
Ecological Shifts: Plant Establishment in an Animal-Based Ecosystem



Abstract
:1. Introduction
2. Materials and Methods
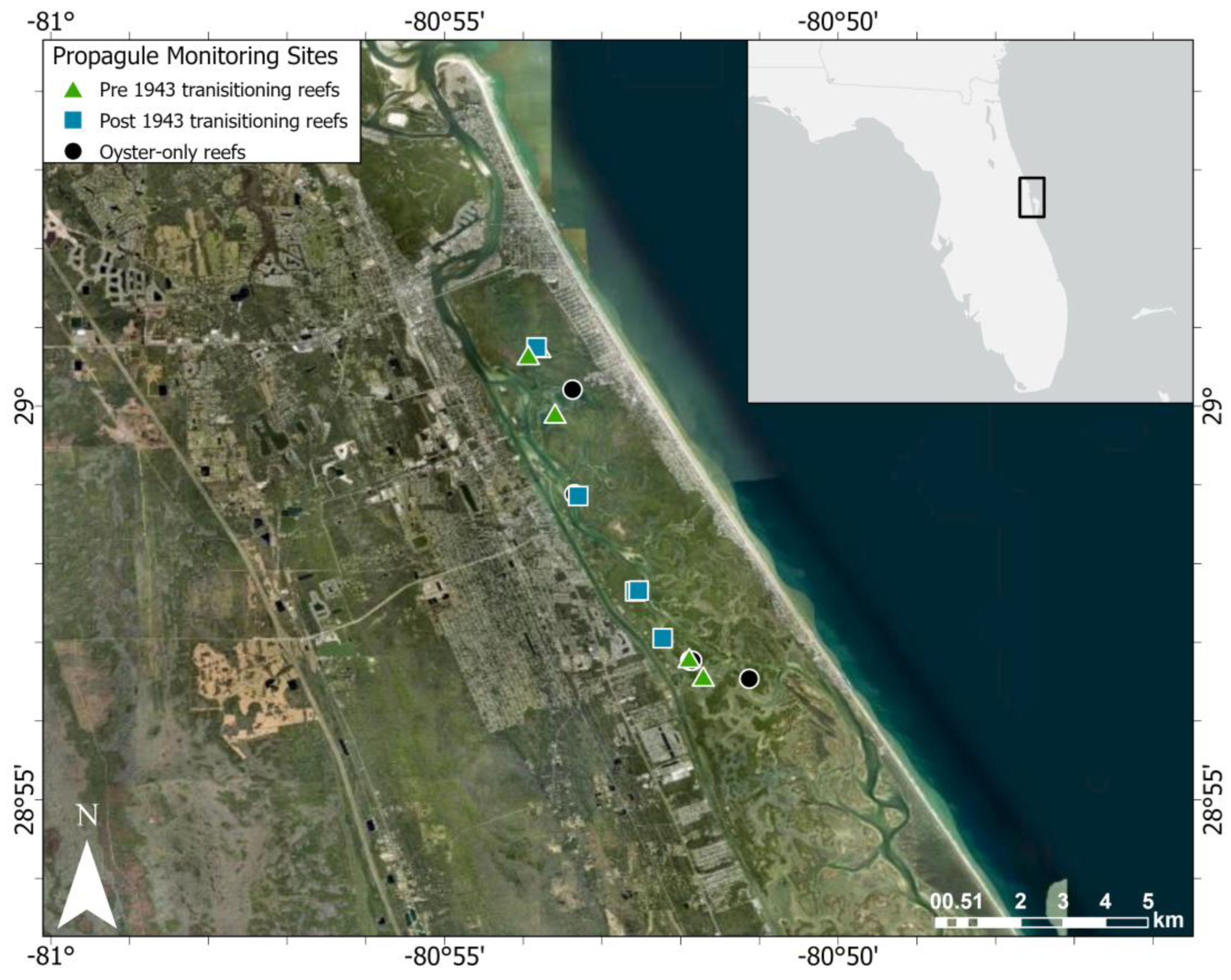
2.1. Study Site
2.2. Reefs of the Eastern Oyster Crassostrea virginica in Mosquito Lagoon
2.3. Mosquito Lagoon Mangroves
2.4. Tracking Propagule Recruitment
2.5. Oyster Reef Metrics
2.6. Temperature, Water Level, and Salinity Data
2.7. Statistical Analyses for Mangrove Recruitment and Success
3. Results
3.1. Temperature Data and Water Levels
3.2. Mangrove versus Oyster Areas on Oyster Reefs
3.3. Oyster Densities on Reefs
3.4. Natural Recruitment of Propagules
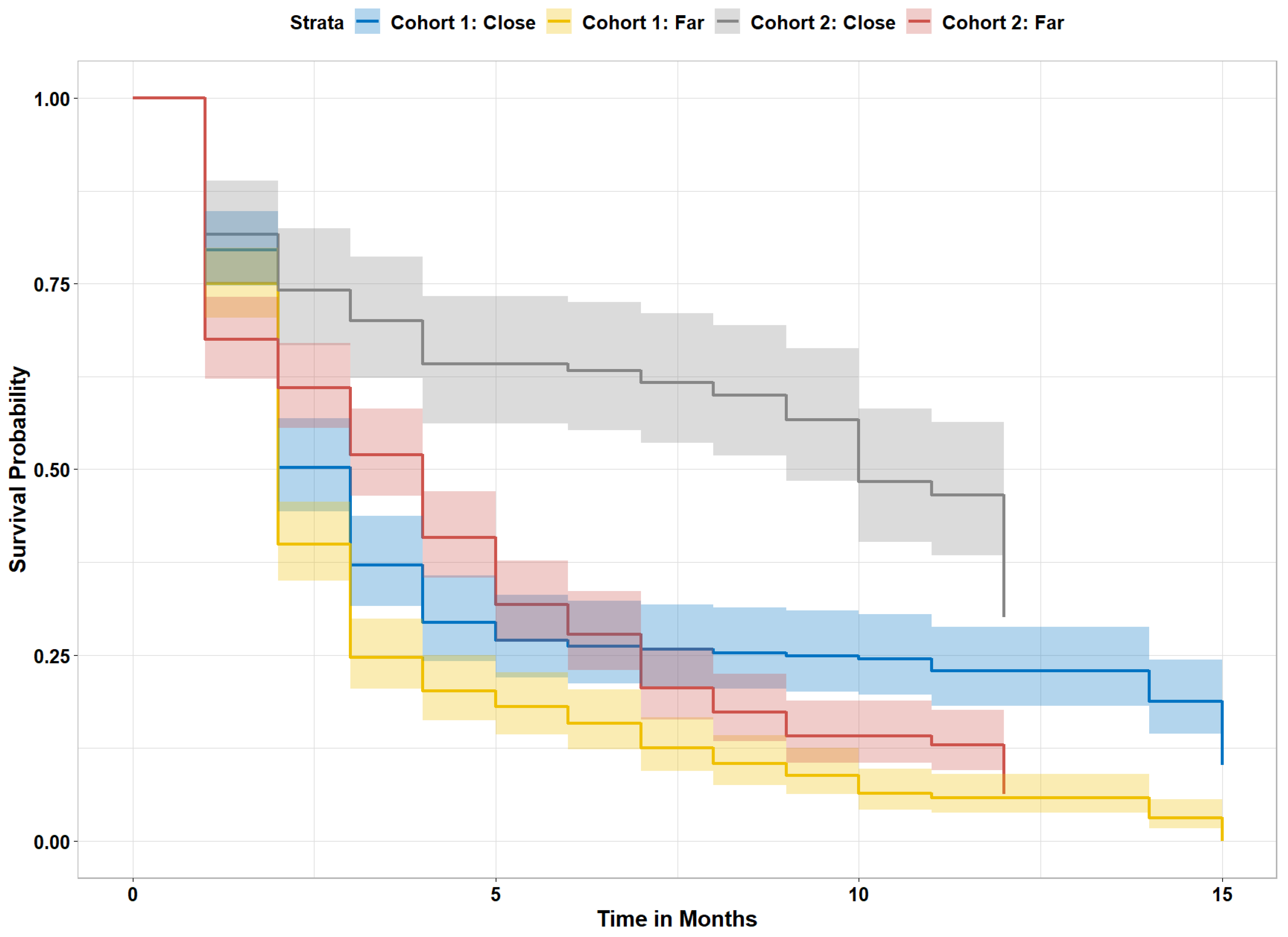
3.5. Propagule Arrival and Survival Densities on Reefs
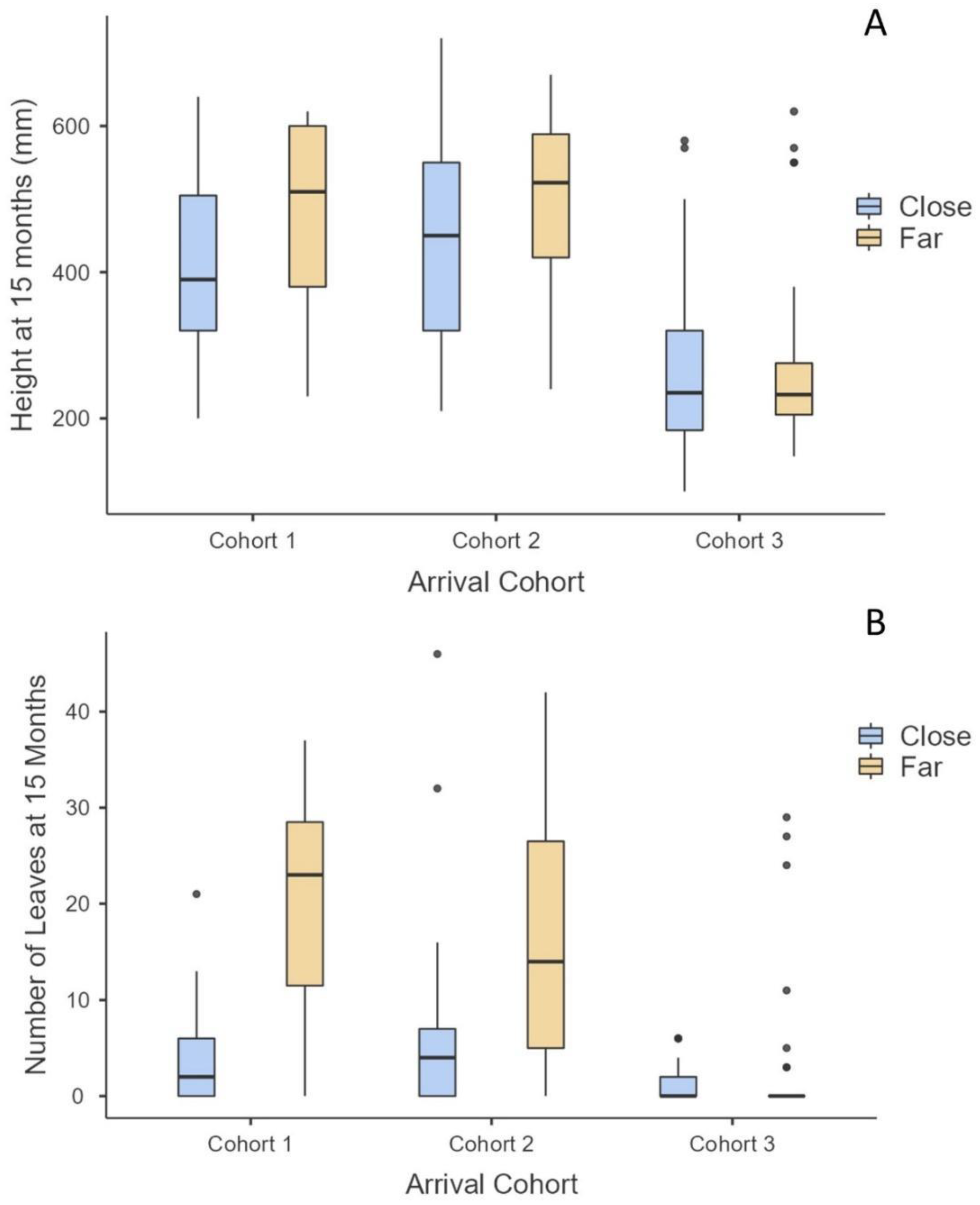
3.6. Adult Mangrove Nursery Impacts
4. Discussion
4.1. Success: Comparisons among Mangrove Species
4.2. Positive Feedback Loops
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sousa, W. The role of disturbance in natural communities. Ann. Rev. Ecol. Syst. 1984, 15, 353–391. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, E.; Foley, J.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, R.C. Mass mortality of Diadema antillarum I. Long term effects on sea urchin population dynamics and coral reef algal communities. Mar. Biol. 1990, 104, 67–77. [Google Scholar] [CrossRef]
- Hughes, T.P. Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science 1994, 265, 1547–1551. [Google Scholar] [CrossRef] [PubMed]
- Mumby, P.J.; Harborne, A.R.; Williams, J.; Kappel, C.V.; Brumbaugh, D.R.; Micheli, F.; Holmes, K.E.; Dahlgren, C.P.; Paris, C.B.; Blackwell, P.G. Trophic cascade facilitates coral recruitment in a marine reserve. Proc. Natl. Acad. Sci. USA 2007, 104, 8362–8367. [Google Scholar] [CrossRef]
- Knowlton, N.; Jackson, J.B.C. Shifting baselines, local impacts, and global change on coral reefs. PLoS Biol. 2008, 6, e54. [Google Scholar] [CrossRef]
- Adam, T.C.; Burkepile, D.E.; Holbrook, S.J.; Carpenter, R.C.; Claudet, J.; Loiseau, C.; Thiault, L.; Brooks, A.J.; Washburn, L.; Schmitt, R.J. Landscape-scale patterns of nutrient enrichment in a coral reef ecosystem: Implications for coral to algae phase shifts. Ecol. Appl. 2021, 31, e2227. [Google Scholar] [CrossRef]
- Knowlton, N. Multiple “stable” states and the conservation of marine ecosystems. Prog. Oceanogr. 2004, 60, 387–396. [Google Scholar] [CrossRef]
- Kuffner, I.; Walters, L.; Becerro, M.; Paul, V.; Ritson-Williams, R.; Beach, K. Inhibition of coral recruitment by macroalgae and cyanobacteria. Mar. Ecol. Prog. Ser. 2006, 323, 107–117. [Google Scholar] [CrossRef]
