

New Evidence for an Episode of Accelerated Environmental Change in the Late Barremian: Geochemical and Paleontological Records from the Subbetic Basin (Western Tethys)

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
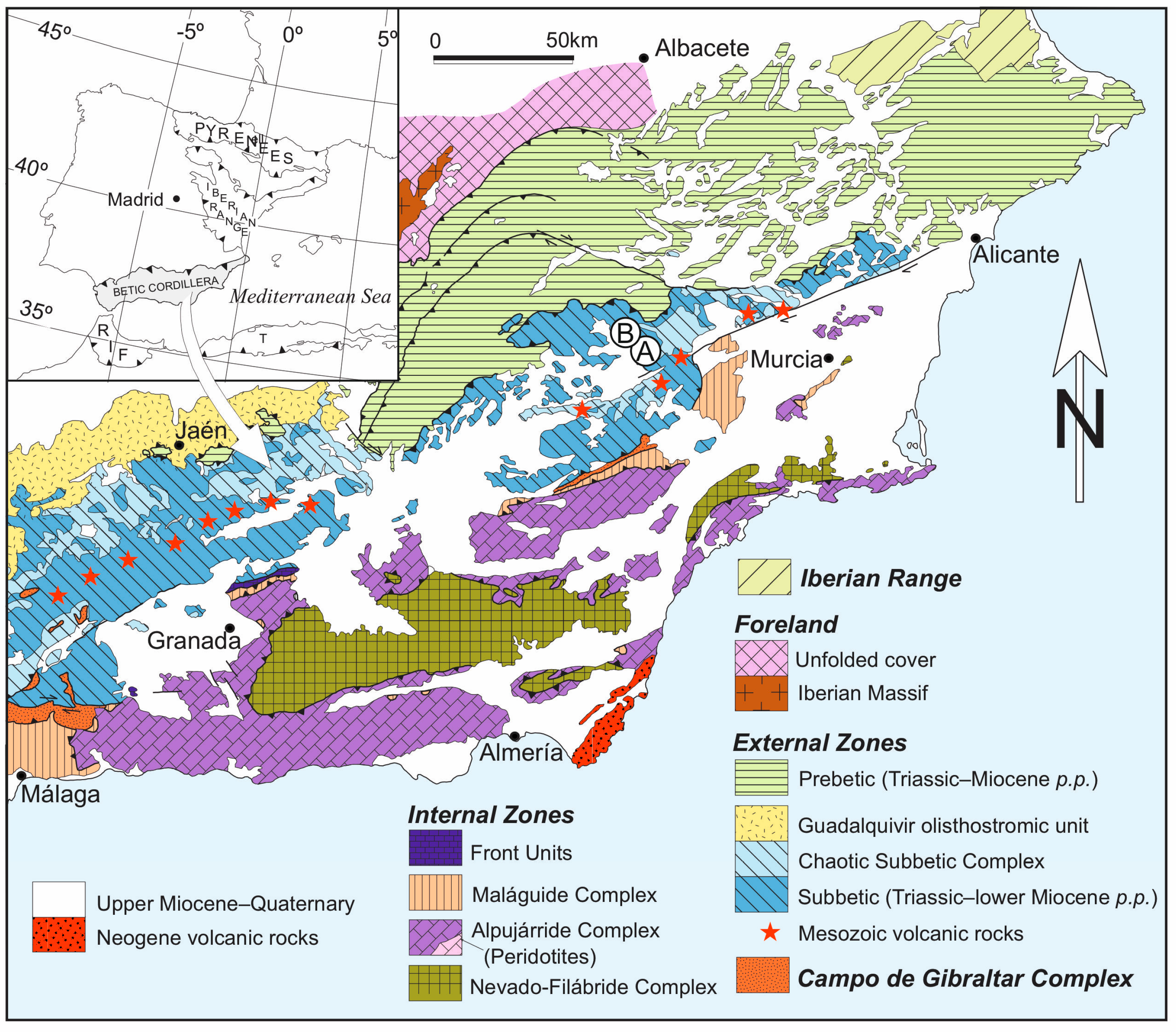
2.1. Geological Background
2.2. Methods
3. Results
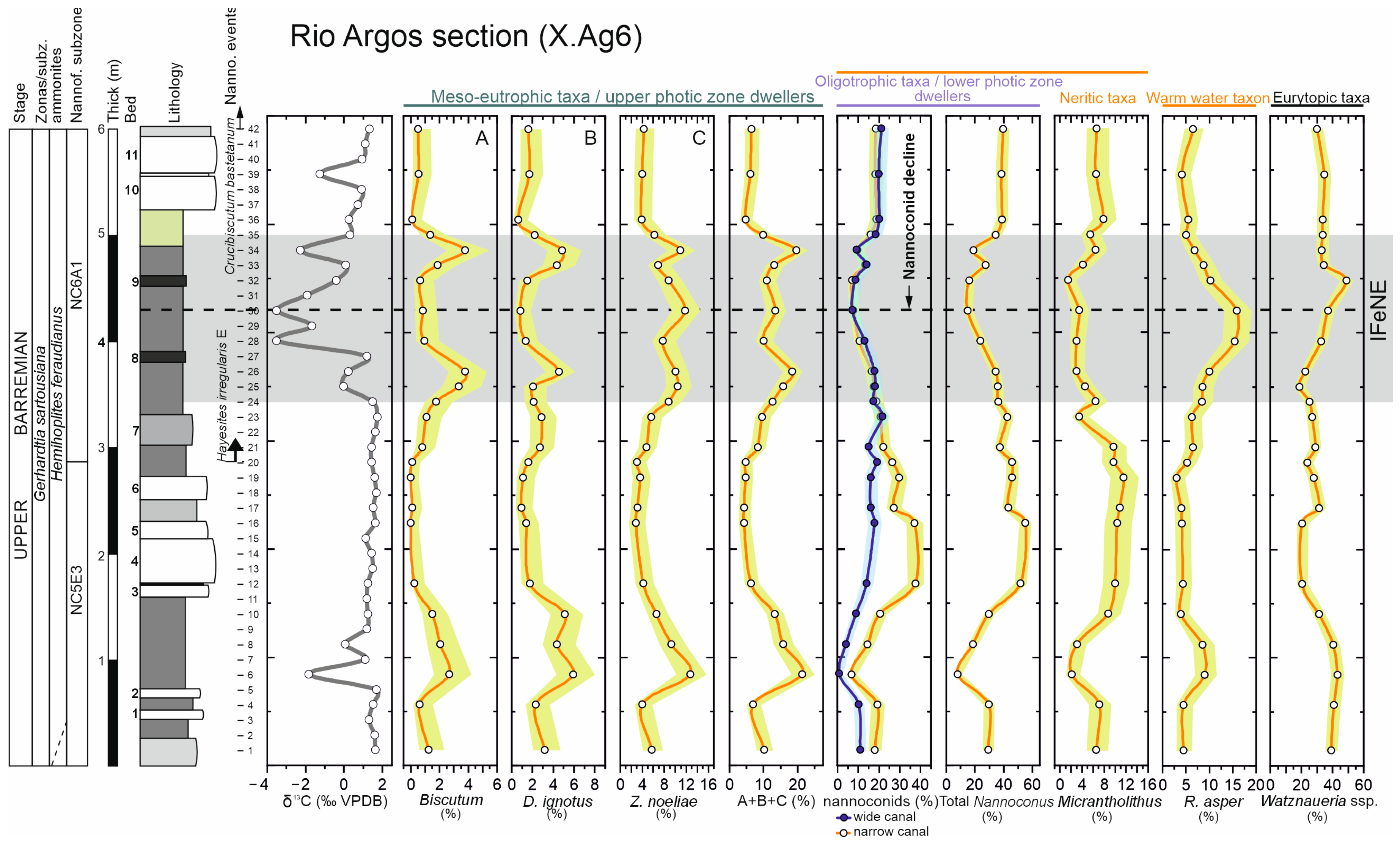
3.1. Calcareous Nannofossil Assemblages
3.2. Geochemistry
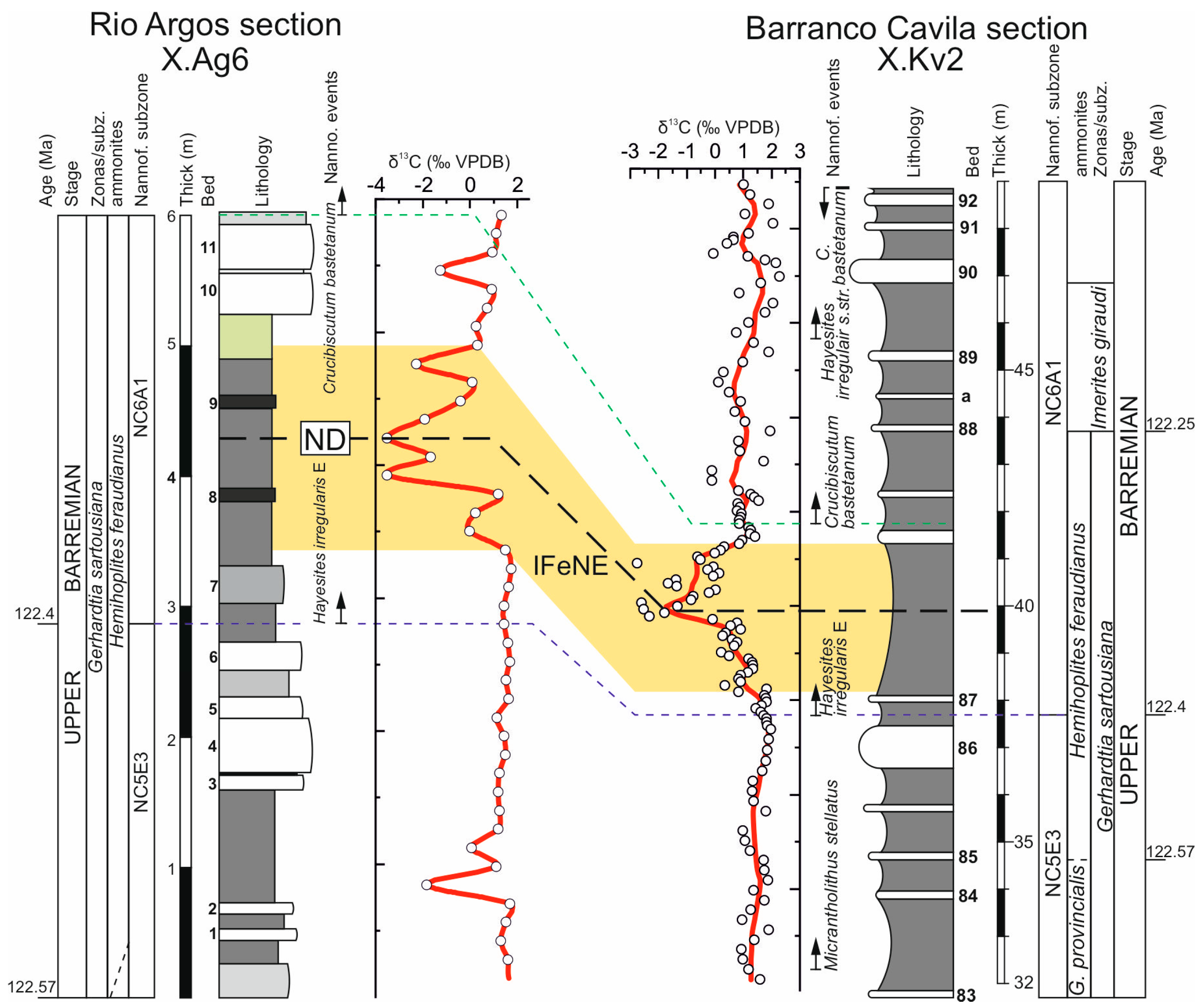
3.2.1. C-Isotope Stratigraphy
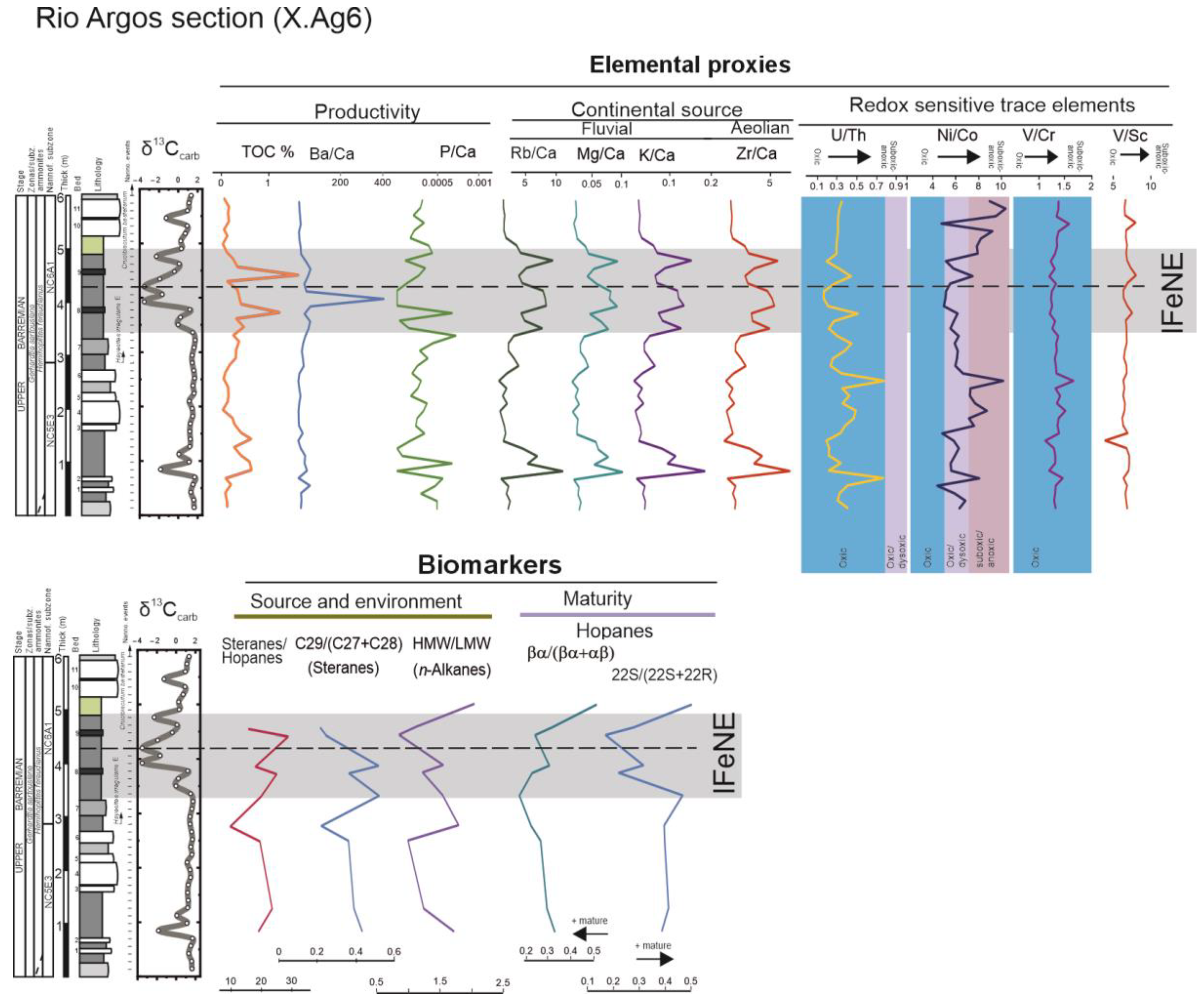
3.2.2. Elemental Geochemistry
3.2.3. Biomarkers
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jenkyns, H.C. Geochemistry of oceanic anoxic events. Geochem. Geophys. Geosyst. 2010, 11, Q03004. [Google Scholar] [CrossRef]
- Föllmi, K.B. Early Cretaceous life, climate and anoxia. Cretac. Res. 2012, 35, 230–257. [Google Scholar] [CrossRef]
- Erba, E.; Duncan, R.A.; Bottini, C.; Tiraboschi, D.; Weissert, H.; Jenkyns, H.C.; Malinverno, A. Environmental consequences of Ontong Java Plateau and Kerguelen Plateau volcanism. In The Origin, Evolution, and Environmental Impact of Oceanic Large Igneous Provinces; Neal, C.R., Sager, W.W., Sano, T., Erba, Y.E., Eds.; Geological Society of America Special Paper 511; Geological Society of America: Boulder, CO, USA, 2015. [Google Scholar]
