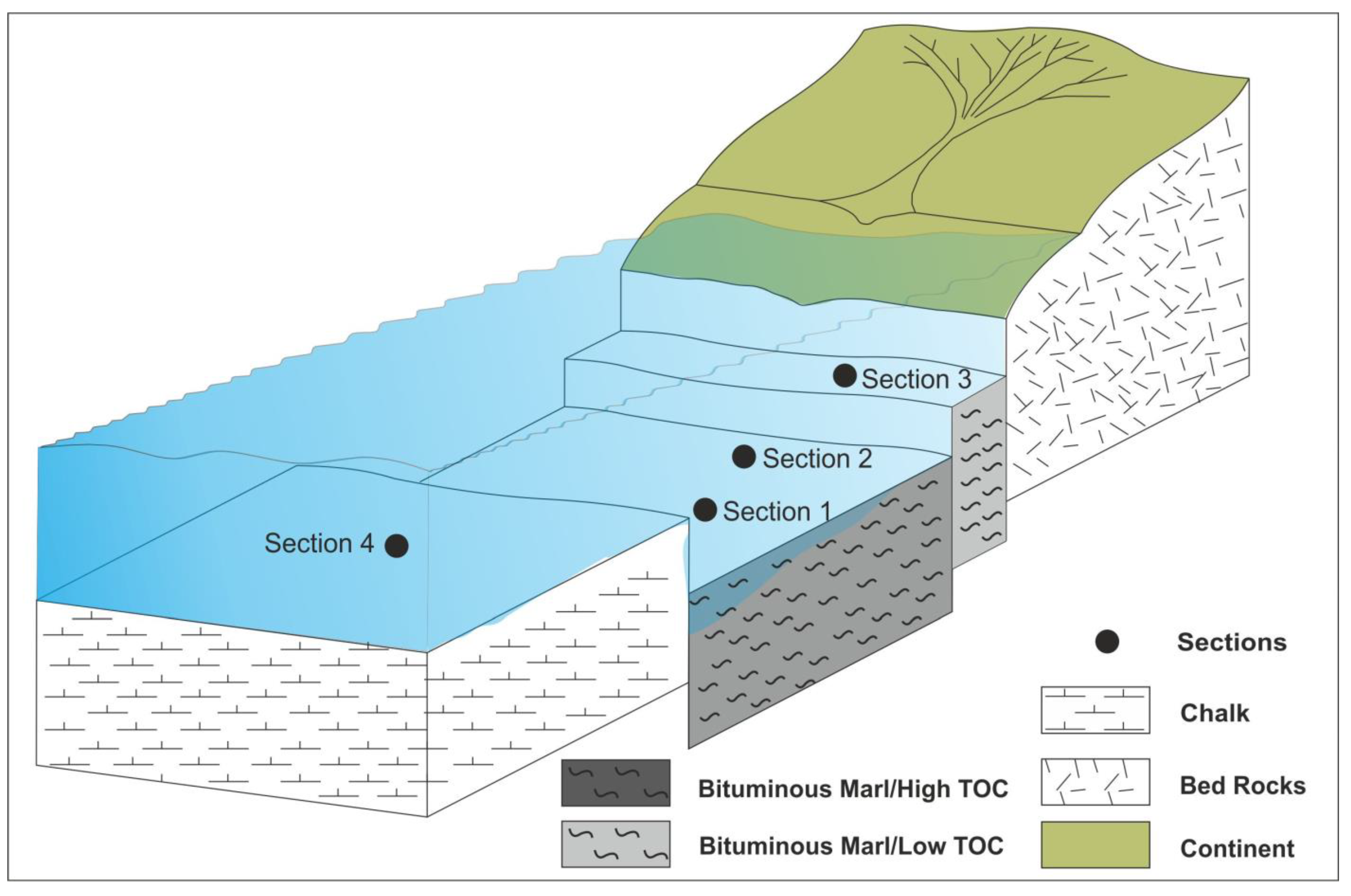
5.5. Basin Restriction and Organic Matter Preservation
6. Conclusions
- The MCM formation demonstrates a richness in calcareous nannofossils, as evidenced by the presence of numerous marker species, including L. quadratus, M. murus, M. prinsii, and Cr. tenuis in the examined sections.