- Ling, S.D.; Johnson, C.R.; Frusher, S.D.; Ridgway, K.R. Overfishing reduces resilience of kelp beds to climate-driven catastrophic phase shift. Proc. Natl. Acad. Sci. USA 2009, 106, 22341–22345. [Google Scholar] [CrossRef]
- Filbee-Dexter, K.; Scheibling, R.E. Sea urchin barrens as alternative stable states of collapsed kelp ecosystems. Mar. Ecol. Prog. Ser. 2014, 495, 1–25. [Google Scholar] [CrossRef]
- Cavanaugh, K.C.; Kellner, J.R.; Forde, A.J.; Gruner, D.S.; Parker, J.D.; Rodriguez, W.; Feller, I.C. Poleward expansion of mangroves is a threshold response to decreased frequency of extreme cold events. Proc. Natl. Acad. Sci. USA 2014, 111, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Cavanaugh, K.C.; Dangremond, E.M.; Doughty, C.L.; Park Williams, A.; Parker, J.D.; Hayes, M.A.; Rodriguez, W.; Feller, I.C. Climate-driven regime shifts in a mangrove–salt marsh ecotone over the past 250 years. Proc. Natl. Acad. Sci. USA 2019, 116, 21602–21608. [Google Scholar] [CrossRef]
- Giri, C.; Long, J. Is the geographic range of mangrove forests in the conterminous United States really expanding? Sensors 2016, 16, 2010. [Google Scholar] [CrossRef] [PubMed]
- Osland, M.J.; Day, R.H.; Hall, C.T.; Brumfield, M.D.; Dugas, J.L.; Jones, W.R. Mangrove expansion and contraction at a poleward range limit: Climate extremes and land-ocean temperature gradients. Ecology 2017, 98, 125–137. [Google Scholar] [CrossRef]
- Osland, M.J.; Hartmann, A.M.; Day, R.H.; Ross, M.S.; Hall, C.T.; Feher, L.C.; Vervaeke, W.C. Microclimate influences mangrove freeze damage: Implications for range expansion in response to changing macroclimate. Estuaries Coasts 2019, 42, 1084–1096. [Google Scholar] [CrossRef]
- Osland, M.J.; Day, R.H.; Hall, C.T.; Feher, L.C.; Armitage, A.R.; Cebrian, J.; Dunton, K.H.; Hughes, A.R.; Kaplan, D.A.; Langston, A.K.; et al. Temperature thresholds for black mangrove (Avicennia germinans) freeze damage, mortality and recovery in North America: Refining tipping points for range expansion in a warming climate. J. Ecol. 2020, 108, 654–665. [Google Scholar] [CrossRef]
- Osland, M.J.; Day, R.H.; Michot, T.C. Frequency of extreme freeze events controls the distribution and structure of black mangroves (Avicennia germinans) near their northern range limit in coastal Louisiana. Divers. Distrib. 2020, 26, 1366–1382. [Google Scholar] [CrossRef]
- Steinmuller, H.E.; Foster, T.E.; Boudreau, P.; Ross Hinkle, C.; Chambers, L.G. Tipping points in the mangrove march: Characterization of biogeochemical cycling along the mangrove–salt marsh ecotone. Ecosystems 2020, 23, 417–434. [Google Scholar] [CrossRef]