- Castro, J.M.; Ruiz-Ortiz, P.A.; de Gea, G.A.; Aguado, R.; Jarvis, I.; Weissert, H.J.; Molina, J.M.; Nieto, L.M.; Pancost, R.D.; Quijano, M.L.; et al. High-resolution C-isotope, TOC and biostratigraphic records of OAE 1a (Aptian) from an expanded hemipelagic cored succession, western Tethys: A new stratigraphic reference for global correlation and paleoenvironmental reconstruction. Paleoceanogr. Paleoclimatol. 2021, 36, e2020PA004004. [Google Scholar] [CrossRef]
- Percival, L.M.E.; Matsumoto, H.; Callegaro, S.; Erba, E.; Ker, A.C. Cretaceous large igneous provinces: From volcanic formation to environmental catastrophes and biological crises. In Cretaceous Project 200 Volume 1: The Cretaceous World; Hart, M.B., Batenburg, S.J., Huber, B.T., Price, G.D., Thibault, N., Wagreich, M., Walaszczyk, I., Eds.; Special Publications, 544; Geological Society: London, UK, 2024. [Google Scholar] [CrossRef]
- Charbonnier, G.; Adatte, T.; Spangenberg, J.E.; Föllmi, K.B. The expression of early Aptian to latest Cenomanian oceanic anoxic events in the sedimentary record of the Briançonnais domain. Glob. Planet. Change 2018, 170, 76–92. [Google Scholar] [CrossRef]
- Martinez, M.; Aguirre-Urreta, B.; Dera, G.; Lescano, M.; Omarini, J.; Tunik, M.; O’Dogherty, L.; Aguado, R.; Company, M.; Bodin, S. Synchrony of carbon cycle fluctuations, volcanism and orbital forcing during the Early Cretaceous. Earth Sci. Rev. 2023, 239, 104356. [Google Scholar] [CrossRef]
- Pictec, A.; Tschanz, K.; Kürsteiner, O. Record of a dense succession of drowning phases in the Alpstein mountains, northeastern Switzerland: Part II—The Lower Cretaceous Schrattenkalk Formation (late Barremian). Swiss J. Geosci. 2023, 116, 1. [Google Scholar] [CrossRef]
