
Svalbard Fjord Sediments as a Hotspot of Functional Diversity and a Reservoir of Antibiotic Resistance
 , , and
, , and
Abstract
:1. Introduction
- (i)
- Since there were differences in the intensity of human activity and the sediment pollution within the selected fjords [45], a higher abundance of antibiotic-resistant bacteria was expected to be found within the benthic microflora in human-impacted areas (e.g., Adventfjorden);
- (ii)
- As a glacier–marine transition occurs from the inner part to the outer part of the fjords, we hypothesized that this spatial gradient would be reflected in a greater complexity of carbon substrate utilization, with a potential shift from simple to complex carbon metabolism when moving from glacier to open-sea environments.
2. Materials and Methods
2.1. Study Area and Sample Collection
2.2. Heterotrophic (Marine and Non-Marine) Bacterial Abundance
2.3. Extracellular Enzymatic Activities (Leucine Aminopeptidase, LAP, Beta-Glucosidase, GLU, and Alkaline Phosphatase, AP)
2.4. Community-Level Physiological Profiles (CLPPs) via Biolog Ecoplates
2.5. Phenotypic and Biochemical Characterization of Bacterial Isolates
2.6. Antibiotic Susceptibility Profiles of Bacterial Isolates
- -
- Cell wall antibiotics such as beta-lactams, including penicillins [ampicillin (10 μg, code CT0003B)];
- -
- Nucleic acid inhibitors, including fluoroquinolones [enrofloxacin (5 μg, CT0639B)];
- -
- Protein synthesis inhibitors, including (1) aminoglycoside antibiotics [gentamycin (10 μg, CT0024B)], (2) lincosamides [clindamycin (2 μg, CT0064B)], and (3) tetracyclines [tetracycline (30 μg, CT0054B)].
2.7. Statistical Analyses
3. Results
3.1. Culturable Heterotrophic Bacterial Abundance and Organic Matter Decomposition by Enzyme Activities
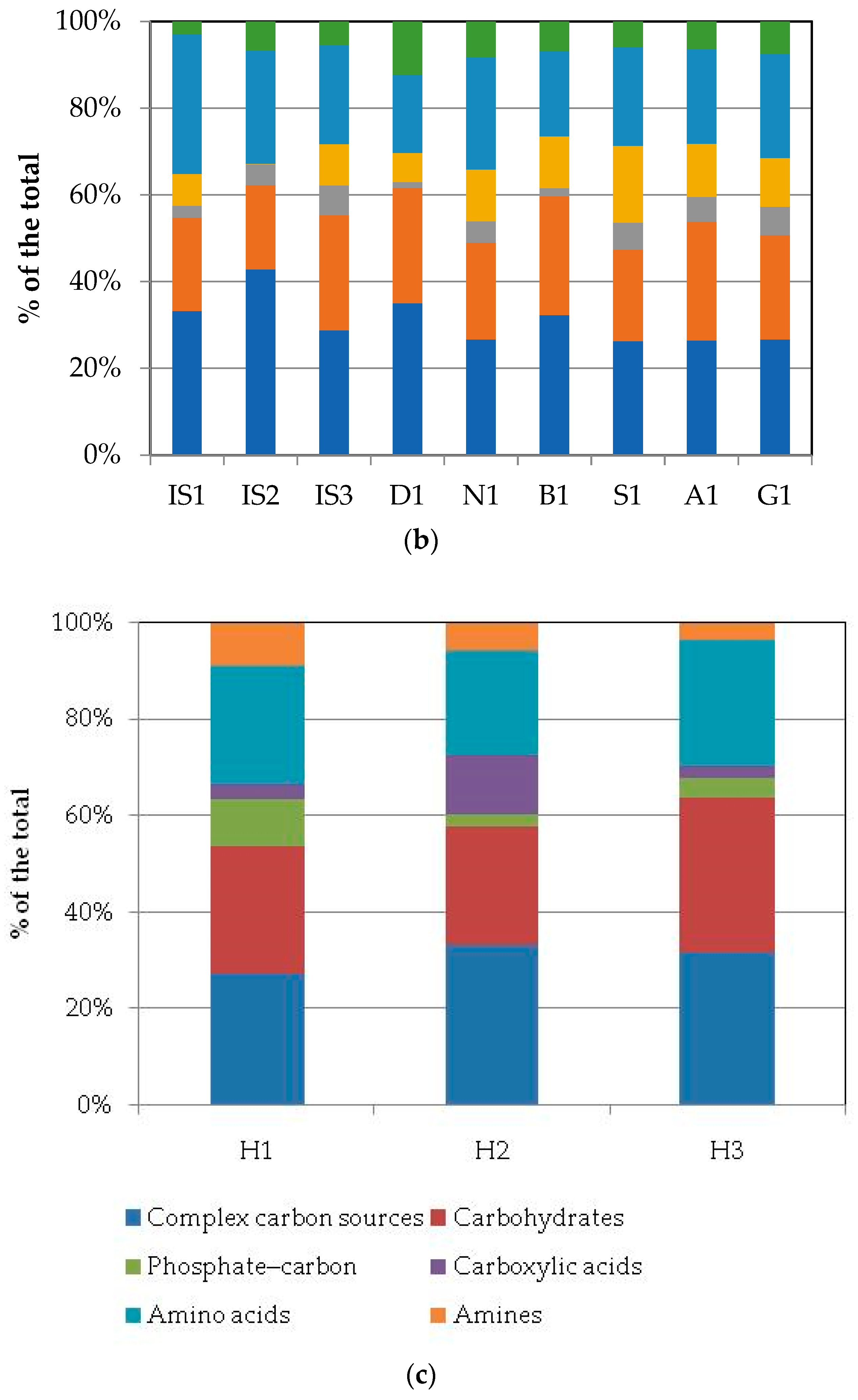
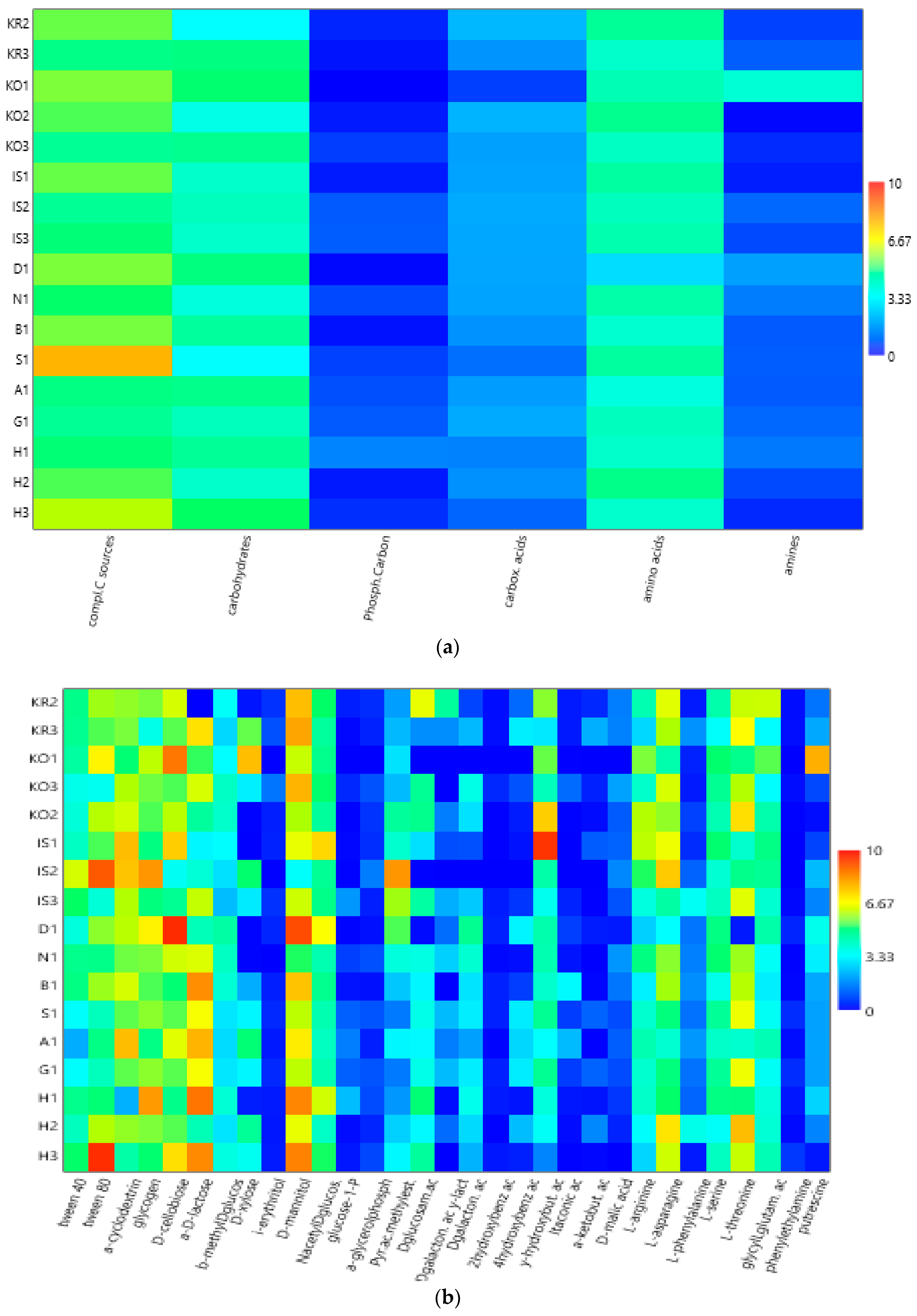
3.2. Community-Level Physiological Profiles (CLPPs) via Biolog Ecoplates
3.3. Statistical Elaboration of the Whole Abundance and Metabolic Dataset
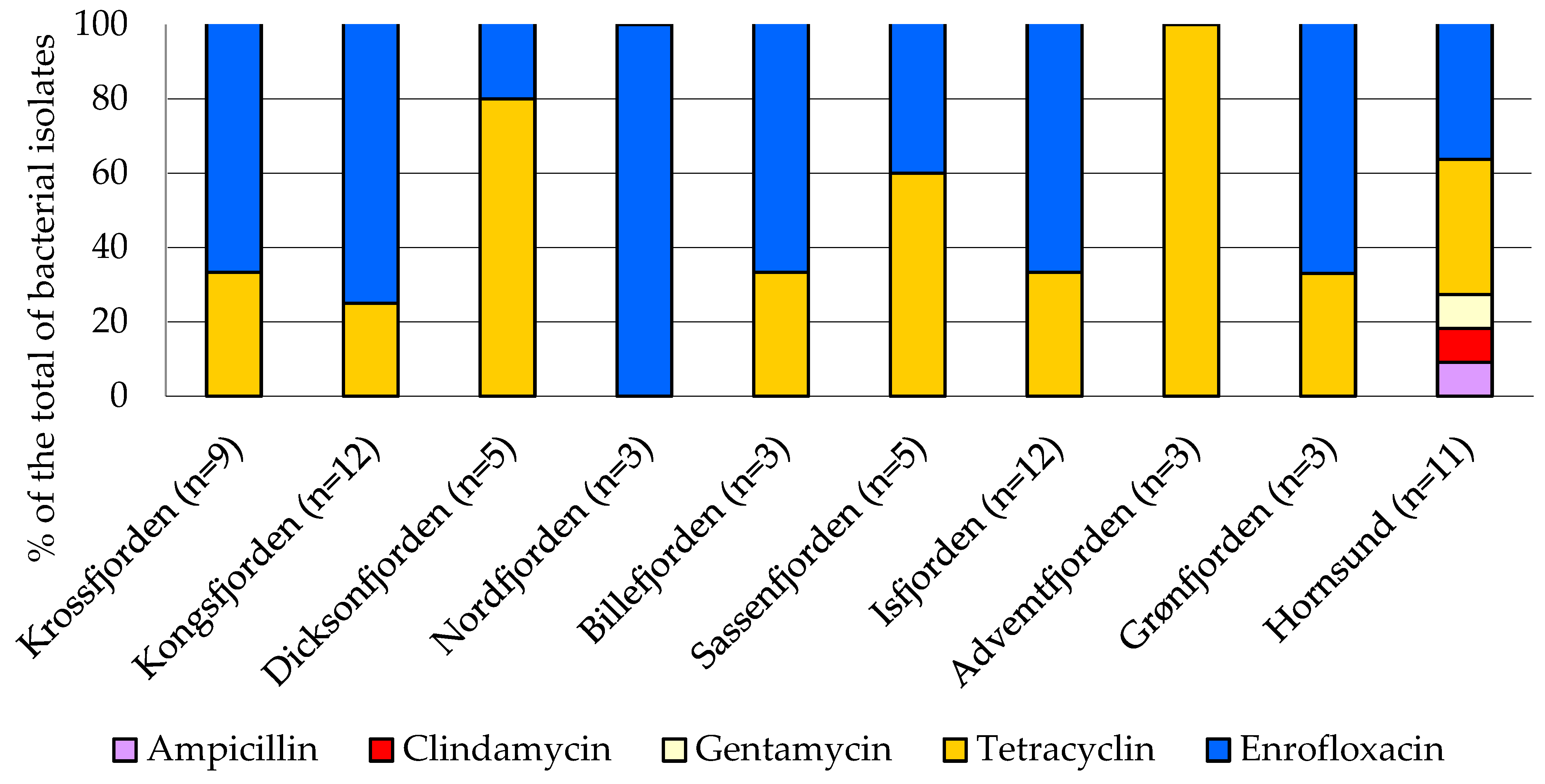
3.4. Antibiotic Susceptibility Profiles of Bacterial Isolates
4. Discussion
4.1. Culturable Heterotrophic Bacterial Abundance and Organic Matter Decomposition by Enzyme Activities
4.2. Microbial Community Metabolism (CLPPs)
4.3. Antibiotic-Resistant Bacteria
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Caruso, G.; Madonia, A.; Bonamano, S.; Miserocchi, S.; Giglio, F.; Maimone, G.; Azzaro, F.; Decembrini, F.; La Ferla, R.; Piermattei, V.; et al. Microbial Abundance and Enzyme Activity Patterns: Response to Changing Environmental Characteristics along a Transect in Kongsfjorden (Svalbard Islands). J. Mar. Sci. Eng. 2020, 8, 824. [Google Scholar] [CrossRef]
- Svendsen, H.; Beszczynska-Møller, A.; Hagen, J.O.; Lefauconnier, B.; Tverberg, V.; Gerland, S.; Ørbaek, J.B.; Bischof, K.; Papucci, C.; Zajaczkowski, M.; et al. The physical environment of Kongsfjorden–Krossfjorden, an Arctic fjord system in Svalbard. Polar Res. 2002, 21, 133–166. [Google Scholar] [CrossRef]
- Pelikan, C.; Jaussi, M.; Wasmund, K.; Seidenkrantz, M.-S.; Pearce, C.; Kuzyk, Z.Z.A.; Herbold, C.W.; Røy, H.; Kjeldsen, K.U.; Loy, A. Glacial runoff promotes deep burial of sulfur cycling-associated microorganisms in marine sediments. Front. Microbiol. 2019, 10, 2558. [Google Scholar] [CrossRef]
- Tobias-Hünefeldt, S.P.; Wing, S.R.; Baltar, F.; Morales, S.E. Changes in microbial community phylogeny and metabolic activity along the water column uncouple at near sediment aphotic layers in fjords. Sci. Rep. 2021, 11, 19303. [Google Scholar] [CrossRef]
- Conte, A.; Papale, M.; Amalfitano, S.; Mikkonen, A.; Rizzo, C.; De Domenico, E.; Michaud, L.; Lo Giudice, A. Bacterial community structure along the subtidal sandy sediment belt of a high Arctic fjord (Kongsfjorden, Svalbard Islands). Sci. Total Environ. 2018, 619–620, 203–211. [Google Scholar] [CrossRef]