- Milbrandt, E.C.; Tinsley, M.N. The role of saltwort (Batis maritima) in regeneration of degraded mangrove forests. Hydrobiologia 2006, 568, 369–377. [Google Scholar] [CrossRef]
- McKee, K.L.; Rooth, J.E.; Feller, I.C. Mangrove recruitment after forest disturbance is facilitated by herbaceous species in the Caribbean. Ecol. Appl. 2007, 17, 1678–1693. [Google Scholar] [CrossRef]
- Peterson, J.M.; Bell, S.S. Tidal events and salt-marsh structure influence black mangrove (Avicennia germinans) recruitment across an ecotone. Ecology 2012, 93, 1648–1658. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, M.; Walters, L. Trapping of Rhizophora mangle by coexisting early successional species. Estuaries Coasts 2014, 37, 1562–1571. [Google Scholar] [CrossRef]
- Stevens, P.W.; Fox, S.L.; Montague, C.L. The interplay between mangroves and saltmarshes at the transition between temperate and sub-tropical climate in Florida. Wetl. Ecol. Manag. 2006, 14, 435–444. [Google Scholar] [CrossRef]
- Krauss, K.W.; From, A.S.; Doyle, T.W.; Doyle, T.J.; Barry, M.J. Sea-level rise and landscape change influence mangrove encroachment onto marsh in the Ten Thousand Islands region of Florida, USA. J. Coast. Conserv. 2011, 15, 629–638. [Google Scholar] [CrossRef]
- Osland, M.J.; Spivak, A.C.; Nestlerode, J.A.; Lessmann, J.M.; Almario, A.E.; Heitmuller, P.T.; Russell, M.J.; Krauss, K.W.; Alvarez, F.; Dantin, D.D.; et al. Ecosystem development after mangrove wetland creation: Plant-soil change across a 20-year chronosequence. Ecosystems 2012, 15, 848–866. [Google Scholar] [CrossRef]
- McKee, K.L.; Rooth, J.E. Where temperate meets tropical: Multi-factorial effects of elevated CO2, nitrogen enrichment, and competition on a mangrove-salt marsh community. Glob. Change Biol. 2008, 14, 971–984. [Google Scholar] [CrossRef]
- Osland, M.J.; Enwright, N.; Day, R.H.; Doyle, T.W. Winter climate change and coastal wetland foundation species: Salt marshes vs. mangrove forests in the southeastern United States. Glob. Change Biol. 2013, 19, 1482–1494. [Google Scholar] [CrossRef]
- McClenachan, G.; Witt, M.; Walters, L.J. Replacement of oyster reefs by mangroves: Unexpected climate-driven ecosystem shifts. Glob. Change Biol. 2021, 27, 1226–1238. [Google Scholar] [CrossRef]
- Hesterberg, S.G.; Jackson, K.; Bell, S.S. Climate drives coupled regime shifts across subtropical estuarine ecosystems. Proc. Natl. Acad. Sci. USA 2022, 119, e2121654119. [Google Scholar] [CrossRef]