- Based on the nannofossil marker species, the age assignment of the oil shale and chalk successions (WA-1, WA-2, WA-3, KAS-1) corresponds to the Late Cretaceous–Paleogene period, delineated within the following biozones in chronological sequence: UC-20a, UC-20b, UC-20c, UC-20d, and NP-2.
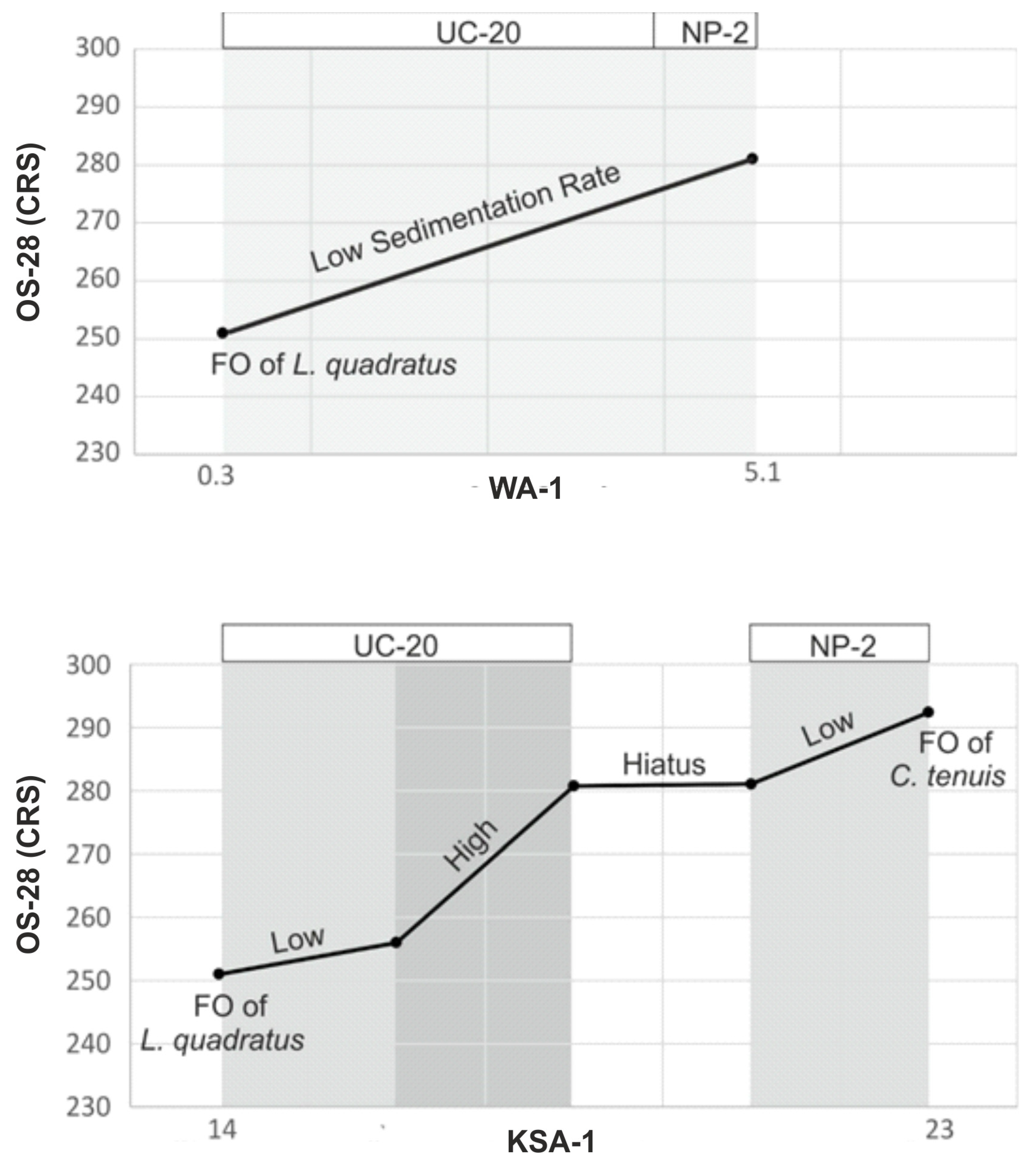
- The study identifies two hiatus intervals within section WA-1 and one within KAS-1.
