
Assessing the Resilience of Enteric Bacteria in Manure in Response to Changes in Relative Humidity and UV-B Light

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.1.1. UV-B
2.1.2. Relative Humidity
2.2. Bacterial Analysis
2.3. Statistical Analysis
Imputation of Missing Counts
3. Results
3.1. Descriptive Statistics
3.1.1. UV-B
3.1.2. Relative Humidity
3.2. Quantitative Bacterial Dynamics
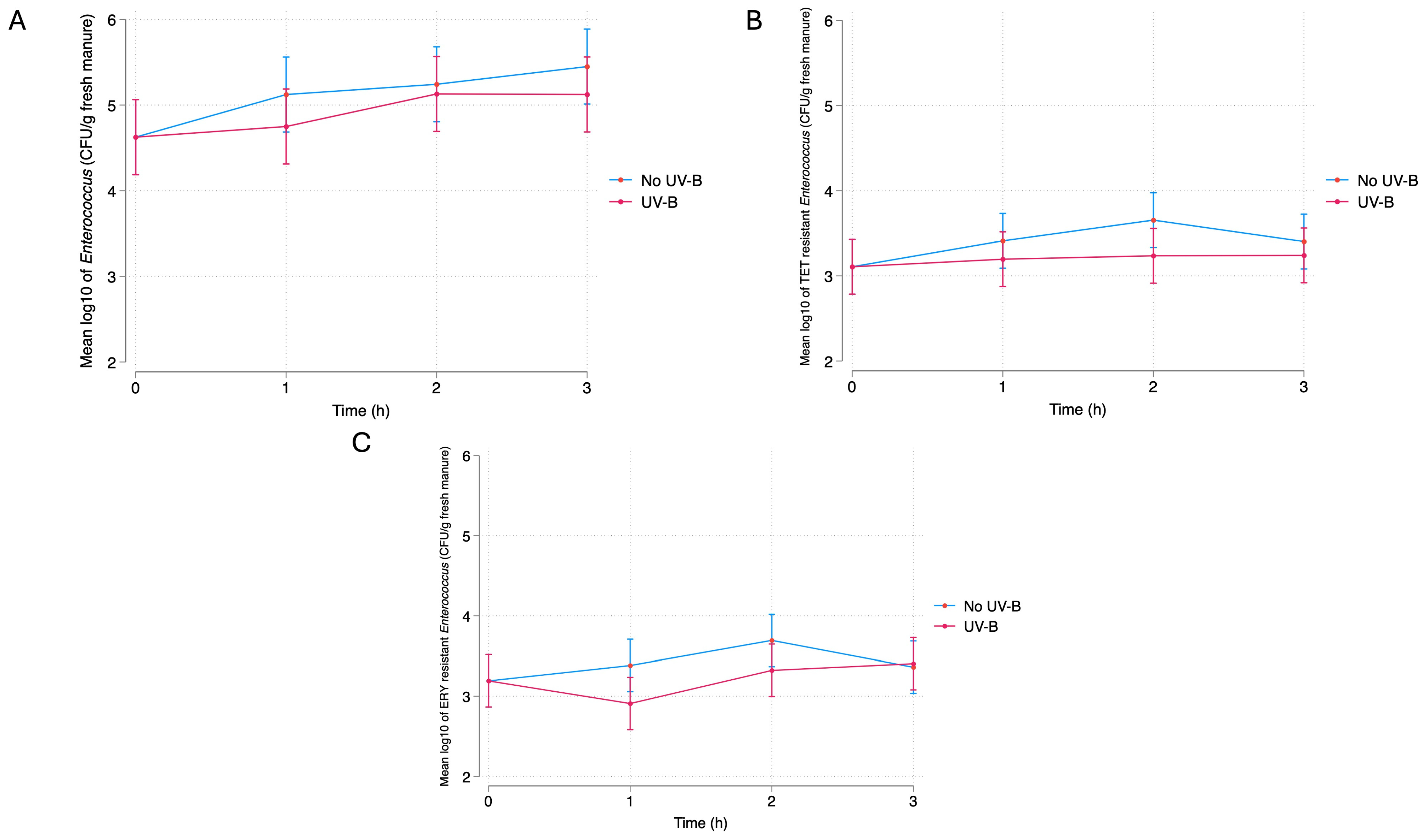
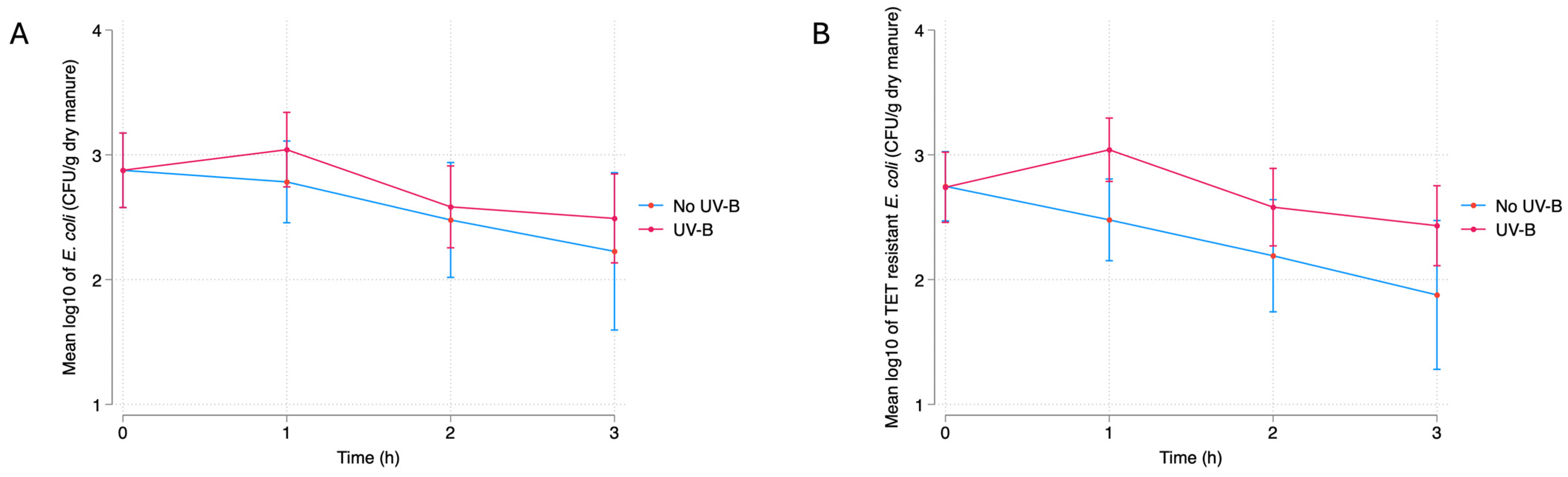
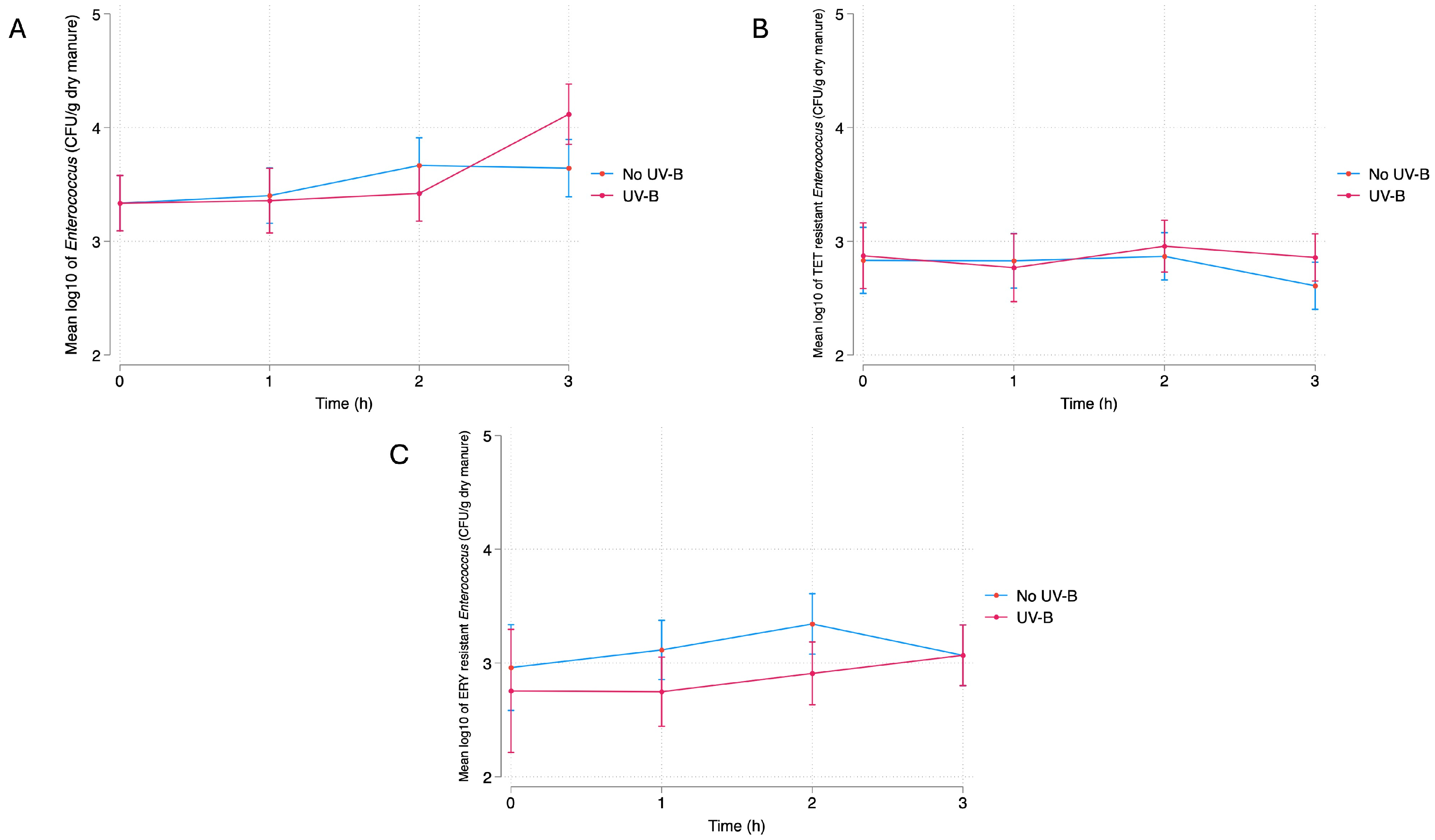
3.2.1. UV-B
Fresh Manure Bacterial Quantification
Dry Manure