- Parkinson, R.W. Decelerating Holocene sea-level rise and its influence on southwest Florida coastal evolution: A transgressive/regressive stratigraphy. J. Sediment. Petrol. 1989, 59, 960–972. [Google Scholar] [CrossRef]
- Osland, M.J.; Stevens, P.W.; Lamont, M.M.; Brusca, R.C.; Hart, K.M.; Waddle, J.H.; Langtimm, C.A.; Williams, C.M.; Keim, B.D.; Terando, A.J.; et al. Tropicalization of temperate ecosystems in North America: The northward range expansion of tropical organisms in response to warming winter temperatures. Glob. Change Biol. 2021, 27, 3009–3034. [Google Scholar] [CrossRef]
- Hurst, N.R.; Locher, B.; Steinmuller, H.E.; Walters, L.J.; Chambers, L.G. Organic carbon dynamics and microbial community response to oyster reef restoration. Limnol. Oceanogr. 2022, 67, 1157–1168. [Google Scholar] [CrossRef]
- Balke, T.; Webb, E.; van den Elzen, E.; Galli, D.; Herman, P.; Bouma, T. Seedling establishment in a dynamic sedimentary environment: A conceptual framework using mangroves. J. Appl. Ecol. 2013, 50, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Brockmeyer, R.K.; Radabaugh, K.; Mitchell, J.; Simpson, L.; Sweat, H.; Donnelly, M. Indian River Lagoon. In Coastal Habitat Integrated Mapping and Monitoring Program; Report for the State of Florida No. 2; Florida Fish and Wildlife Conservation Commission: St. Petersburg, FL, USA; Fish and Wildlife Research Institute: St. Petersburg, FL, USA, 2022. [Google Scholar]
- Phlips, E.; Badylak, S.; Lasi, M.; Chamberlain, R.; Green, W.; Hall, L.; Hart, J.; Lockwood, J.; Miller, J.; Morris, L.; et al. From red tides to green and brown tides: Bloom dynamics in a restricted subtropical lagoon under shifting climatic conditions. Estuaries Coasts 2015, 38, 886–904. [Google Scholar] [CrossRef]
- Walters, L.J.; Sacks, P.E.; Campbell, D.E. Boating impacts and boat-wake resilient restoration of the eastern oyster Crassostrea virginica in Mosquito Lagoon, Florida, USA. Fla. Sci. 2021, 84, 173–199. [Google Scholar]
- Smith, N. Tidal and nontidal flushing of Florida’s Indian River Lagoon. Estuaries 1993, 16, 739–746. [Google Scholar] [CrossRef]
- Steward, J.; Virnstein, R.; Lasi, M.; Morris, L.; Miller, J.; Hall, L.; Tweedale, W. The impacts of the 2004 hurricanes on hydrology, water quality, and seagrass in the Central Indian River Lagoon, Florida. Estuaries Coasts 2006, 29, 954–965. [Google Scholar] [CrossRef]
- United States Geological Survey. 2023. Available online: https://waterdata.usgs.gov/monitoring-location/02248380/#parameterCode=00065&period=P7D&showMedian=false (accessed on 20 August 2023).