- Aguado, R.; Company, M.; O’Dogherty, L.; Sandoval, J.; Martinez, M. New insights into the Barremian–lower Aptian calcareous nannofossils of the Mediterranean Tethys: Chronostratigraphic and paleobiogeographic implications. Mar. Micropaleontol. 2022, 173, 102114. [Google Scholar] [CrossRef]
- Vera, J.A. Cordillera Bética y Baleares. In Geología de España; Vera, J.A., Ed.; Sociedad Geológica de España—Instituto Geológico y Minero de España: Madrid, Spain, 2004; pp. 345–464. [Google Scholar]
- Martín-Chivelet, J.; López-Gómez, J.; Aguado, R.; Arias, C.; Arribas, J.; Arribas, M.E.; Aurell, M.; Bádenas, B.; Benito, M.I.; Bover-Arnal, T.; et al. The Late Jurassic–early cretaceous rifting. In The Geology of Iberia: A Geodynamic Approach. Volume 3: The Alpine Cycle; Quesada, C., Oliveira, J.T., Eds.; Regional Geology Reviews; Springer: Cham, Switzerland, 2019; pp. 169–249. [Google Scholar] [CrossRef]
- Cao, W.; Williams, S.; Flament, N.; Zahirovic, S.; Scotese, C.; Müller, R.D. Palaeolatitudinal distribution of lithologic indicators of climate in a palaeogeographic framework. Geol. Mag. 2018, 156, 331–354. [Google Scholar] [CrossRef]
- Masse, J.P.; Bellion, Y.; Benkhelil, J.; Boulin, J.; Cornee, J.J.; Dercourt, J.; Guiraud, R.; Mascle, G.; Poisson, A.; Ricou, L.E.; et al. Lower Aptian palaeoenvironments 114–112 Ma. In Atlas Tethys Palaeoenvironmental Maps; Dercourt, J., Ed.; Beceip-Franlab: Rueil-Malmaison, France, 1993. [Google Scholar]