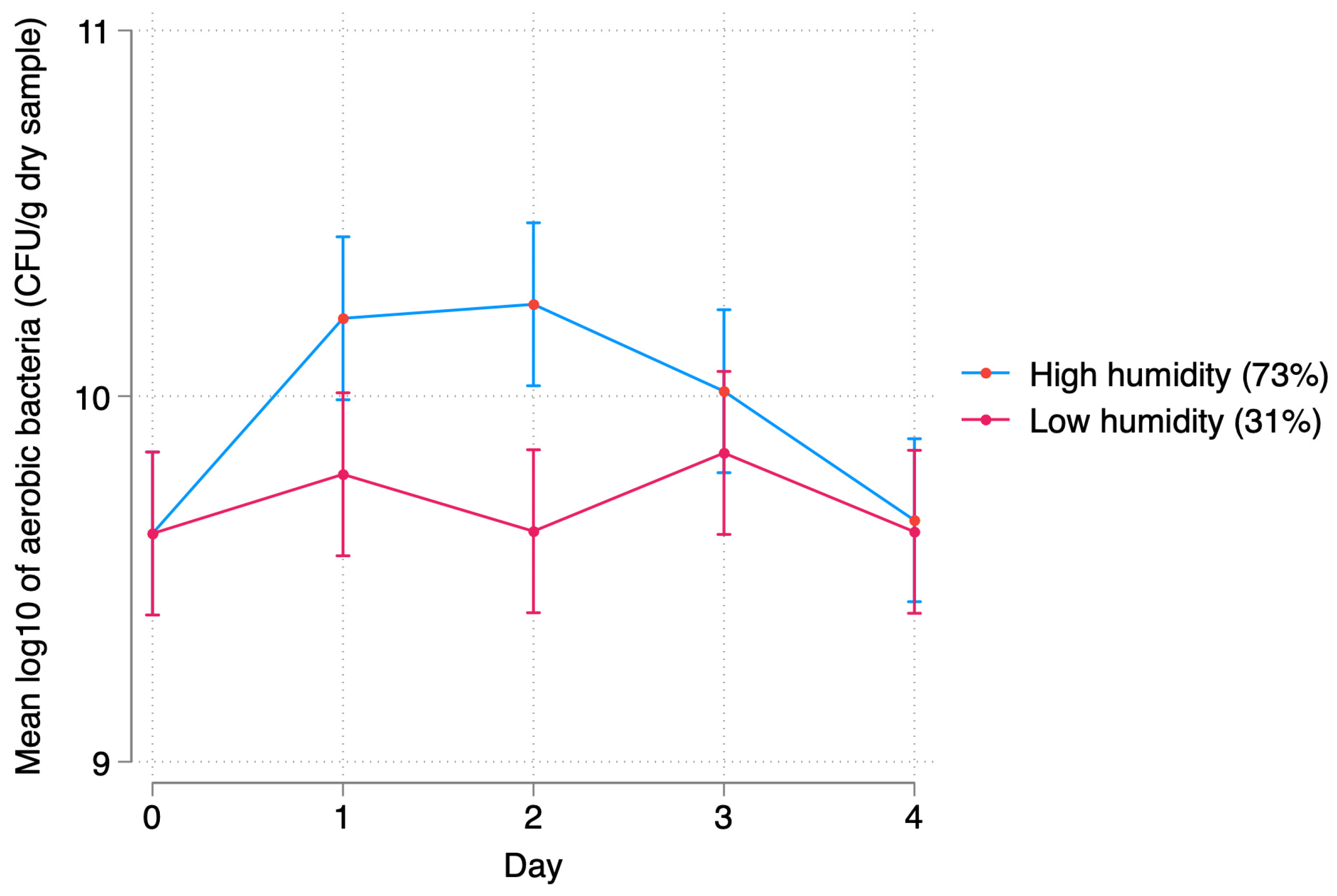
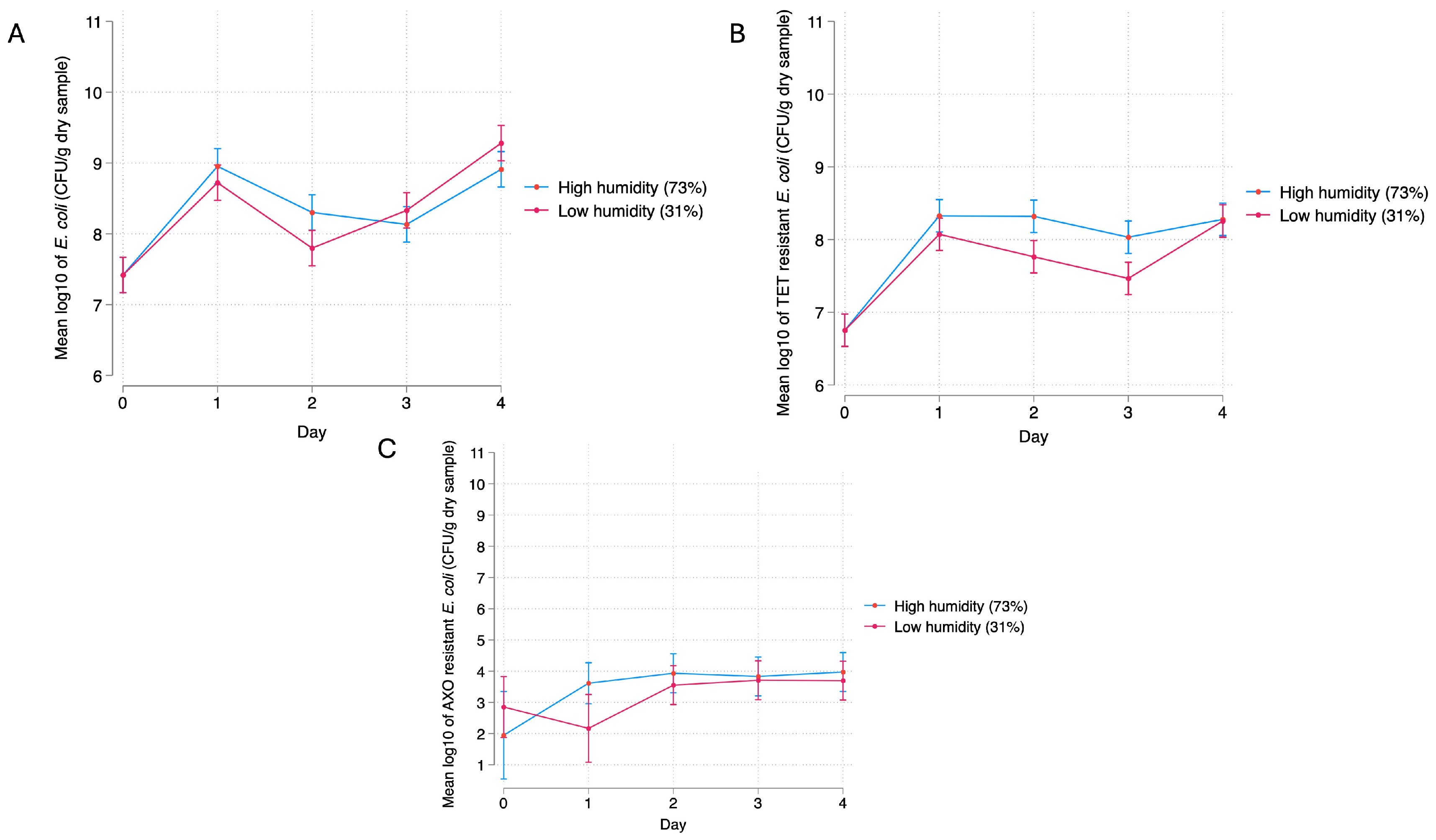
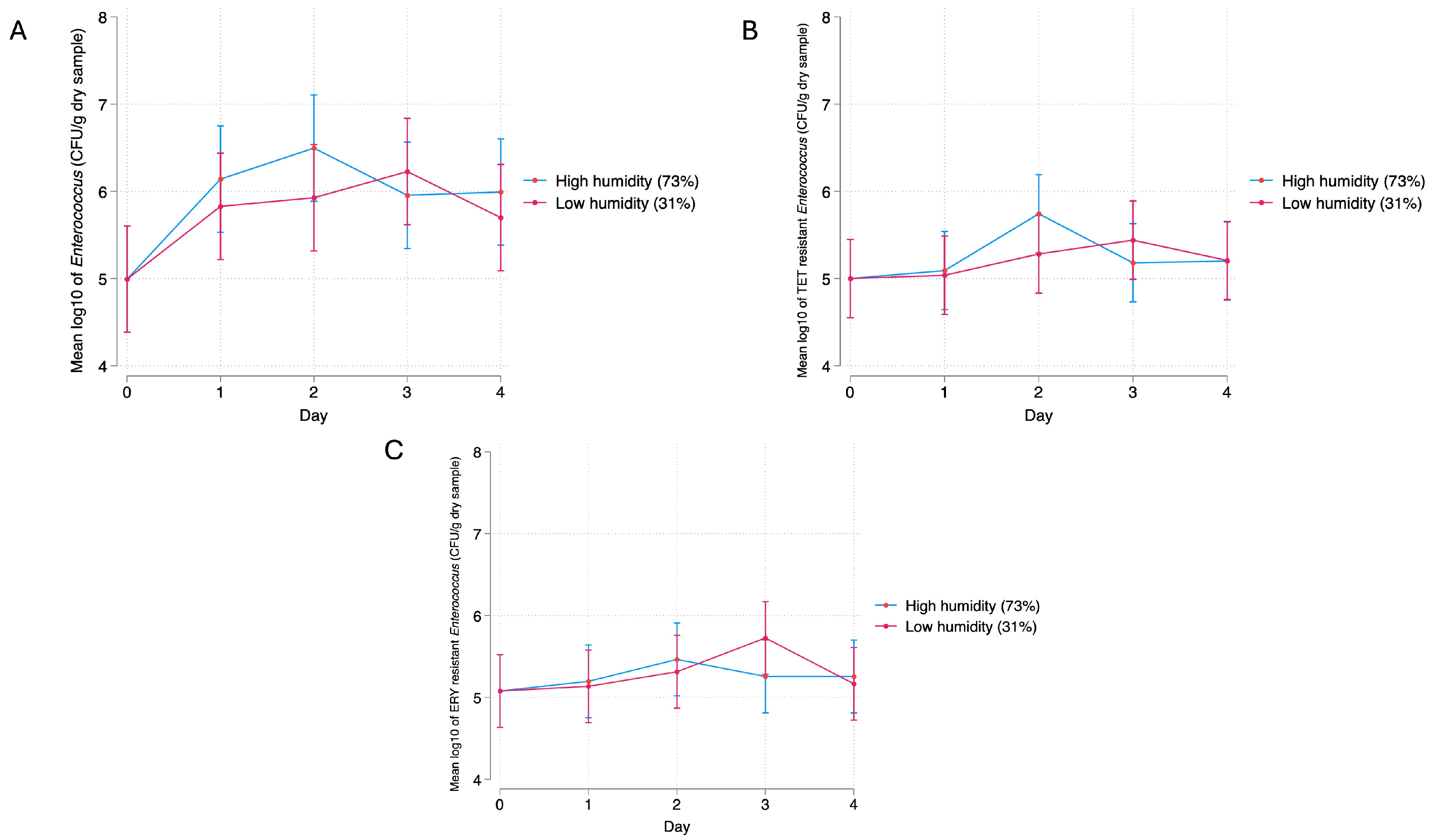
3.2.2. Relative Humidity
4. Discussion
4.1. Bacterial Response to UV-B Exposure
4.1.1. Fresh Manure
4.1.2. Dry Manure
4.2. Bacterial Response to Relative Humidity
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, D.; Chai, T.; Xia, X.; Gao, Y.; Cai, Y.; Li, X.; Miao, Z.; Sun, L.; Hao, H.; Roesler, U.; et al. Formation and transmission of Staphylococcus aureus (including MRSA) aerosols carrying antibiotic-resistant genes in a poultry farming environment. Sci. Total Environ. 