- Granberg, M.; Winberg von Friesen, L.; Bach, L.; Collard, F.; Strand, J.; Gabrielsen, G.W. Anthropogenic Microlitter in Wastewater and Marine Samples from Ny-Ålesund, Barentsburg and Signehamna, Svalbard; Report Number C 373; IVL Swedish Environmental Research Institute: Stockholm, Sweden, 2019; pp. 1–28. ISBN 978-91-7883-020-6. [Google Scholar]
- Hedges, J.I.; Keil, R.G. Sedimentary organic matter preservation: An assessment and speculative synthesis. Mar. Chem. 1995, 49, 81–115. [Google Scholar] [CrossRef]
- Evenset, A.; Leknes, H.; Christensen, G.N.; Warner, N.; Remberger, M.; Gabrielsen, G.W. Screening of New Contaminants in Samples from the Norwegian Arctic: Silver, Platinum, Sucralose, Bisphenol A, Tetrabrombisphenol A, Siloxanes, Phtalates (DEHP), Phosphororganic Flame Retardants; SPFO-Report: 1049/2009; Norwegian Pollution Control Authority: Oslo, Norway, 2009. [Google Scholar]
- Jiao, L.; Zheng, G.J.; Minh, T.B.; Richardson, B.; Chen, L.; Zhang, Y.; Yeung, L.W.; Lam, J.C.W.; Yang, X.; Lam, P.K.S.; et al. Persistent toxic substances in remote lake and coastal sediments from Svalbard, Norwegian Arctic: Levels, sources and fluxes. Environ. Pollut. 2009, 157, 1342–1351. [Google Scholar] [CrossRef]
- Sapota, G.P.; Wojtasik, B.; Burska, D.; Nowiński, K. Persistent organic pollutants (POPs) and polycyclic aromatic hydrocarbons (PAHs) in surface sediments from selected fjords, tidal plains and lakes of the North Spitsbergen. Pol. Polar Res. 2009, 30, 59–76. [Google Scholar]
- Kallenborn, R.; Borgå, K.; Christensen, J.H.; Dowdall, M.; Evenset, A.; Odland, J.Ø.; Ruus, A.; Aspmo Pfaffhuber, K.; Pawlak, J.; Reiersen, L.-O. Combined Effects of Selected Pollutants and Climate Change in the Arctic Environment; Arctic Monitoring and Assessment Programme (AMAP): Oslo, Norway, 2011; pp. 1–108. [Google Scholar]
- Choudhary, S.; Nayak, G.N.; Khare, N. Source, mobility, and bioavailability of metals in fjord sediments of Krossfjord-Kongsfjord system, Arctic, Svalbard. Environ. Sci. Pollut. Res. 2020, 27, 15130–15148. [Google Scholar] [CrossRef]
- Vishnu Sagar, M.K.; Kannan, V.M.; Gopikrishna, V.G.; Krishnan, K.P.; Mohan, M. Geochemistry and distribution of Metals in the Sediments of Kongsfjorden, Svalbard, Arctic. Reg. Stud. Mar. Sci. 2021, 44, 101729. [Google Scholar] [CrossRef]
- Rudnicka-Kępa, P.; Bełdowska, M.; Zaborska, A. Enhanced heavy metal discharges to marine deposits in glacial bays of two Arctic fjords (Hornsund and Kongsfjorden). J. Mar. Syst. 2024, 241, 103915. [Google Scholar] [CrossRef]
- Evenset, A.; Christensen, G.N.; Palerud, R. Environmental Toxins in Marine Sediments in Isfjorden, Svalbard 2009; Research Outside Longyearbyen, Barentsburg, Pyramiden and Coles Bay. Akvaplan-niva report 4707-1, 1-134; Akvaplan-NIVA: Tromsø, Norway, 2009. (In Norwegian) [Google Scholar]
- Van den Heuvel-Greve, M.J.; Szczybelski, A.S.; van den Brink, N.W.; Kotterman, M.J.J.; Kwadijk, C.J.A.F.; Evenset, A.; Murk, A.J. Low organotin contamination of harbour sediment in Svalbard. Polar Biol. 2016, 39, 1699–1709. [Google Scholar] [CrossRef]
- Evenset, A.; Hallanger, I.G.; Tessmann, M.; Warner, N.; Ruus, A.; Borgå, K.; Gabrielsen, G.W.; Christensen, G.; Renaud, P.E. Seasonal variation in accumulation of persistent organic pollutants in an Arctic marine benthic food web. Sci. Tot. Environ. 2016, 542, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Pouch, A.; Zaborska, A.; Pazdro, K. Concentrations and origin of polychlorinated biphenyls (PCBs) and polycyclic aromatic hydrocarbons (PAHs) in sediments of western Spitsbergen fjords (Kongsfjorden, Hornsund, and Adventfjorden). Environ. Monit. Assess. 2017, 189, 175. [Google Scholar] [CrossRef] [PubMed]
- Jæger, I.; Hop, H.; Gabrielsen, G.W. Biomagnification of mercury in selected species from an Arctic marine food web in Svalbard. Sci. Total Environ. 2009, 407, 4744–4751. [Google Scholar] [CrossRef] [PubMed]