- Barrier, E.; Vrielynck, B.; Brouillet, J.F.; Brunet, M.F. Paleotectonic Reconstruction of the Central Tethyan Realm; CCGM-CGMW: Paris, France, 2018. [Google Scholar]
- Ziegler, P.A. Geological Atlas of Western and Central Europe. In Shell Internationale Petroleum; Geological Society: London, UK, 1990. [Google Scholar]
- Molina, J.M. Análisis de Facies del Mesozoico en el Subbético Externo. (Prov. de Córdoba y sur de Jaén). Ph.D. Thesis, Universidad de Granada, Granada, Spain, 1987. [Google Scholar]
- Geisen, M.; Bollmann, J.; Herrle, J.O.; Mutterlose, J.; Young, J.R. Calibration of the Random Settling Technique for Calculation of Absolute Abundances of Calcareous Nannoplankton. Micropaleontology 1999, 45, 437–442. [Google Scholar] [CrossRef]
- Fatela, F.; Taborda, R. Confidence limits of species proportions in microfossil assemblages. Mar. Micropaleontol. 2002, 45, 169–174. [Google Scholar] [CrossRef]
- Suchéras-Marx, B.; Escarguel, G.; Ferreira, J.; Hammer, Ø. Statistical confidence intervals for relative abundances and abundance-based ratios: Simple practical solutions for an old overlooked question. Mar. Micropaleontol. 2019, 151, 101751. [Google Scholar] [CrossRef]
- Aguado, R.; de Gea, G.A.; Castro, J.M.; O’Dogherty, L.; Quijano, M.L.; Naafs, B.D.A.; Pancost, R.D. Late Barremian–early Aptian dark facies of the Subbetic (Betic Cordillera, southern Spain): Calcareous nannofossil quantitative analyses, chemostratigraphy and palaeoceanographic reconstructions. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 395, 198–221. [Google Scholar] [CrossRef]