- Kennedy, V.; Newell, R.; Eble, A.; Leffler, M.; Harpe, S.R.D. The Eastern Oyster: Crassostrea virginica; Maryland Sea Grant College, University of Maryland: College Park, MD, USA, 1996. [Google Scholar]
- Copertino, J.L.; Harris, K.; Chute, L.; Walters, L.J. Impact of oyster (Crassostrea virginica) reef restoration on benthic invertebrates and coastal birds in a subtropical estuary. Sustainability 2022, 14, 2371. [Google Scholar] [CrossRef]
- Searles, A.R.; Gipson, E.E.; Walters, L.J.; Cook, G.S. Oyster reef restoration facilitates the recovery of macroinvertebrate abundance, diversity, and composition in estuarine communities. Nat. Sci. Rep. 2022, 12, 8163. [Google Scholar] [CrossRef]
- Troast, B.V.; Walters, L.J.; Cook, G.S. A multi-tiered assessment of fish community responses to habitat restoration in a coastal lagoon. Mar. Ecol. Prog. Ser. 2022, 698, 1–14. [Google Scholar] [CrossRef]
- Kibler, K.M.; Kitsikoudis, V.; Donnelly, M.; Spiering, D.W.; Walters, L. Flow–vegetation interaction in a living shoreline restoration and potential effect to mangrove recruitment. Sustainability 2019, 11, 3215. [Google Scholar] [CrossRef]
- McClenachan, G.M.; Donnelly, M.J.; Shaffer, M.N.; Sacks, P.E.; Walters, L.J. Does size matter? Quantifying the cumulative impact of small-scale living shoreline and oyster reef restoration projects on shoreline erosion. Restor. Ecol. 2020, 28, 1365–1371. [Google Scholar] [CrossRef]
- Chambers, L.G.; Gaspar, S.A.; Pilato, C.J.; Steinmuller, H.E.; McCarthy, K.J.; Sacks, P.E.; Walters, L.J. How well do restored intertidal oyster reefs support key biogeochemical properties in a coastal lagoon? Estuaries Coasts 2018, 41, 784–799. [Google Scholar] [CrossRef]
- Fodrie, F.J.; Rodriguez, A.B.; Gittman, R.K.; Grabowski, J.H.; Lindquist, N.L.; Peterson, C.H.; Piehler, M.F.; Ridge, J.T. Oyster reefs as carbon sources and sinks. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170891. [Google Scholar] [CrossRef]
- Walters, L.J.; Craig, C.A.; Dark, E.; Wayles, J.; Encomio, V.; Coldren, G.; Sailor-Tynes, T.; Fox, D.W.; Zhai, L. Quantifying spatial and temporal trends of microplastic pollution in surface water and in the Eastern oyster Crassostrea virginica for a dynamic Florida estuary. Environments 2022, 9, 131. [Google Scholar] [CrossRef]
- Rifenberg, J.; Litwak, J.; Fillyaw, R. Vertebrate impact on a newly deployed shoreline stabilization project by wildlife camera analysis. Pegasus Rev. UCF Undergrad. Res. J. 2021, 13, 3. Available online: https://stars.library.ucf.edu/urj/vol13/iss2/3 (accessed on 1 January 2024).