- Pouch, A.; Zaborska, A.; Legeżyńska, J.; Deja, K.; Pazdro, K. Assessment of exposure of benthic organisms to selected organochlorine pollutants in the west Spitsbergen fjords. Sci. Total Environ. 2023, 896, 165262. [Google Scholar] [CrossRef]
- Macdonald, R.W.; Harner, T.T.; Fyfe, J. Recent climate change in the Arctic and its impact on contaminants pathway and interpretation on temporal trend data. Sci. Total Environ. 2005, 342, 5–86. [Google Scholar] [CrossRef] [PubMed]
- Ager, D.; Evans, S.; Li, H.; Lilley, A.K.; van der Gast, C.J. Anthropogenic disturbance affects the structure of bacterial communities. Environ. Microbiol. 2010, 12, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Granberg, M.E.; Ask, A.; Gabrielsen, G.G. Local Contamination in Svalbard. Overview and Suggestions for Remediation Actions; Report no. 044; Norsk Polarinstitutt: Tromsø, Norway, 2017. [Google Scholar]
- Zhan, J.; Gao, Y.; Li, W.; Chen, L.; Lin, H.; Lin, Q. Effects of ship emissions on summertime aerosols at Ny–Alesund in the Arctic. Atmos. Pollut. Res. 2014, 5, 500–510. [Google Scholar] [CrossRef]
- Khan, A.L.; Dierssen, H.; Schwarz, J.P.; Schmitt, C.; Chlus, A.; Hermanson, M.; Painter, T.H.; McKnight, D.M. Impacts of coal dust from an active mine on the spectral reflectance of Arctic surface snow in Svalbard, Norway. J. Geophys. Res. Atmos. 2017, 122, 1767–1778. [Google Scholar] [CrossRef]
- Kalinowska, A.; Szopińska, M.; Chmiel, S.; Kończak, M.; Polkowska, Ż.; Artichowicz, W.; Jankowska, K.; Nowak, A.; Łuczkiewicz, A. Heavy Metals in a High Arctic Fiord and Their Introduction with the Wastewater: A Case Study of Adventfjorden-Longyearbyen System, Svalbard. Water 2020, 12, 794. [Google Scholar] [CrossRef]
- Zaborska, A.; Strzelewicz, A.; Rudnicka, P.; Moskalik, M. Processes driving heavy metal distribution in the seawater of an Arctic fjord (Hornsund, southern Spitsbergen). Mar. Pollut. Bull. 2020, 161A, 111719. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Kitae, K.; Deokwon, K.; Moon, H.-B.; Jeon, J. Ny-Ålesund-oriented organic pollutants in sewage effluent and receiving seawater in the Arctic region of Kongsfjorden. Environ. Pollut. 2020, 258, 113792. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, N.E.; Aksu, A.; Karacık, B.; Bayırhan, İ.; Çağlar, N.; Gazioğlu, C.; Özsoy, B. Presence of some commonly used pharmaceutical residues in seawater and net plankton: A case study of Spitsbergen, Svalbard Archipelago. Int. J. Environ. Geoinform. 2022, 9, 1–10. [Google Scholar] [CrossRef]
- Sokołowski, A.; Mordec, M.; Caban, M.; Øverjordet, I.B.; Wielogórska, E.; Włodarska-Kowalczuk, M.; Balazy, P.; Chełchowski, M.; Lepoint, G. Bioaccumulation of pharmaceuticals and stimulants in macrobenthic food web in the European Arctic as determined using stable isotope approach. Sci. Total Environ. 2024, 909, 168557. [Google Scholar] [CrossRef]
- Jokanović, S.; Kajan, K.; Perović, S.; Ivanić, M.; Mačić, V.; Orlić, S. Anthropogenic influence on the environmental health along Montenegro coast based on the bacterial and chemical characterization. Environ. Pollut. 2021, 271, 116383. [Google Scholar] [CrossRef] [PubMed]
- Nogales, B.; Lanfranconi, M.P.; Piña-Villalonga, J.M.; Bosch, R. Anthropogenic perturbations in marine microbial communities. FEMS Microbiol. Rev. 2011, 35, 275–298. [Google Scholar] [CrossRef] [PubMed]
- Gillan, D.C.; Danis, B.; Pernet, P.; Joly, G.; Dubois, P. Structure of sediment-associated microbial communities along a heavy-metal contamination gradient in the marine environment. Appl. Environ. Microbiol. 2005, 71, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Varma, A. Role of Enzymes in Maintaining Soil Health. In Soil Enzymology. Soil Biology; Shukla, G., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 22. [Google Scholar] [CrossRef]