- Calvert, S.E.; Pedersen, T.F. Chapter fourteen: Elemental proxies for palaeoclimatic and palaeoceanographic variability in marine sediments: Interpretation and application. Dev. Mar. Geol. 2007, 1, 567–644. [Google Scholar]
- Peters, K.E.; Walters, C.C.; Moldowan, J.M. The Biomarker Guide, 2nd ed.; Cambridge University Press: Cambridge, UK, 2005; 1155p. [Google Scholar]
- Martinez, M.; Aguado, R.; Company, M.; Sandoval, J.; O´Dogherty, L. Integrated astrochronology of the Barremian Stage (Early Cretaceous) and its biostratigraphic subdivisions. Glob. Planet. Change 2020, 195, 103368. [Google Scholar] [CrossRef]
- Eglinton, G.; Hamilton, R.J. Leaf epicuticular waxes. Science 1967, 156, 1322–1335. [Google Scholar] [CrossRef] [PubMed]
- Ourisson, G.; Albrecht, P.; Rohmer, M. Predictive microbial biochemistry from molecular fossils to prokaryotic membranes. Trends Biochem. Sci. 1982, 7, 236–239. [Google Scholar] [CrossRef]
- Volkman, J.K.; Barrett, S.M.; Blackburn, S.I.; Man-Sour, M.P.; Sikes, E.L.; Gelin, F. Microalgal biomarkers: A review of recent research developments. Org. Geochem. 1998, 29, 1163–1179. [Google Scholar] [CrossRef]
- Summons, R.E.; Jahnke, L.L.; Hope, J.M.; Logan, G.A. 2-Methylhopanoids as bio- markers for cyanobacterial oxygenic photosynthesis. Nature 1999, 400, 554–557. [Google Scholar] [CrossRef]