- Kibler, K.M.; Pilato, C.; Walters, L.J.; Donnelly, M.; Taye, J. Hydrodynamic limitations to mangrove seedling retention in subtropical estuaries. Sustainability 2022, 14, 8605. [Google Scholar] [CrossRef]
- Bouillon, S.; Borges, A.V.; Castañeda-Moya, E.; Diele, K.; Dittmar, T.; Duke, N.C.; Kristensen, E.; Lee, S.Y.; Marchand, C.; Middelburg, J.J.; et al. Mangrove production and carbon sinks: A revision of global budget estimates. Glob. Biogeochem. Cycles 2008, 22, GB2013. [Google Scholar] [CrossRef]
- Breithaupt, J.L.; Smoak, J.M.; Smith, T.J.; Sanders, C.J.; Hoare, A. Organic carbon burial rates in mangrove sediments: Strengthening the global budget. Glob. Biogeochem. Cycles 2012, 26, GB3011. [Google Scholar] [CrossRef]
- Breithaupt, J.L.; Smoak, J.M.; Sanders, C.J.; Troxler, T.G. Spatial variability of organic carbon, CaCO3 and nutrient burial rates spanning a mangrove productivity gradient in the coastal Everglades. Ecosystems 2019, 22, 844–858. [Google Scholar] [CrossRef]
- Benson, G.W.; Donnelly, M.J.; Sacks, P.E.; Walters, L.J. Documenting loss and fragmentation of intertidal oyster (Crassostrea virginica) reefs in a subtropical estuary. Environments 2023, 10, 133. [Google Scholar] [CrossRef]
- Garvis, S.; Sacks, P.; Walters, L. Assessing the formation, movement, and restoration of dead intertidal oyster reefs over time using remote sensing in Canaveral National Seashore and Mosquito Lagoon, Florida. J. Shellfish Res. 2015, 34, 251–258. [Google Scholar] [CrossRef]
- Fillyaw, R.M.; Donnelly, M.J.; Litwak, J.W.; Rifenberg, J.L.; Walters, L.J. Strategies for successful mangrove living shoreline stabilizations in shallow water subtropical estuaries. Sustainability 2021, 13, 11704. [Google Scholar] [CrossRef]
- Friess, D.A.; Krauss, K.W.; Horstman, E.M.; Balke, T.; Bouma, T.J.; Galli, D.; Webb, E.L. Are all intertidal wetlands naturally created equal? Bottlenecks, thresholds and knowledge gaps to mangrove and saltmarsh ecosystems. Biol. Rev. 2012, 87, 346–366. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.R. Ecological engineering for successful management and restoration of mangrove forests. Ecol. Eng. 2005, 24, 403–418. [Google Scholar] [CrossRef]
- McKee, K.L. Soil physicochemical patterns and mangrove species distribution—Reciprocal effects? J. Ecol. 1993, 81, 477–487. [Google Scholar] [CrossRef]
- McKee, K.L. Growth and physiological responses of neotropical mangrove seedlings to root zone hypoxia. Tree Physiol. 1996, 16, 883–889. [Google Scholar] [CrossRef]
- Elster, C. Reasons for reforestation success and failure with three mangrove species in Colombia. For. Ecol. Manag. 2000, 131, 201–214. [Google Scholar] [CrossRef]
- Baggett, L.; Powers, S.; Brumbaugh, R.; Coen, L.; DeAngelis, B.; Greene, J.; Hancock, B.; Morlock, S.; Allen, B.; Breitburg, D.; et al. Guidelines for evaluating performance of oyster habitat restoration. Restor. Ecol. 2015, 23, 737–745. [Google Scholar] [CrossRef]
- Parker, M.L.; Arnold, W.S.; Geiger, S.P.; Gorman, P.; Leone, E.H. Impacts of freshwater management activities on eastern oyster (Crassostrea virginica) density and recruitment: Recovery and long-term stability in seven Florida estuaries. J. Shellfish Res. 2013, 32, 695–708. [Google Scholar] [CrossRef]
- McFarland, K.; Vignier, J.; Standen, E.; Volety, A.K. Synergistic effects of salinity and temperature on the eastern oyster Crassostrea virginica throughout the lifespan. Mar. Ecol. Prog. Ser. 2022, 700, 111–124. [Google Scholar] [CrossRef]
- Dangremond, E.M.; Feller, I.C.; Sousa, W.P. Environmental tolerances of rare and common mangroves along light and salinity gradients. Oecologia 2015, 179, 1187–1198. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Therneau, T. A Package for Survival Analysis in R, R Package Version 3.7-0; R Core Team: Vienna, Austria, 2024; Available online: https://CRAN.R-project.org/package=survival (accessed on 10 January 2024).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 10 January 2024).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 10 January 2024).