- Alkorta, I.; Aizpurua, A.; Riga, P.; Albizu, I.; Amézaga, I.; Garbisu, C. Soil Enzyme Activities as Biological Indicators of Soil Health. Rev. Environ. Health 2003, 18, 65–73. [Google Scholar] [CrossRef]
- Rao, M.A.; Scelza, R.; Acevedo, F.; Diez, M.C.; Gianfreda, L. Enzymes as useful tools for environmental purposes. Chemosphere 2014, 107, 145–162. [Google Scholar] [CrossRef]
- Maher, W.; Batley, G.E.; Lawrence, I. Assessing the health of sediment ecosystems: Use of chemical measurements. Freshw. Biol. 1999, 41, 361–372. [Google Scholar] [CrossRef]
- Astudillo-García, C.; Hermans, S.M.; Stevenson, B.; Buckley, H.L.; Lear, G. Microbial assemblages and bioindicators as proxies for ecosystem health status: Potential and limitations. Appl. Microbiol. Biotechnol. 2019, 103, 6407–6421. [Google Scholar] [CrossRef]
- Caruso, G.; Genovese, L.; Mancuso, M.; Modica, A. Effects of fish farming on microbial enzyme activities and densities: Comparison between three Mediterranean sites. Lett. Appl. Microbiol. 2003, 37, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Chen, Z.; Deng, Y.; Zhao, D.; Gao, P.; Zhang, L.; Tu, Q.; Qu, L.; Zheng, L.; Zhang, Y.; et al. Contrasting archaeal and bacterial community assembly processes and the importance of rare taxa along a depth gradient in shallow coastal sediments. Sci. Total Environ. 2022, 852, 158411. [Google Scholar] [CrossRef]
- Caruso, G.; Maimone, G.; Rappazzo, A.C.; Dell’Acqua, O.; Laganà, P.; Azzaro, M. Microbial Biofilm Colonizing Plastic Substrates in the Ross Sea (Antarctica): First Overview of Community-Level Physiological Profiles. J. Mar. Sci. Eng. 2023, 11, 1317. [Google Scholar] [CrossRef]
- Rutgers, M.; Wouterse, M.; Drost, S.M.; Breure, A.M.; Mulder, C.; Stone, D.; Creamer, R.E.; Winding, A.; Bloem, J. Monitoring soil bacteria with community-level physiological profiles using Biolog™ ECO-plates in the Netherlands and Europe. Appl. Soil Ecol. 2016, 97, 23–35. [Google Scholar] [CrossRef]
- Wan, L.; Caruso, G.; Cao, X.; Song, C.; Maimone, G.; Rappazzo, A.C.; Laganà, P.; Zhou, Y. Microbial Response to Coastal-Offshore Gradients in Taiwan Straits: Community Metabolism and Total Prokaryotic Abundance as Potential Proxies. Microb. Ecol. 2023, 85, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Handler, E.R.; Andersen, S.D.J.; Gradinger, R.; McGovern, M.; Vader, A.; Poste, A.E. Seasonality in land-ocean connectivity and local processes control sediment bacterial community structure and function in a High Arctic tidal flat. FEMS Microb. Ecol. 2024, 100, fiad162. [Google Scholar] [CrossRef]
- Zaborska, A.; Włodarska-Kowalczuk, M.; Legeżyńska, J.; Jankowska, E.; Winogradow, A.; Deja, K. Sedimentary organic matter sources, benthic consumption and burial in west Spitsbergen fjords—Signs of maturing of Arctic fjordic systems? J. Mar. Syst. 2018, 180, 112–123. [Google Scholar] [CrossRef]
- Piquet, A.M.-T.; Scheepens, J.F.; Bolhuis, H.; Wiencke, C.; Buma, A.G.J. Variability of protistan and bacterial communities in two Arctic fjords (Spitsbergen). Polar Biol. 2010, 33, 1521–1536. [Google Scholar] [CrossRef]
- Skogseth, R.; Olivier, L.L.A.; Nilsen, F.; Falck, E.; Fraser, N.; Tverberg, V.; Ledang, A.B.; Vader, A.; Jonassen, M.O.; Søreide, J.; et al. Variability and decadal trends in the Isfjorden (Svalbard) ocean climate and circulation—An indicator for climate change in the European Arctic. Progr. Oceanogr. 2020, 187, 102394. [Google Scholar] [CrossRef]
- Prominska, A.; Cisek, M.; Walczowski, W. Kongsfjorden and Hornsund hydrography—Comparative study based on a multiyear survey in fjords of West Spitsbergen. Oceanologia 2017, 59, 397–412. [Google Scholar] [CrossRef]
- Norwegian Polar Institute. TOPOSVALBARD. Available online: https://toposvalbard.npolar.no/ (accessed on 3 June 2024).