2012, 426, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States; CDC: Atlanta, GA, USA, 2019; p. 150. [Google Scholar]
- Stanton, I.C.; Bethel, A.; Leonard, A.F.C.; Gaze, W.H.; Garside, R. What is the research evidence for antibiotic resistance exposure and transmission to humans from the environment? A systematic map protocol. Environ. Evid. 2020, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Antibiotic Resistance and NARMS Surveillance. Available online: https://www.cdc.gov/narms/faq.html#:~:text=NARMS%20scientists%20track%20and%20study,person%20contact%2C%20and%20environmental%20contamination. (accessed on 19 June 2024).
- Bokhary, H.; Pangesti, K.N.A.; Rashid, H.; Abd El Ghany, M.; Hill-Cawthorne, G.A. Travel-Related Antimicrobial Resistance: A Systematic Review. Trop. Med. Infect. Dis. 2021, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Allcock, S.; Young, E.H.; Holmes, M.; Gurdasani, D.; Dougan, G.; Sandhu, M.S.; Solomon, L.; Török, M.E. Antimicrobial resistance in human populations: Challenges and opportunities. Glob. Health Epidemiol. Genom. 2017, 2, e4. [Google Scholar] [CrossRef]
- UN Environment Programme. Frontiers 2017: Emerging Issues of Environmental Concern; UN: Nairobi, Kenya, 2017; p. 11. [Google Scholar]
- U.S. Food and Drug Administration (FDA). About NARMS. Available online: https://www.fda.gov/animal-veterinary/national-antimicrobial-resistance-monitoring-system/about-narms#bacteria (accessed on 12 March 2024).
- National Antimicrobial Resistance Monitoring System. 2018 NARMS Update: Integrated Report Summary Interactive Version; FDA: Silver Spring, MD, USA, 2018. Available online: https://www.fda.gov/animal-veterinary/national-antimicrobial-resistance-monitoring-system/2018-narms-update-integrated-report-summary (accessed on 11 December 2020).