- Suchonic, E. Climate Change and Coastal Development Impacts on Oyster Abundances in Mosquito Lagoon, FL. Master’s Thesis, University of Central Florida, Orlando, FL, USA, 28 May 2024. Available online: https://stars.library.ucf.edu/etd2023/182 (accessed on 1 June 2024).
- Smith, T.J., III. Seed predation in relation to tree dominance and distribution in mangrove forests. Ecology 1987, 68, 266–273. [Google Scholar] [CrossRef]
- Smith, T.J., III; Chan, H.T.; McIvor, C.C.; Robblee, M.B. Comparisons of seed predation in tropical, tidal forests from three continents. Ecology 1989, 70, 146–151. [Google Scholar] [CrossRef]
- McGuinness, K. Seed predation in a tropical mangrove forest: A test of the dominance-predation model in northern Australia. J. Trop. Ecol. 1997, 13, 293–302. [Google Scholar] [CrossRef]
- Devaney, J.L.; Lehmann, M.; Feller, I.C.; Parker, J.D. Mangrove microclimates alter seedling dynamics at the range edge. Ecology 2017, 98, 2513–2520. [Google Scholar] [CrossRef]
- Van der Stocken, T.; Wee, A.K.S.; De Ryck, D.J.R.; Vanschoenwinkel, B.; Friess, D.A.; Dahdouh-Guebas, F.; Simard, M.; Koedam, N.; Webb, E.L. A general framework for propagule dispersal in mangroves. Biol. Rev. 2019, 94, 1547–1575. [Google Scholar] [CrossRef]
- Ellison, J.C. Impacts of sediment burial on mangroves. Mar. Pollut. Bull. 1999, 37, 420–426. [Google Scholar] [CrossRef]
- Phillips, D.H.; Kumara, M.P.; Jayatissa, L.P.; Krauss, K.W.; Huxham, M. Impacts of Mangrove Density on Surface Sediment Accretion, Belowground Biomass and Biogeochemistry in Puttalam Lagoon, Sri Lanka. Wetlands 2017, 37, 471–483. [Google Scholar] [CrossRef]
- Kumara, M.P.; Jayatissa, L.P.; Krauss, K.W.; Phillips, D.H.; Huxham, M. High mangrove density enhances surface accretion, surface elevation change, and tree survival in coastal areas susceptible to sea-level rise. Oecologia 2010, 164, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Lenihan, H.S. Physical–biological coupling on oyster reefs: How habitat structure influences individual performance. Ecol. Monogr. 1999, 69, 251–275. [Google Scholar]
- Colden, A.M.; Lipcius, R.N. Lethal and sublethal effects of sediment burial on the eastern oyster Crassostrea virginica. Mar. Ecol. Prog. Ser. 2015, 527, 105–117. [Google Scholar] [CrossRef]
- Poirier, L.A.; Clements, J.C.; Coffin, M.R.; Craig, T.; Davidson, J.; Miron, G.; Davidson, J.D.; Hill, J.; Comeau, L.A. Siltation negatively affects settlement and gaping behaviour in eastern oysters. Mar. Environ. Res. 2021, 170, 105432. [Google Scholar] [CrossRef] [PubMed]
- Middelberg, J.; Soetaert, K.; Herman, P.; Heip, C. Denitrification in marine sediments: A model study. Glob. Biogeochem. Cycles 1996, 10, 661–673. [Google Scholar] [CrossRef]
- Clark, M.W.; McConchie, D.; Lewis, D.W.; Saenger, P. Redox stratification and heavy metal partitioning in Avicennia-dominated mangrove sediments: A geochemical model. Chem. Geol. 1998, 149, 147–171. [Google Scholar] [CrossRef]
- Marchand, C.; Lallier-Vergès, E.; Baltzer, F. The composition of sedimentary organic matter in relation to the dynamic features of a mangrove-fringed coast in French Guiana. Estuar. Coast. Shelf Sci. 2003, 56, 119–130. [Google Scholar] [CrossRef]
- Marchand, C.; Baltzer, F.; Lallier-Verges, E.; Alberic, P. Porewater chemistry in mangrove sediments: Relationship with species composition and developmental stages (French Guiana). Mar. Geol. 2004, 208, 361–381. [Google Scholar] [CrossRef]
- Dakora, F.D.; Phillips, D.A. Root exudates as mediators of mineral acquisition in low-nutrient environments. In Food Security in Nutrient-Stressed Environments: Exploiting Plants’ Genetic Capabilities; Adu-Gyamfi, J.J., Ed.; Developments in Plant and Soil Sciences; Springer: Dordrecht, The Netherlands, 2002; Volume 95, pp. 201–213. [Google Scholar] [CrossRef]
- Beniash, E.; Ivanina, A.; Lieb, N.S.; Kuronchkin, I.; Sokolova, I.M. Elevated level of carbon dioxide affects metabolism and shell formation in oysters Crassostrea virginica. Mar. Ecol. Prog. Ser. 2010, 419, 95–108. [Google Scholar] [CrossRef]
- Waldbusser, G.G.; Steenson, R.A.; Green, M.A. Oyster shell dissolution rates in estuarine waters: Effects of pH and shell legacy. J. Shellfish Res. 2011, 30, 659–669. [Google Scholar] [CrossRef]
- Keppel, A.G.; Breitburg, D.L.; Burrell, R.B. Effects of co-varying diel-cycling hypoxia and pH on growth in the juvenile Eastern oyster, Crassostrea virginica. PLoS ONE 2016, 11, e0161088. [Google Scholar] [CrossRef]