- Hoppe, H.G. Use of fluorogenic model substrates for extracellular enzyme activity (EEA) measurement of bacteria. In Handbook of Methods in Aquatic Microbial Ecology; Kemp, P.F., Sherr, B.F., Sherr, E.B., Cole, J.J., Eds.; Lewis Publisher: London, UK, 1993; pp. 423–431. [Google Scholar]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [PubMed]
- Sala, M.M.; Arin, L.; Balagué, V.; Felipe, J.; Guadayol, O.; Vaqué, D. Functional diversity of bacterioplankton assemblages in Western Antarctic seawaters during late spring. Mar. Ecol. Progr. Ser. 2005, 292, 13–21. [Google Scholar] [CrossRef]
- Sala, M.M.; Estrada, M.; Gasol, J.M. Seasonal changes in the functional diversity of bacterioplankton in contrasting coastal environments of the NW Mediterranean. Aquat. Microb. Ecol. 2006, 44, 1–9. [Google Scholar] [CrossRef]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Bergey’s Manual of Determinative Bacteriology, 9th ed.; Williams & Wilkins: Baltimore, MD, USA, 1994; pp. 1–787. [Google Scholar]
- Laganà, P.; Votano, L.; Caruso, G.; Azzaro, M.; Lo Giudice, A.; Delia, S. Bacterial isolates from the Arctic region (Pasvik River, Norway): Assessment of biofilm production and antibiotic susceptibility profiles. Environ. Sci. Pollut. Res. 2018, 25, 1089–1102. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turk, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- EUCAST (European Committee on Antimicrobial Susceptibility Testing). Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12.0. 2022. Available online: http://www.eucast.org (accessed on 3 June 2024).
- Petro, C.; Starnawski, P.; Schramm, A.; Kjeldsen, K.U. Microbial community assembly in marine sediments. Aquat. Microb. Ecol. 2017, 79, 177–195. [Google Scholar] [CrossRef]
- Hamdan, L.J.; Coffin, R.B.; Sikaroodi, M.; Greinert, J.; Treude, T.; Gillevet, P.M. Ocean currents shape the microbiome of Arctic marine sediments. ISME J. 2013, 7, 685–696. [Google Scholar] [CrossRef]
- Delpech, L.M.; Vonnahme, T.R.; McGovern, M.; Gradinger, R.; Præbel, K.; Poste, A.E. Terrestrial inputs shape coastal bacterial and archaea communities in a high Arctic fjord (Isfjorden, Svalbard). Front. Microbiol. 2021, 12, 614634. [Google Scholar] [CrossRef]
- Jørgensen, B.B.; Laufer, K.; Michaud, A.B.; Wehrmann, L.M. Biogeochemistry and microbiology of high Arctic marine sediment ecosystems—Case study of Svalbard fjords. Limnol. Oceanogr. 2020, 66, S273–S292. [Google Scholar] [CrossRef]
- Kim, H.; Kwon, S.Y.; Lee, K.; Lim, D.; Han, S.; Kim, T.W.; Joo, Y.J.; Lim, J.; Kang, M.H.; Nam, S.I. Input of terrestrial organic matter linked to deglaciation increased mercury transport to the Svalbard fjords. Sci. Rep. 2020, 10, 3446. [Google Scholar] [CrossRef]
- Kim, J.-H. Large ancient organic matter contributions to Arctic marine sediments (Svalbard). Limnol. Oceanogr. 2011, 56, 1463–1474. [Google Scholar] [CrossRef]
- Thomas, F.A.; Mohan, M.; Krishnan, K.P. Bacterial diversity and their metabolic profiles in the sedimentary environments of Ny-Ålesund, Arctic. Antonie van Leeuwenhoek 2021, 114, 1339–1360. [Google Scholar] [CrossRef]
- Vishnupriya, S.; Jabir, T.; Krishnan, K.P.; Mohamed Hatha, A.A. Bacterial community structure and functional profiling of high Arctic fjord sediments. World J. Microbiol. Biotechnol. 2021, 37, 133. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.N.; Winding, A. Microorganisms as Indicators of Soil Health; Technical Report No. 388; National Environmental Research Institute: Roskilde, Denmark, 2002; pp. 1–85. [Google Scholar]
- Fierer, N.; Wood, S.A.; Bueno de Mesquita, C.P. How microbes can, and cannot, be used to assess soil health. Soil Biol. Biochem. 2021, 153, 108111. [Google Scholar] [CrossRef]
- Fang, X.M.; Zhang, T.; Li, J.; Wang, N.-F.; Wang, Z.; Yu, L.-Y. Bacterial community pattern along the sediment seafloor of the Arctic fjorden (Kongsfjorden, Svalbard). Antonie van Leeuwenhoek 2019, 112, 1121–1136. [Google Scholar] [CrossRef]
- Venkatachalam, S.; Gopinath, A.; Krishnan, K.P. Fjords of the western and northern regions of Svalbard harbour distinct bacterioplankton community structures. World J. Microbiol. Biotechnol. 2023, 39, 57. [Google Scholar] [CrossRef]
- Kalinowska, A.; Ameryk, A.; Jankowska, K. Microbiological survey in two Arctic fjords: Total bacterial number and biomass comparison of Hornsund and Kongsfjorden. In Impact of Climate Changes on Marine Environments, GeoPlanet: Earth and Planetary Sciences; Zielinski, T., Weslawski, M., Kuliński, K., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 115–126. [Google Scholar] [CrossRef]
- Børsheim, K.Y.; Drinkwater, K.F. Different temperature adaptation in Arctic and Atlantic heterotrophic bacteria in the Barents Sea Polar Front region. J. Mar. Syst. 2014, 130, 160–166. [Google Scholar] [CrossRef]
- Beszczynska-Moller, A.; Weslawski, J.M.; Walczowski, W.; Zajaczkowski, M. Estimation of glacial meltwater discharge into Svalbard coastal waters. Oceanologia 1997, 39, 289–299. [Google Scholar]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The microbial engines that drive Earth’s biogeochemical cycles. Science 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.S.; Elliott, M. Ecology of Marine Sediments: From Science to Management; Oxford University Press: Oxford, UK, 2009; pp. 1–216. [Google Scholar] [CrossRef]
- Brakstad, O.G.; Nonstad, I.; Faksness, L.G.; Brandvik, P.J. Responses of Microbial Communities in Arctic Sea Ice After Contamination by Crude Petroleum Oil. Microb. Ecol. 2008, 55, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, M.T.; Kirchman, D.L. Natural assemblages of marine proteobacteria and members of the Cytophaga-Flavobacter cluster consuming low- and high-molecular-weight dissolved organic matter. Appl. Environ. Microbiol. 2000, 66, 1692–1697. [Google Scholar] [CrossRef] [PubMed]