- Aarestrup, F.M.; Wegener, H.C.; Collignon, P. Resistance in bacteria of the food chain: Epidemiology and control strategies. Expert. Rev. Anti Infect. Ther. 2008, 6, 733–750. [Google Scholar] [CrossRef]
- Byappanahalli, M.N.; Nevers, M.B.; Korajkic, A.; Staley, Z.R.; Harwood, V.J. Enterococci in the environment. Microbiol. Mol. Biol. Rev. MMBR 2012, 76, 685–706. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration (FDA). 2020 Summary Report On Antimicrobials Sold or Distributed for Use in Food-Producing Animals; FDA: Silver Spring, MD, USA, 2021; p. 49. [Google Scholar]
- United States Department of Agriculture, Veterinary Services, Centers for Epidemiology and Animal Health. Types and Costs of Respiratory Disease Treatments in U.S. Feedlots; United States Department of Agriculture: Washington, DC, USA, 2013; Volume 2. [Google Scholar]
- Environmental Protection Agency. Risk Assessment Evaluation for Concentrated Animal Feeding Operations; United States Environmental Protection Agency: Washington, DC, USA, 2004; p. 138. [Google Scholar]
- He, L.-Y.; He, L.-K.; Liu, Y.-S.; Zhang, M.; Zhao, J.-L.; Zhang, Q.-Q.; Ying, G.-G. Microbial diversity and antibiotic resistome in swine farm environments. Sci. Total Environ. 2019, 685, 197–207. [Google Scholar] [CrossRef]
- Létourneau, V.; Nehmé, B.; Mériaux, A.; Massé, D.; Cormier, Y.; Duchaine, C. Human pathogens and tetracycline-resistant bacteria in bioaerosols of swine confinement buildings and in nasal flora of hog producers. Int. J. Hyg. Environ. Health 2010, 213, 444–449. [Google Scholar] [CrossRef]
- Dohmen, W.; Schmitt, H.; Bonten, M.; Heederik, D. Air exposure as a possible route for ESBL in pig farmers. Environ. Res. 2017, 155, 359–364. [Google Scholar] [CrossRef]
- Kormos, D.; Lin, K.; Pruden, A.; Marr, L.C. Critical review of antibiotic resistance genes in the atmosphere. Environ. Sci. Process. Impacts 2022, 24, 870–883. [Google Scholar] [CrossRef]
- Dungan, R.S. Board-invited review: Fate and transport of bioaerosols associated with livestock operations and manures. J. Anim. Sci. 2010, 88, 3693–3706. [Google Scholar] [CrossRef] [PubMed]
- Auvermann, B.; Bottcher, R.; Heber, A.; Meyer, D.; B. Parnell, C., Jr.; Shaw, B.; Worley, J. Particulate Matter Emissions from Animal Feeding Operations; American Society of Agricultural and Biological Engineers (ASABE): St. Joseph, MI, USA, 2006. [Google Scholar]
- Himathongkham, S.; Bahari, S.; Riemann, H.; Cliver, D. Survival of Escherichia coli O157:H7 and Salmonella Typhimurium in cow manure and cow manure slurry. FEMS Microbiol. Lett. 1999, 178, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef] [PubMed]
- Lleo, M.M.; Tafi, M.C.; Canepari, P. Nonculturable Enterococcus faecalis cells are metabolically active and capable of resuming active growth. Syst. Appl. Microbiol. 1998, 21, 333–339. [Google Scholar] [CrossRef]
- Vernocchi, V.; Abd El, E.; Brunoldi, M.; Danelli, S.G.; Gatta, E.; Isolabella, T.; Mazzei, F.; Parodi, F.; Prati, P.; Massabò, D. Airborne bacteria viability and air quality: A protocol to quantitatively investigate the possible correlation by an atmospheric simulation chamber. Atmos. Meas. Tech. 2023, 16, 5479–5493. [Google Scholar] [CrossRef]
- Poole, K. Bacterial stress responses as determinants of antimicrobial resistance. J. Antimicrob. Chemother. 2012, 67, 2069–2089. [Google Scholar] [CrossRef] [PubMed]
- Laskowska, E.; Kuczyńska-Wiśnik, D. New insight into the mechanisms protecting bacteria during desiccation. Curr. Genet. 2020, 66, 313–318. [Google Scholar] [CrossRef]
- Mattimore, V.; Battista, J.R. Radioresistance of Deinococcus radiodurans: Functions necessary to survive ionizing radiation are also necessary to survive prolonged desiccation. J. Bacteriol. 1996, 178, 633–637. [Google Scholar] [CrossRef]
- Goyal, K.; Walton, L.J.; Browne, J.A.; Burnell, A.M.; Tunnacliffe, A. Molecular Anhydrobiology: Identifying Molecules Implicated in Invertebrate Anhydrobiosis1. Integr. Comp. Biol. 2005, 45, 702–709. [Google Scholar] [CrossRef]
- Barron, J.C.; Forsythe, S.J. Dry stress and survival time of Enterobacter sakazakii and other Enterobacteriaceae in dehydrated powdered infant formula. J. Food Prot. 2007, 70, 2111–2117. [Google Scholar] [CrossRef]
- Wang, Z.; Zhu, T.; Chen, Z.; Meng, J.; Simpson, D.J.; Gänzle, M.G. Genetic determinants of stress resistance in desiccated Salmonella enterica. Appl. Environ. Microbiol. 2021, 87, e0168321. [Google Scholar] [CrossRef] [PubMed]