- Reef, R.; Feller, I.C.; Lovelock, C.E. Nutrition of mangroves. Tree Physiol. 2010, 30, 1148–1160. [Google Scholar] [CrossRef]
- Kellogg, M.L.; Cornwell, J.C.; Owens, M.S.; Paynter, K.T. Denitrification and nutrient assimilation on a restored oyster reef. Mar. Ecol. Prog. Ser. 2013, 480, 1–19. [Google Scholar] [CrossRef]
- Malhotra, H.; Vandana, S.S.; Pandey, R. Phosphorus nutrition: Plant growth in response to deficiency and excess. In: Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds. Plant Nutr. Abiotic Stress Toler. 2018, 7, 171–190. [Google Scholar]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Positive and negative effects of organisms as physical ecosystem engineers. Ecology 1997, 78, 1946–1957. [Google Scholar] [CrossRef]
- McAfee, D.; Bishop, M.J. The mechanisms by which oysters facilitate invertebrates vary across environmental gradients. Oecologia 2019, 189, 1095–1106. [Google Scholar] [CrossRef]
- Drexler, M.; Parker, M.L.; Geiger, S.P.; Arnold, W.S.; Hallock, P. Biological assessment of eastern oysters (Crassostrea virginica) inhabiting reef, mangrove, seawall, and restoration substrates. Estuaries Coasts 2014, 37, 962–972. [Google Scholar] [CrossRef]
- Aquino-Thomas, J.; Proffitt, C.E. Oysters (Crassostrea virginica) on red mangrove Rhizophora mangle prop roots: Facilitation of one foundation species by another. Mar. Ecol. Prog. Ser. 2014, 503, 177–194. [Google Scholar] [CrossRef]
- Bishop, M.J.; Byers, J.E.; Marcek, B.J.; Gribben, P.E. Density-dependent facilitation cascades determine epifaunal community structure in temperate Australian mangroves. Ecology 2012, 93, 1388–1401. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, J.H.; Brumbaugh, R.D.; Conrad, R.F.; Keeler, A.G.; Opaluch, J.J.; Peterson, C.H.; Piehler, M.F.; Powers, S.P.; Smyth, A.R. Economic valuation of ecosystem services provided by oyster reefs. Bioscience 2012, 62, 900–909. [Google Scholar] [CrossRef]
- East Central Florida Regional Planning Council Treasure Coast Regional Planning Council. Indian River Lagoon Economic Valuation Update Final Report; Treasure Coast Regional Planning Council: Stuart, FL, USA, 2016; Available online: https://files.tcrpc.org/portfolio%20of%20work/Economic%20Development/IRL%20Valuation/FinalReportIRL08_26_2016.pdf (accessed on 10 June 2024).
- Indian River Lagoon National Estuary Program. Comprehensive Conservation and Management Plan; The Indian River Lagoon NEP: Sebastian, FL, USA, 2019; Available online: https://onelagoon.org/wp-content/uploads/IRLNEP_Final-Draft-CCMP-REVISION_2018-12-07_LowRes__20200204.pdf (accessed on 1 June 2024).
- The Nature Conservancy. Valuing the Flood Risk Reduction Benefits of Florida’s Mangroves; Nature Conservancy: Mountainair, NM, USA, 2017; Available online: https://www.conservationgateway.org/SiteAssets/Pages/floridamangroves/Mangrove_Report_digital_FINAL.pdf (accessed on 1 June 2024).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Red Mangroves | Black Mangroves | White Mangroves | All Mangroves | |
|---|---|---|---|---|
| Number tracked | 1523 (90.6%) | 157 (9.3%) | 1 (0.1%) | 1681 (100%) |
| Number alive at month 15 | 201 (13.2%) | 1 (0.6%) | 0 (0.0%) | 202 (12.0%) |
| Arrival months | All months except May | December–March | October | All months except May |
| Number that produced leaves by month 15 | 168 (cohort 1 only) | 23 (cohort 2 only) | 1 (cohort 1 only) | 192 |
| Month of initial leaf production | All months of year | January–August | December | All months of the year |
| Number alive at month 15 with no leaf production | 30 (cohort 1 only) | 0 | 0 | 30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walters, L.J.; Sacks, P.E.; Harris, K.; McClenachan, G. Ecological Shifts: Plant Establishment in an Animal-Based Ecosystem. Environments 2024, 11, 193. https://doi.org/10.3390/environments11090193
Walters LJ, Sacks PE, Harris K, McClenachan G. Ecological Shifts: Plant Establishment in an Animal-Based Ecosystem. Environments. 2024; 11(9):193. https://doi.org/10.3390/environments11090193
Chicago/Turabian StyleWalters, Linda J., Paul E. Sacks, Katherine Harris, and Giovanna McClenachan. 2024. "Ecological Shifts: Plant Establishment in an Animal-Based Ecosystem" Environments 11, no. 9: 193. https://doi.org/10.3390/environments11090193