- Teske, A.; Durbin, A.; Ziervogel, K.; Cox, C.; Arnosti, C. Microbial community composition and function in permanently cold seawater and sediments from an Arctic fjord of Svalbard. Appl. Environ. Microbiol. 2011, 77, 2008–2018. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, K.; Hassenrück, C.; Salman-Carvalho, V.; Holtappels, M.; Bienhold, C. Response of bacterial communities to different detritus compositions in arctic deep-sea sediments. Front. Microbiol. 2017, 8, 266. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, S.; Kerhervé, P.; Calleja, M.L.; Many, G.; Morata, N. Glacier inputs influence organic matter composition and prokaryotic distribution in a high Arctic fjord (Kongsfjorden, Svalbard). J. Mar. Syst. 2016, 164, 112–127. [Google Scholar] [CrossRef]
- Hargrave, B.T.; Holmer, M.; Newcombe, C.P. Towards a classification of organic enrichment in marine sediments based on biogeochemical indicators. Mar. Pollut. Bull. 2008, 56, 810–824. [Google Scholar] [CrossRef]
- Fenice, M.; Gallo, A.; Juarez-Jimenez, B.; Gonzalez-Lopez, J. Screening for extracellular enzyme activities by bacteria isolated from samples collected in the Tyrrhenian Sea. Ann. Microbiol. 2007, 57, 93–99. [Google Scholar] [CrossRef]
- Davey, K.E.; Kirby, R.R.; Turley, C.M.; Weightman, A.J.; Fry, J.C. Depth variation of bacterial extracellular enzyme activity and population diversity in the northeastern North Atlantic Ocean. Deep Sea Res. Part II 2001, 48, 1003–1017. [Google Scholar] [CrossRef]
- Sjölund, M.; Bonnedahl, J.; Hernandez, J.; Bengtsson, S.; Cederbrant, G.; Pinhassi, J.; Kahlmeter, G.; Olsen, B. Dissemination of multidrug-resistant bacteria into the Arctic. Emerg. Infect. Dis. 2008, 14, 70–72. [Google Scholar] [CrossRef]
- Kalinowska, A.; Jankowska, K.; Fudala-Ksiazek, S.; Pierpaoli, M.; Luczkiewicz, A. The microbial community, its biochemical potential, and the antimicrobial resistance of Enterococcus spp. in Arctic lakes under natural and anthropogenic impact (West Spitsbergen). Sci. Total Environ. 2021, 763, 142998. [Google Scholar] [CrossRef] [PubMed]
- Visnupriya, S.; Jabir, T.; Akhil Prakash, E.; Mohamed Hatha, A.A. Antibiotic resistance of heterotrophic bacteria from the sediments of adjoining high Arctic fjords, Svalbard. Braz. J. Microbiol. 2024. [Google Scholar] [CrossRef]
- Allen, H.; Donato, J.; Wang, H.H.; Cloud-Hansen, K.A.; Davies, J.; Handelsman, J. Call of the wild: Antibiotic resistance genes in natural environments. Nat. Rev. Microbiol. 2010, 8, 251–259. [Google Scholar] [CrossRef]
- Tan, L.; Li, L.; Ashbolt, N.; Wang, X.; Cui, Y.; Zhu, X.; Xu, Y.; Yang, Y.; Mao, D.; Luo, Y. Arctic antibiotic resistance gene contamination, a result of anthropogenic activities and natural origin. Sci. Tot. Environ. 2018, 621, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.C.; Lee, N.; Aw, T.G. Antibiotic resistance in minimally human-impacted environments. Int. J. Environ. Res. Public Health 2020, 17, 3939. [Google Scholar] [CrossRef] [PubMed]
- Mohamed Hatha, A.A.; Neethu, S.; Nikhil, S.M.; Mujeeb Rahiman, K.M.; Krishnan, K.P.; Saramma, A.V. Relatively high antibiotic resistance among heterotrophic bacteria from Arctic fjord sediments than water—Evidence towards better selection pressure in the fjord sediments. Polar Sci. 2015, 9, 382–388. [Google Scholar] [CrossRef]
- AMAP (Arctic Monitoring and Assessment Programme). AMAP Assessment 2018: Biological Effects of Contaminants on Arctic Wildlife and Fish; Arctic Monitoring and Assessment Programme (AMAP): Tromsø, Norway, 2018; pp. vii+84. [Google Scholar]
- Rauseo, J.; Spataro, F.; Pescatore, T.; Patrolecco, L. Multiresidue determination and predicted risk assessment of emerging contaminants in sediments from Kongsfjorden, Svalbard. Sci. Tot. Environ. 2024, 922, 171156. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, G.; Song, W.; Ye, C.; Lin, H.; Li, Z.; Liu, W. Plastics in the marine environment are réservoirs for antibiotic and metal resistance genes. Environ. Int. 2019, 123, 79–86. [Google Scholar] [CrossRef]
- Agnew, A.; Wang, J.; Fanning, S.; Bearhop, S.; McMahon, B.J. Insights into antimicrobial resistance among long distance migratory East Canadian High Arctic light-bellied Brent geese (Branta bernicla hrota). Ir. Vet. 2015, 69, 13. [Google Scholar] [CrossRef]
- Pouch, A.; Zaborska, A. Climate change influence on migration of contaminants in the Arctic marine environment. GeoPlanet Earth Planet. Sci. 2015, 22, 75–90. [Google Scholar] [CrossRef]
- Grenni, P.; Ancona, V.; Barra Caracciolo, A. Ecological effects of antibiotics on natural ecosystems: A review. Microchem. J. 2018, 136, 25–39. [Google Scholar] [CrossRef]
- Zaborska, A.; Beszczyńska-Moller, A.; Włodarska-Kowalczuk, M. History of heavy metal accumulation in the Svalbard area: Distribution, origin and transport pathways. Environ. Poll. 2017, 231, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Evenset, A.; Christensen, G.N.; Carroll, J.; Zaborska, A.; Berger, U.; Herzke, D.; Gregor, D. Historical trends in persistent organic pollutants and metals recorded in sediment from Lake Ellasjøen, Bjørnøya, Norwegian Arctic. Environ. Pollut. 2007, 146, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Li, L.-G.; Xia, Y.; Zhang, T. Co-occurrence of antibiotic and metal resistance genes revealed in complete genome collection. ISME J. 2017, 11, 651–662. [Google Scholar] [CrossRef]
- Gianfreda, L.; Rao, M.A. Interactions Between Xenobiotics and Microbial and Enzymatic Soil Activity. Crit. Rev. Environ. Sci. Technol. 2008, 38, 269–310. [Google Scholar] [CrossRef]
- Makowska-Zawierucha, N.; Mokracka, J.; Małecka, M.; Balazy, P.; Chełchowski, M.; Ignatiuk, D.; Zawierucha, K. Quantification of class 1 integrons and characterization of the associated gene cassettes in the high Arctic—Interplay of humans and glaciers in shaping the aquatic resistome. Ecol. Indic. 2022, 145, 109633. [Google Scholar] [CrossRef]
- Astrup, E. Antibiotic Resistance in Norway- NIPH. Public Health Report. 2014. Available online: https://www.fhi.no/en/op/public-health-report-2014/health--disease/antibiotic-resistance-in-norway---p/ (accessed on 14 September 2023).