- The application of a semi-quantitative method of correlation reveals that the UC-20 interval commenced with a low sedimentation rate, subsequently intensifying due to the acceleration of topographic changes between geological blocks.
- The UC-20C biozone reveals a clear trend toward warming and nutrient enrichment. This trend is linked to the presence of abundant and diverse species, and it aligns with wet seasons marked by an intensified continental influx into the subbasin. Consequently, nutrient levels are enhanced, facilitating the existence of diverse calcareous nannofossil assemblages.
- During these wet periods, terrigenous inputs transported nutrients, sustaining thriving marine ecosystems. High nutrient indicator species thrived in conjunction with the abundant nutrient-rich fossils.
- In the fourth section, a higher sampling interval is necessitated by the steep slope, posing a constraint on this research.
- Geochemical analyses could offer valuable insights for establishing correlations with calcareous nannofossil data, particularly in cases where certain elements exhibit strong associations with terrigenous input.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Herrle, J.O. Reconstructing nutricline dynamics of mid-Cretaceous oceans: Evidence from calcareous nannofossils from the Niveau Paquier black shale (SE France). Mar. Micropaleontol. 2003, 47, 307–321. [Google Scholar] [CrossRef]
- Bown, P. Calcareous Nannofossil Biostratigraphy; British Micropalaeontological Society Publication Series; Chapman and Hall: London, UK, 1998. [Google Scholar]
- Thierstein, H.R. Tentative Lower Cretaceous calcareous nannoplankton zonation. Eclogae Geol. Helv. 1971, 64, 458–488. [Google Scholar]
- Mutterlose, J.; Wise, S.W., Jr. Lower Cretaceous Nannofossil Biostratigraphy of ODP Leg 113 Holes 692B and 693A, Continental Slope off East Antarctica, Weddell Sea. Proc. Ocean. Drill. Program Sci. Res. 1990, 113, 689–697. [Google Scholar]
- Bender, F. Geology of the Arabian Peninsula, Jordan; United States Department of the Interior, Geological Survey: Reston, VA, USA, 1975. [Google Scholar]
- Powell, J.H. Stratigraphy and Sedimentation of the Phanerozoic Rocks in Central and Southern Jordan; Part B: Kurnub, Ajlun and Belqa Groups; Bulletin 11; Geology Directorate, Natural Resources Authority (Ministry of Energy and Mineral Resources): Amman, Jordan, 1989; 161p. [Google Scholar]
- Abed, A.M.; Yaghan, R. On the Paleoclimate of Jordan during the Last Glacial Maximum. Paleogeogr. Palaeoclimatol. Palaeoecol. 2000, 160, 23–33. [Google Scholar] [CrossRef]
- Yassini, I. Maastrichtian-Lower Eocene Biostratigraphy and the Planktonic Foraminiferal Biozonation in Jordan. Rev. Esp. Micropaleontol. 1979, 11, 5–57. [Google Scholar]
- Powell, J.H.; Ibrahim, K.; Rashdan, M. Jabal Khureij: A reinterpretation of sedimentation and tectonics (Upper Cretaceous and Tertiary) in the Dead Sea-Gulf of Aqaba Rift, Jordan. In Proceedings of the 3rd Jordanian Geologists’ Conference, Abstracts, Amman, Jordan; 1988. [Google Scholar]
- Powell, J.H.; Moh’d, B.K. Evolution of Cretaceous to Eocene Alluvial and Carbonate Platform Sequences in Central and South Jordan. GeoArabia 2011, 16, 29–82. [Google Scholar] [CrossRef]