- Garmiri, P.; Coles, K.E.; Humphrey, T.J.; Cogan, T.A. Role of outer membrane lipopolysaccharides in the protection of Salmonella enterica serovar Typhimurium from desiccation damage. FEMS Microbiol. Lett. 2008, 281, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Mandal, R.K.; Kwon, Y.M. Global screening of Salmonella enterica serovar Typhimurium genes for desiccation survival. Front. Microbiol. 2017, 8, 1723. [Google Scholar] [CrossRef] [PubMed]
- Nasim, A.; James, A. Life under Conditions of High Irradiation; Academic Press: New York, NY, USA, 1978. [Google Scholar]
- Goldman, R.P.; Travisano, M. Experimental evolution of ultraviolet radiation resistance in Escherichia coli. Evolution 2011, 65, 3486–3498. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Radiation: Ultraviolet (UV) Radiation; WHO: Geneva, Switzerland, 2016. [Google Scholar]
- Brozyna, A.; Zbytek, B.; Granese, J.; Carlson, J.A.; Ross, J.; Slominski, A. Mechanism of UV-related carcinogenesis and its contribution to nevi/melanoma. Expert Rev. Dermatol. 2007, 2, 451–469. [Google Scholar] [CrossRef]
- Chautrand, T.; Souak, D.; Kondakova, T.; Depayras, S.; Machour, N.; Heipieper, H.; Feuilloley, M.; Orange, N.; Poc, C. Air Pollution and Other Environmental Stresses: Gaseous No 2 Exposure Leads to Specific Alterations of Pseudomonas Fluorescens; Casares, J., Longhurst, J., Barnes, J., Eds.; WIT Press: Southampton, UK, 2020; Volume 244. [Google Scholar]
- Goosen, N.; Moolenaar, G.F. Repair of UV damage in bacteria. DNA Repair. 2008, 7, 353–379. [Google Scholar] [CrossRef]
- Alcántara-Díaz, D.; Breña-Valle, M.; Serment-Guerrero, J. Divergent adaptation of Escherichia coli to cyclic ultraviolet light exposures. Mutagenesis 2004, 19, 349–354. [Google Scholar] [CrossRef]
- Davies, R.; Sinskey, A.J. Radiation-resistant mutants of Salmonella Typhimurium LT2: Development and characterization. J. Bacteriol. 1973, 113, 133–144. [Google Scholar] [CrossRef]
- Weigand, M.R.; Sundin, G.W. Long-term effects of inducible mutagenic DNA repair on relative fitness and phenotypic diversification in Pseudomonas cichorii 302959. Genetics 2009, 181, 199–208. [Google Scholar] [CrossRef]
- Wassmann, M.; Moeller, R.; Reitz, G.; Rettberg, P. Adaptation of Bacillus subtilis cells to Archean-like UV climate: Relevant hints of microbial evolution to remarkably increased radiation resistance. Astrobiology 2010, 10, 605–615. [Google Scholar] [CrossRef]
- Álvarez-Molina, A.; de Toro, M.; Ruiz, L.; López, M.; Prieto, M.; Alvarez-Ordóñez, A. Selection for antimicrobial resistance in foodborne pathogens through exposure to UV Light and nonthermal atmospheric plasma decontamination techniques. Appl. Environ. Microbiol. 2020, 86, e00102-20. [Google Scholar] [CrossRef] [PubMed]
- Wexler, A.; Hasegawa, S. Relative humidity-temperature relationships of some saturated salt solutions in the temperature. J. Res. Natl. Bur. Stand. 1954, 53, 19–26. [Google Scholar] [CrossRef]
- Koyama, K.; Hokunan, H.; Hasegawa, M.; Kawamura, S.; Koseki, S. Modeling Stochastic Variability in the Numbers of surviving Salmonella enterica, Enterohemorrhagic Escherichia coli, and Listeria monocytogenes cells at the single-cell level in a desiccated environment. Appl. Environ. Microbiol. 2017, 83, e02974-16. [Google Scholar] [CrossRef]
- Boll, J.M.; Tucker, A.T.; Klein, D.R.; Beltran, A.M.; Brodbelt, J.S.; Davies, B.W.; Trent, M.S. Reinforcing lipid A acylation on the cell surface of Acinetobacter baumannii promotes cationic zntimicrobial peptide resistance and desiccation survival. mBio 2015, 6, e00478-15. [Google Scholar] [CrossRef] [PubMed]
- Zeidler, S.; Müller, V. The role of compatible solutes in desiccation resistance of Acinetobacter baumannii. Microbiologyopen 2019, 8, e00740. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing. CLSI Document M100, 33rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2023. [Google Scholar]
- Murray, S.A.; Holbert, A.C.; Norman, K.N.; Lawhon, S.D.; Sawyer, J.E.; Scott, H.M. Effects of tylosin, a direct-fed microbial and feedlot pen environment on phenotypic resistance among Enterococci isolated from beef cattle feces. Antibiotics 2022, 11, 106. [Google Scholar] [CrossRef]
- Stata. Pwcompare—Pairwise Comparisons; StataCorp LLC.: College Station, TX, USA, 2023; p. 34. [Google Scholar]
- Stata. Mi Impute Chained—Impute Missing Values Using Chained Equations; StataCorp LLC.: College Station, TX, USA, 2023; p. 30. [Google Scholar]
- Harel, O.; Zhou, X.H. Multiple imputation: Review of theory, implementation and software. Stat. Med. 2007, 26, 3057–3077. [Google Scholar] [CrossRef]
- Schafer, J.L. Multiple imputation in multivariate problems when the imputation and analysis models differ. Stat. Neerl. 2003, 57, 19–35. [Google Scholar] [CrossRef]
- Eddings, W.; Marchenko, Y. Diagnostics for multiple imputation in Stata. Stata J. 2012, 12, 353–367. [Google Scholar] [CrossRef]