- Norwegian Ministries. National Strategy against Antibiotic Resistance 2015–2020; Norwegian Ministry of Health and Care Services: Oslo, Norway, 2015; pp. 1–36. [Google Scholar]
- NORM/NORM-VET 2021. Usage of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Norway. Tromsø, Norway. September 2022, pp. 1–64. Available online: www.vetinst.no (accessed on 14 September 2023).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Area | Latitude | Longitude | Depth (m) | Sediment Type | Sediment Color |
|---|---|---|---|---|---|---|
| KR1 | Krossfjorden inner | 79.3282 | 11.6341 | 174 | Clay | Gray |
| KR2 | Krossfjorden central | 79.2015 | 11.7395 | 282 | Clay | Gray |
| KR3 | Krossfjorden outer | 79.1261 | 11.8177 | 60 | Clay | Gray |
| KO1 | Kongsfjorden inner | 78.8933 | 12.4722 | 74 | Mud | Brown |
| KO2 | Kongsfjorden central | 78.965 | 11.8994 | 347 | Clay | Gray |
| KO3 | Kongsfjorden outer | 79.0112 | 11.4058 | 369 | Mud | Black |
| IS1 | Isfjorden central | 78.2657 | 15.1257 | 248 | Mud | Black |
| IS2 | Isfjorden central | 78.395 | 15.5663 | 202 | Mud | Black |
| IS3 | Isfjorden inner | 78.4479 | 16.078 | 94 | Mud | Black |
| D1 | Dicksonfjorden inner | 78.8097 | 15.3842 | 16 | Mud | Brown |
| N1 | Nordfjorden central | 78.5152 | 14.9392 | 144 | Mud | Black |
| B1 | Billefjorden inner | 78.6090 | 16.4805 | 141 | Clay | Gray |
| S1 | Sassenfjorden central | 78.4003 | 16.3756 | 31 | Mud | Black |
| A1 | Adventfjorden central | 78.2372 | 15.6360 | 58 | Mud | Black |
| G1 | Grønfjorden outer | 78.0873 | 14.1221 | 122 | Mud | Black |
| H1 | Hornsund inner | 76.9664 | 15.7297 | 242 | Mud | Black |
| H2 | Hornsund central | 77.0019 | 16.0846 | 84 | Mud | Black |
| H3 | Hornsund outer | 77.0021 | 16.4646 | 125 | Mud | Black |
| Richness | Shannon–Weaver | |
|---|---|---|
| Northern area | ||
| KR2 | 28 | 3.054 |
| KR3 | 28 | 3.171 |
| KO1 | 19 | 2.875 |
| KO2 | 24 | 3.012 |
| KO3 | 29 | 3.162 |
| Central area | ||
| IS1 | 26 | 3.001 |
| IS2 | 21 | 2.935 |
| IS3 | 29 | 3.191 |
| D1 | 23 | 2.965 |
| N1 | 25 | 3.108 |
| B1 | 28 | 3.127 |
| S1 | 31 | 3.251 |
| A1 | 28 | 3.180 |
| G1 | 31 | 3.251 |
| Southern area | ||
| H1 | 26 | 3.048 |
| H2 | 31 | 3.177 |
| H3 | 27 | 2.999 |
| GLOBAL | LAP | GLU | AP | MA | TSA | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Factors | df | PsF (P) | PsF (P) | PsF (P) | PsF (P) | PsF (P) | ||||
| Areas | 2 | 0.01 (0.98) | 0.074 (0.09) | 1.74 (0.13) | 0.84 (0.35) | 0.42 (0.58) | ||||
| Stations | 17 | |||||||||
| GLOBAL | AWCD | Richness | Shannon–Weaver | Complex carbon sources | Carbohydrates | Phosphate–carbon sources | Carboxylic acids | Amino acids | Amines | |
| Factors | df | PsF (P) | PsF (P) | PsF (P) | PsF (P) | PsF (P) | PsF (P) | PsF (P) | PsF (P) | PsF (P) |
| Areas | 2 | 0.44 (0.57) | 0.25 (0.72) | 0.33 (0.65) | 0.09 (0.88) | 60.14 (8 × 10−11 ***) | 1.46 (0.18) | 0.95 (0.31) | 0.71 (0.41) | 0.07 (0.90) |
| Stations | 17 | |||||||||
| Carbohydrates | D-cellobiose | A-D-lactose | B-methyl-D-glucoside | D-xylose | i-erythritol | D-mannitol | N-acetyl-D-glucosamin | |||
| Factors | df | PsF (P) | PsF (P) | PsF (P) | PsF (P) | PsF (P) | PsF (P) | PsF (P) | ||
| Areas | 2 | 0.13 (0.83) | 2.12 (0.15) | 0.17 (0.80) | 0.04 (0.95) | 1.93 (0.18) | 1.18 (0.24) | 0.06 (0.91) | ||
| Stations | 17 | |||||||||
| PAIR- WISE | D-cellobiose | A-D-lactose | B-methyl-D-glucoside | D-xylose | I-erythritol | D-mannitol | N-acetyl-D-glucosamin | |||
| Areas | 4.91 (0.01 **) | 3.98 (0.03 **) | 4.60 (0.02 **) | 1.02 (0.37) | 2.59 (0.09) | 3.14 (0.06) | 5.13 (0.01 **) |
| D | Variable (% Contribution) | ||
|---|---|---|---|