- Ricci, J.N.; Morton, R.; Kulkarni, G.; Summers, M.L.; Newman, D.K. Hopanoids play a role in stress tolerance and nutrient storage in the cyanobacterium Nostoc punctiforme. Geobiology 2016, 15, 173–183. [Google Scholar] [CrossRef]
- van Breugel, Y.; Schouten, S.; Tsikos, H.; Erba, E.; Price, G.D.; Sinninghe-Damsté, J.S. Synchronous negative carbon isotope shifts in marine and terrestrial biomarkers at the onset of the early Aptian oceanic anoxic event 1a: Evidence for the release of 13C-depleted carbon into the atmosphere. Paleoceanography 2007, 22, PA1210. [Google Scholar] [CrossRef]
- Castro, J.M.; de Gea, G.A.; Quijano, M.L.; Aguado, R.; Froehner, S.; Naafs, B.D.A.; Pancost, R.D. Complex and protracted environmental and ecological perturbations during OAE 1a—Evidence from an expanded pelagic section from south Spain (Western Tethys). Glob. Planet. Change 2019, 183, 103030. [Google Scholar] [CrossRef]
- Huang, W.-Y.; Meinschein, W.G. Sterols as ecological indicators. Geochim. Cosmochim. Acta 1979, 43, 739–745. [Google Scholar] [CrossRef]
- Mahoney, J.J.; Storey, M.; Duncan, R.A.; Spencer, K.J.; Pringle, M. Geochemistry and age of the Ontong Java Plateau. In The Mesozoic Pacific: Geology, Tectonics, and Volcanism; AGU Geophysical Monograph Series; American Geophysical Union: Washington, DC, USA, 1993; Volume 77, pp. 233–261. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Gea, G.A.; Castro, J.M.; Company, M.; O’Dogherty, L.; Sandoval, J.; Quijano, M.L.; Sequero, C.; Froehner, S.; Aguado, R. New Evidence for an Episode of Accelerated Environmental Change in the Late Barremian: Geochemical and Paleontological Records from the Subbetic Basin (Western Tethys). Geosciences 2024, 14, 187. https://doi.org/10.3390/geosciences14070187
de Gea GA, Castro JM, Company M, O’Dogherty L, Sandoval J, Quijano ML, Sequero C, Froehner S, Aguado R. New Evidence for an Episode of Accelerated Environmental Change in the Late Barremian: Geochemical and Paleontological Records from the Subbetic Basin (Western Tethys). Geosciences. 2024; 14(7):187. https://doi.org/10.3390/geosciences14070187
Chicago/Turabian Stylede Gea, Ginés A., José Manuel Castro, Miguel Company, Luis O’Dogherty, José Sandoval, María Luisa Quijano, Cristina Sequero, Sandro Froehner, and Roque Aguado. 2024. "New Evidence for an Episode of Accelerated Environmental Change in the Late Barremian: Geochemical and Paleontological Records from the Subbetic Basin (Western Tethys)" Geosciences 14, no. 7: 187. https://doi.org/10.3390/geosciences14070187
APA Stylede Gea, G. A., Castro, J. M., Company, M., O’Dogherty, L., Sandoval, J., Quijano, M. L., Sequero, C., Froehner, S., & Aguado, R. (2024). New Evidence for an Episode of Accelerated Environmental Change in the Late Barremian: Geochemical and Paleontological Records from the Subbetic Basin (Western Tethys). Geosciences, 14(7), 187. https://doi.org/10.3390/geosciences14070187