- Abed, A.M.; Amireh, B.S. Petrography and Geochemistry of Some Jordanian Oil Shales from North Jordan. J. Pet. Geol. 1983, 5, 261–274. [Google Scholar] [CrossRef]
- Abed, A.M. The Eastern Mediterranean Phosphorite Giants: An Interplay between Tectonics and Upwelling. GeoArabia 2013, 18, 67–94. [Google Scholar] [CrossRef]
- Alqudah, M.; Hussein, M.A.; van den Boorn, S.; Giraldo, V.M.; Kolonic, S.; Podlaha, O.G.; Mutterlose, J. Eocene oil shales from Jordan—Paleoenvironmental implications from reworked microfossils. Mar. Pet. Geol. 2014, 52, 93–106. [Google Scholar] [CrossRef]
- Farouk, S.; Marzouk, A.M.; Ahmad, F. The Cretaceous/Paleogene boundary in Jordan. J. Asian Earth Sci. 2014, 94, 113–125. [Google Scholar] [CrossRef]
- Giraldo-Gómez, V.M.; Mutterlose, J.; Beik, I.; Podlaha, O.G.; Kolonic, S. Oil shales from the K-Pg boundary interval of Jordan—Climate controlled archives of surface and bottom water conditions in a shelf setting. Mar. Pet. Geol. 2021, 123, 104724. [Google Scholar] [CrossRef]
- Farouk, S.; Ahmad, F.; Baioumy, H.; Lehmann, B.; Mohammed, I.Q.; Al-Kahtany, K. Geochemical Characteristics of Carbonaceous Chalk near the Cretaceous/Paleogene Transition, Central Jordan: Strong Metal Enrichment of Redox-Sensitive and Biophile Elements from Remineralized Calcitic Plankton. Mar. Pet. Geol. 2020, 120, 104535. [Google Scholar] [CrossRef]
- Alhejoj, I.; Farouk, S.; Bazeen, Y.S.; Ahmad, F. Depositional Sequences and Sea-Level Changes of the Upper Maastrichtian-Middle Eocene Succession in Central Jordan: Evidence from Foraminiferal Biostratigraphy and Paleoenvironments. J. Afr. Earth Sci. 2020, 161, 103663. [Google Scholar] [CrossRef]
- Gradstein, F.M.; Ogg, J.G.; Schmitz, M.D.; Ogg, G.M. (Eds.) Geologic Time Scale; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar] [CrossRef]
- Moh’d. Geological Map of Irbid. Nat. Mapp. Map Sheet 3155 II Bull 46 Program 1997. [Google Scholar]
- Bender, F. Geologie von Jordanien. Beitrag zur regionalen Geologie der Erde; v. 7; Bornträger Publication: Berlin, Germany, 1968; p. 230. [Google Scholar]
- Jaber, J.O.; Abu-Rahmeh, T.M.; Alawin, A.; Al-Kloub, N. Surface Retorting of Jordanian Oil Shale and Associated CO2 Emissions. Jordan J. Mech. Ind. Eng. 2010, 4, 591–596. [Google Scholar]
- Powell, J.H.; Moh’d, B.K. Early Diagenesis of Late Cretaceous Chalk-Chert-Phosphorite Hardgrounds in Jordan: Implications for Sedimentation on a Coniacian-Campanian Pelagic Ramp. GeoArabia 2012, 17, 17–38. [Google Scholar] [CrossRef]
- Dill, H.G.; Kus, J.; Abed, A.M.; Sachsenhofer, R.F.; Khair, H.A. Diagenetic and Epigenetic Alteration of Cretaceous to Paleogene Organic-Rich Sedimentary Successions in Northwestern Jordan, Typical of the Western Margin of the Arabian Plate. GeoArabia 2009, 14, 101–140. [Google Scholar] [CrossRef]
- Ali Hussein, M.; Alqudah, M.; Blessenohl, M.; Podlaha, O.G.; Mutterlose, J. Depositional environment of Late Cretaceous to Eocene organic-rich marls from Jordan. GeoArabia 2015, 20, 191–210. [Google Scholar] [CrossRef]
- Chaimov, T.A.; Barazangi, M.; Al-Saad, D.; Sawaf, T.; Gebran, A.L.I. Mesozoic and Cenozoic deformation inferred from seismic stratigraphy in the southwestern intracontinental Palmyride fold-thrust belt, Syria. Geol. Soc. Am. Bull. 1992, 104, 704–715. [Google Scholar] [CrossRef]