| KR1 vs. | KR2 | 0.08 | Shannon-Weaver (32.55%) |
| KR3 | 92.22 | Glycyl-L-glutamic acid (10.41%) | |
| KO3 | 80.43 | AP (12.55%) | |
| KO3 | 51.96 | AP (17.86%) | |
| KO1 | 147.57 | Carboxylic acids (10.03%) | |
| KR2 vs. | KR3 | 91.91 | Glycyl-L-glutamic acid (10.45%) |
| KO3 | 80.84 | D-galactonic acid y-lactone (12.22%) | |
| KO2 | 50.87 | AP (17.64%) | |
| KO1 | 145.05 | Putrescine (10.03%) | |
| KR3 vs. | KO3 | 49.87 | A-ketobutyric acid (13.99%) |
| KO2 | 78.33 | L-arginine (11.82%) | |
| KO1 | 143.27 | Putrescine (8.02%) | |
| KO2 vs. | KO1 | 121.19 | Amines (17.53%) |
| KO3 vs. | KO2 | 56.92 | I-erythritol (15.78%) |
| KO1 | 164.41 | Putrescine (10.43%) | |
| IS1 vs. | A1 | 87.24 | N-acetyl-D-glucosamin (14.34%) |
| G1 | 87.62 | N-acetyl-D-glucosamin (12.52%) | |
| IS2 vs. | N1 | 94.28 | Complex C sources (13.69%) |
| IS1 | 110.18 | Complex C sources (8.96%) | |
| A1 | 152.84 | Tween 40 (13.16%) | |
| G1 | 158.65 | Complex C sources (11.69%) | |
| IS3 vs. | S1 | 44.71 | LAP (14.81%) |
| N1 | 60.15 | LAP (19.94%) | |
| IS1 | 101.57 | LAP (11.85%) | |
| IS2 | 139.92 | Complex C sources (10.55%) | |
| A1 | 62.01 | LAP (18.74%) | |
| G1 | 49.95 | LAP (19.76%) | |
| A1 vs. | G1 | 41.59 | Itaconic acid(8.29%) |
| B1 vs. | IS1 | 105.99 | GLU (14.79%) |
| IS2 | 153.11 | GLU (10.69%) | |
| IS3 | 53.68 | Itaconic acid (18.76%) | |
| S1 | 67.13 | GLU (16.88%) | |
| N1 | 72.67 | GLU (22.26%) | |
| A1 | 66.16 | GLU (23.88%) | |
| G1 | 70.16 | GLU (20.48%) | |
| D1 vs. | IS1 | 111.98 | Amino acids (11.67%) |
| IS2 | 192.02 | D-mannitol (9.33%) | |
| IS3 | 142.49 | L-threonine (9.51%) | |
| B1 | 119.01 | GLU (12.26%) | |
| S1 | 116.14 | L-threonine (12.15%) | |
| N1 | 102.85 | Amino acids (11.41%) | |
| A1 | 94.46 | N-acetyl-D-glucosamin (8.83%) | |
| G1 | 115.04 | L-threonine (12.26%) | |
| N1 vs. | IS1 | 58.32 | N-acetyl-D-glucosamin (18.64%) |
| A1 | 61.30 | Tween 40 (12.65%) | |
| G1 | 45.88 | Phenylethyl-amine (12.13%) | |
| S1 vs. | IS1 | 88.62 | N-acetyl-D-glucosamin (12.38%) |
| IS2 | 157.13 | Complex C sources (11.81%) | |
| N1 | 46.31 | Phenylethyl-amine (12.02%) | |
| A1 | 644.73 | TSA (9.41%) | |
| G1 | 1.92 | MA (71.09%) | |
| H1 vs. | H2 | 105.09 | MA (10.24%) |
| H3 | 83.55 | MA (20.25%) | |
| H2 vs. | H3 | 77.07 | L-phenylalanine (14.43%) |
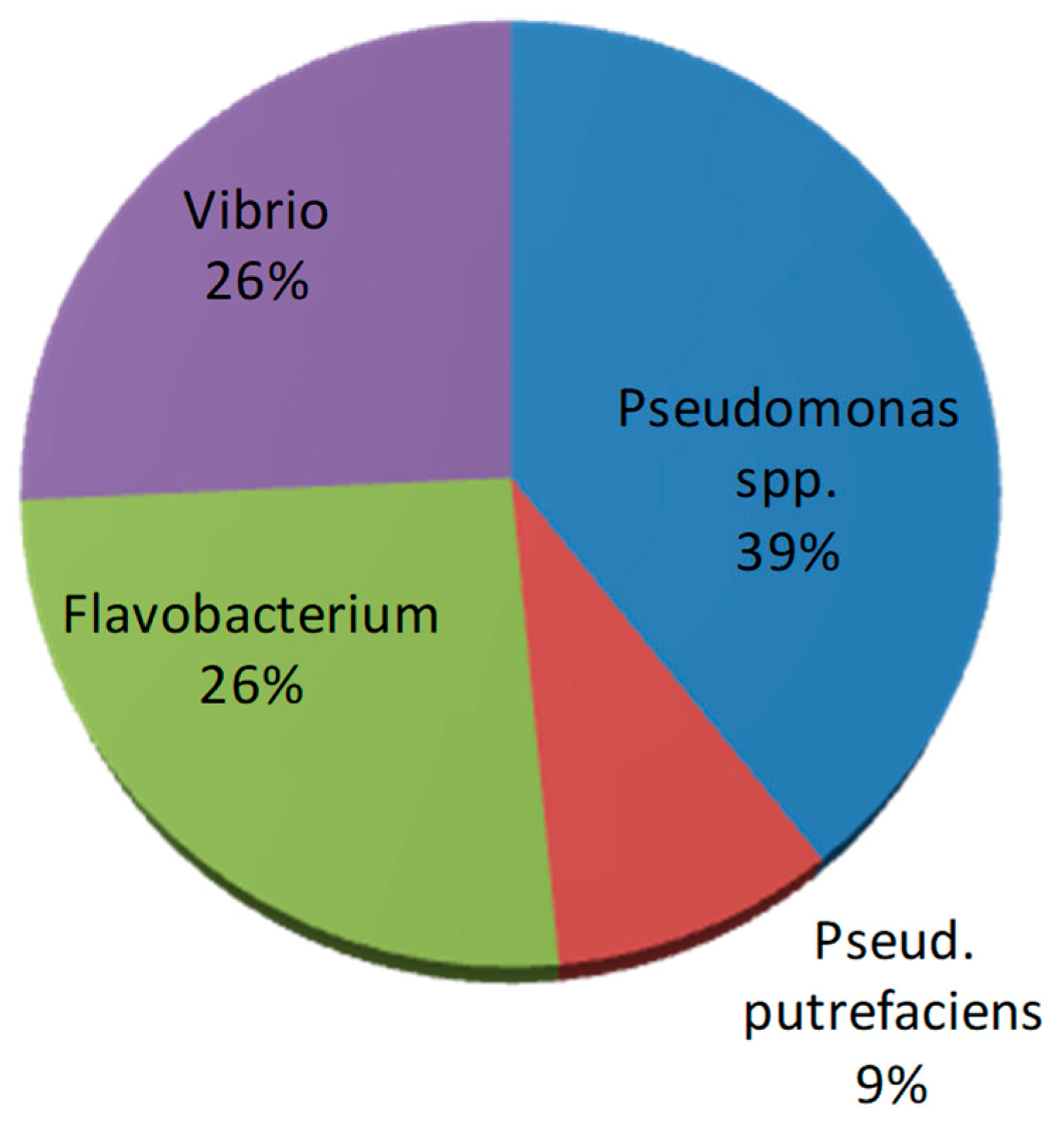
| Pseudomonas spp. | Pseud. putrefaciens | Flavobacterium spp. | Vibrio spp. | Total Isolates | ||
|---|---|---|---|---|---|---|
| Northern area | Krossfjorden | 4 | 5 | 9 | ||
| Kongsfjorden | 3 | 5 | 1 | 3 | 12 | |
| Central area | Isfjorden | 17 | 4 | 13 | 34 | |
| Southern area | Hornsund | 2 | 1 | 7 | 1 | 11 |
| Total isolated strains | 26 | 6 | 17 | 17 | 66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caruso, G.; Rappazzo, A.C.; Maimone, G.; Zappalà, G.; Cosenza, A.; Szubska, M.; Zaborska, A. Svalbard Fjord Sediments as a Hotspot of Functional Diversity and a Reservoir of Antibiotic Resistance. Environments 2024, 11, 148. https://doi.org/10.3390/environments11070148
Caruso G, Rappazzo AC, Maimone G, Zappalà G, Cosenza A, Szubska M, Zaborska A. Svalbard Fjord Sediments as a Hotspot of Functional Diversity and a Reservoir of Antibiotic Resistance. Environments. 2024; 11(7):148. https://doi.org/10.3390/environments11070148
Chicago/Turabian StyleCaruso, Gabriella, Alessandro Ciro Rappazzo, Giovanna Maimone, Giuseppe Zappalà, Alessandro Cosenza, Marta Szubska, and Agata Zaborska. 2024. "Svalbard Fjord Sediments as a Hotspot of Functional Diversity and a Reservoir of Antibiotic Resistance" Environments 11, no. 7: 148. https://doi.org/10.3390/environments11070148