- Alqudah, M.; Hussein, M.A.; van den Boorn, S.; Podlaha, O.G.; Mutterlose, J. Biostratigraphy and depositional setting of Maastrichtian–Eocene oil shales from Jordan. Mar. Pet. Geol. 2015, 60, 87–104. [Google Scholar] [CrossRef]
- Armstrong, H.A.; Brasier, M.D. Microfossil, stable isotopes and ocean-atmosphere history. Microfossils 2005, 2, 25–34. [Google Scholar] [CrossRef]
- Burnett, J.A.; Gallagher, L.T.; Hampton, M.J. Upper cretaceous. In Calcareous Nannofossil Biostratigraphy; Springer: Heidelberg, The Netherlands, 1998; pp. 132–199. [Google Scholar] [CrossRef]
- Thibault, N.; Gardin, S.; Galbrun, B. Latitudinal migration of calcareous nannofossil Micula murus in the Maastrichtian: Implications for global climate change. Geology 2010, 38, 203–206. [Google Scholar] [CrossRef]
- Butterlin, J.; Vrielynck, B.; Bignot, G.; Clermonte, J.; Colchen, M.; Dercourt, J.; Guiraud, R.; Poisson, A.; Ricou, L. Lutetian, (46–40 Ma). In Atlas of Tethys Palaeoenvironmental Maps; Dercourt, J., Ricou, L., Vrirlynck, B., Eds.; Explanatory Notes; Gauthier-Villars: Paris, France, 1993; pp. 197–209. [Google Scholar]
- Zachos, J.; Dickens, G.; Zeebe, R. An Early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature 2008, 451, 279–283. [Google Scholar] [CrossRef]
- Broggi, J.A. “Jacob Staff” and Measurements of Stratigraphic Sequences—A Correction: ERRATUM. AAPG Bull. 1946, 30, 1934. [Google Scholar]
- Bordiga, M.; Henderiks, J.; Tori, F.; Monechi, S.; Fenero, R.; Thomas, E. The Eocene–Oligocene Transition at ODP Site 1263, Atlantic Ocean: Decreases in Nannoplankton Size and Abundance and Correlation with Benthic Foraminiferal Assemblages. Clim. Past Discuss. 2015, 11, 1615–1664. [Google Scholar] [CrossRef]
- Martini, E. Standard Paleogene calcareous nannoplankton zonation. Nature 1970, 226, 560–561. [Google Scholar] [CrossRef]
- Roth, P.H. Jurassic and lower cretaceous calcareous nannofossils in the western north-atlantic (site-534)-biostratigraphy, preservation, and some observations on biogeography and paleoceanography. Initial. Rep. Deep. Sea Drill. Proj. 1983, 76, 587–621. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Mutterlose, J. Calcareous nannofossil palaeoceanography of the Early Cretaceous of NW Europe. Mitteilungen Geol. Staatsinst. Hambg. 1996, 77, 291–313. [Google Scholar]
- Aizawa, C.; Oba, T.; Okada, H. Late Quaternary Paleoceanography Deduced from Coccolith Assemblages in a Piston Core Recovered off the Central Japan Coast. Mar. Micropaleontol. 2004, 52, 277–297. [Google Scholar] [CrossRef]
- Show-yung, H. Carnosaurian remains from Alashan, inner Mongolia. Certebrata Palasiat 1964, 8, 42–63. [Google Scholar]
- Mutterlose, J.; Bornemann, A.; Herrle, J.O. Mesozoic calcareous nannofossils-state of art. Paläontol. Z. 2005, 79, 113–133. [Google Scholar] [CrossRef]
- Alhejoj, I.; Faris, M.; Ahmad, F.; Farouk, S.; Sreepat, J.; Musa, G.; Shama, M. Calcareous Nannofossil Biostratigraphy and Bioevents across the Ypresian/Lutetian Boundary in Egypt and Jordan. J. Afr. Earth Sci. 2023, 201, 104881. [Google Scholar] [CrossRef]
- Bazeen, Y.S.; Farouk, S.; El-Balkiemy, A.F.; Ahmad, F. Late Maastrichtian-early Danian sealevel changes at Gebel Tawil Shihaq in Central Jordan: Implications of chemostratigraphy and foraminiferal biostratigraphy. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 556, 109875. [Google Scholar] [CrossRef]
- Smadi, A. Calcareous nannofossils and planktonic foraminifera among KT boundary in Azraq, Jordan. Dirasat Pure Sci. 2002, 29, 71–88. [Google Scholar]
- Haq, B.U.; Vail, P.R.; Hardenbol, J.; Van Wagoner, J.C.; Matthews, R.K. Sea Level History: Response. Science 1988, 241, 596–599. [Google Scholar] [CrossRef] [PubMed]
- Cloetingh, S.; Haq, B.U. Inherited Landscapes and Sea Level Change. Science 2015, 347, 1258375. [Google Scholar] [CrossRef]
- Miller, K.G.; Schmelz, W.J.; Browning, J.V.; Kopp, R.E.; Mountain, G.S.; Wright, J.D. Ancient Sea Level. Oceanography 2020, 33, 32–41. [Google Scholar] [CrossRef]
- Al-Soud, H.; Al-Mashakbeh, H. Depositional Environment, Microfacies Analysis, and Planktonic Foraminifera of Oil Shale Deposits in the Wadi Al-Shallala Area, NW Jordan. JJEES 2021, 12, 113–121. [Google Scholar]
- Abu-Mahfouz, I.S.; Wicaksono, A.N.; Idiz, E.; Cartwright, J.; Santamarina, J.C.; Vahrenkamp, V.C. Modelling the Initiation of Bitumen-Filled Microfractures in Immature, Organic-Rich Carbonate Mudrocks: The Maastrichtian Source Rocks of Jordan. Mar. Pet. Geol. 2022, 141, 105700. [Google Scholar] [CrossRef]
- Alqudah, M.; Al Alaween, M.; Fermor, A.; Ali, E.; Wagner, T.; Alhakimi, M.H.; Rahim, A. Impact of Thermal Maturation of the Upper Cretaceous Bituminous Limestone of Attarat Um Ghudran Central Jordan on Calcareous Nannofossil Preservation. ACS Omega 2023, 42, 39830–39846. [Google Scholar] [CrossRef]
- Lees, J.A. Calcareous nannofossil biogeography illustrates palaeoclimate change in the Late Cretaceous Indian Ocean. Cretac. Res. 2002, 23, 537–634. [Google Scholar] [CrossRef]
- Thibault, N.; Gardin, S. Maastrichtian calcareous nannofossil biostratigraphy and paleoecology in the Equatorial Atlantic (Demerara Rise, ODP Leg 207 Hole 1258A). Rev. Micropaléontol. 2006, 49, 199–214. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussein, A.; Al-Tarawneh, O.M.; Alqudah, M. Calcareous Nannofossils Biostratigraphy of Late Cretaceous–Paleocene Successions from Northern Jordan and Their Implications for Basin Analysis. Geosciences 2023, 13, 351. https://doi.org/10.3390/geosciences13110351
Hussein A, Al-Tarawneh OM, Alqudah M. Calcareous Nannofossils Biostratigraphy of Late Cretaceous–Paleocene Successions from Northern Jordan and Their Implications for Basin Analysis. Geosciences. 2023; 13(11):351. https://doi.org/10.3390/geosciences13110351
Chicago/Turabian StyleHussein, Albesher, Osama M. Al-Tarawneh, and Mohammad Alqudah. 2023. "Calcareous Nannofossils Biostratigraphy of Late Cretaceous–Paleocene Successions from Northern Jordan and Their Implications for Basin Analysis" Geosciences 13, no. 11: 351. https://doi.org/10.3390/geosciences13110351
APA StyleHussein, A., Al-Tarawneh, O. M., & Alqudah, M. (2023). Calcareous Nannofossils Biostratigraphy of Late Cretaceous–Paleocene Successions from Northern Jordan and Their Implications for Basin Analysis. Geosciences, 13(11), 351. https://doi.org/10.3390/geosciences13110351