- Tran, D.T.Q.; Bradbury, M.I.; Ogtrop, F.F.V.; Bozkurt, H.; Jones, B.J.; McConchie, R. Environmental drivers for persistence of Escherichia coli and Salmonella in manure-amended soils: A meta-analysis. J. Food Prot. 2020, 83, 1268–1277. [Google Scholar] [CrossRef]
- Zhao, Y.; Aarnink, A.J.A.; De Jong, M.C.M.; Groot Koerkamp, P.W.G. Airborne microorganisms from livestock production systems and their relation to dust. Crit. Rev. Environ. Sci. Technol. 2014, 44, 1071–1128. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Martin, C. Airborne Infectious Microorganisms. In Encyclopedia of Microbiology, 4th ed.; Schmidt, T.M., Ed.; Academic Press: Cambridge, MA, USA; Oxford, UK, 2019; pp. 52–60. [Google Scholar]
- Baghdadi, M.; Brassard, P.; Godbout, S.; Létourneau, V.; Turgeon, N.; Rossi, F.; Lachance, É.; Veillette, M.; Gaucher, M.L.; Duchaine, C. Contribution of manure-spreading operations to bioaerosols and antibiotic resistance genes’ emission. Microorganisms 2023, 11, 1797. [Google Scholar] [CrossRef]
- Sweeten, J.M. Manure and Wastewater Management for Cattle Feedlots. In Reviews of Environmental Contamination and Toxicology: Continuation of Residue Reviews; Ware, G.W., Ed.; Springer: New York, NY, USA, 2000; pp. 121–153. [Google Scholar]
- Santos, A.L.; Oliveira, V.; Baptista, I.; Henriques, I.; Gomes, N.C.M.; Almeida, A.; Correia, A.; Cunha, Â. Wavelength dependence of biological damage induced by UV radiation on bacteria. Arch. Microbiol. 2013, 195, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Rames, J.; Chaloupecký, V.; Sojková, N.; Bencko, V. An attempt to demonstrate the increased resistance of selected bacterial strains during repeated exposure to UV radiation at 254 nm. Cent. Eur. J. Public Health 1997, 5, 30–31. [Google Scholar] [PubMed]
- Singh, P.; El-Din, M.G.; Ikehata, K.; Craik, S.A.; Bromley, D. UV inactivation of bacteria in raw and pre-treated liquid swine manure. Environ. Technol. 2006, 27, 1261–1270. [Google Scholar] [CrossRef]
- Madge, B.A.; Jensen, J.N. Ultraviolet disinfection of fecal coliform in municipal wastewater: Effects of particle size. Water Environ. Res. 2006, 78, 294–304. [Google Scholar] [CrossRef]
- Macauley, J.J.; Qiang, Z.; Adams, C.D.; Surampalli, R.; Mormile, M.R. Disinfection of swine wastewater using chlorine, ultraviolet light and ozone. Water Res. 2006, 40, 2017–2026. [Google Scholar] [CrossRef]
- Berman, D.; Rice, E.W.; Hoff, J.C. Inactivation of particle-associated coliforms by chlorine and monochloramine. Appl. Environ. Microbiol. 1988, 54, 507–512. [Google Scholar] [CrossRef]
- Kempf, B.; Bremer, E. Uptake and synthesis of compatible solutes as microbial stress responses to high-osmolality environments. Arch. Microbiol. 1998, 170, 319–330. [Google Scholar] [CrossRef]
- Romero-Martínez, L.; Duque-Sarango, P.; González-Martín, C.; Moreno-Andrés, J.; Acevedo-Merino, A.; Nebot, E. Inactivation efficacy and reactivation of fecal bacteria with a flow-through LED ultraviolet reactor: Intraspecific response prevails over interspecific differences. J. Water Process Eng. 2023, 52, 103497. [Google Scholar] [CrossRef]
- Zafar, L.; Javed, A.; Ali, S.S.; Anwar, Z. Potential of waste water sludge as environmental-friendly manure after UV-treatment. Res. J. Environ. Earth Sci. 2012, 4, 917–922. [Google Scholar]
- Zhang, W.; Huang, R.; Zhang, T.; Wang, B.; Li, N.; Sun, Y.; Ma, H.; Zhang, Q.; Zhang, J.; Liu, Y. Study on the inactivation and reactivation mechanism of pathogenic bacteria in aquaculture by UVC-LED. Front. Mar. Sci. 2023, 10, 1139713. [Google Scholar] [CrossRef]
- Crowley, D.J.; Hanawalt, P.C. Induction of the SOS response increases the efficiency of global nucleotide excision repair of cyclobutane pyrimidine dimers, but not 6–4 photoproducts, in UV-irradiated Escherichia coli. J. Bacteriol. 1998, 180, 3345–3352. [Google Scholar] [CrossRef] [PubMed]
- Koch, W.; Woodgate, R. DNA Damage and Repair: DNA Repair in Prokaryotes and Lower Eukaryotes; Humana: Totowa, NJ, USA, 1998. [Google Scholar]
- Huang, J.-J.; Xi, J.; Hu, H.-Y.; Li, Y.; Lu, S.-Q.; Tang, F.; Pang, Y.-C. UV light tolerance and reactivation potential of tetracycline-resistant bacteria from secondary effluents of a wastewater treatment plant. J. Environ. Sci. 2016, 41, 146–153. [Google Scholar] [CrossRef]
- McKenzie, R.; Connor, B.; Bodeker, G. Increased Summertime UV Radiation in New Zealand in Response to Ozone Loss. Science 1999, 285, 1709–1711. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). Solar and Ultraviolet Radiation. Available online: https://www.ncbi.nlm.nih.gov/books/NBK304366/#:~:text=The%20UV%20component%20of%20terrestrial,for%20additional%20exposure%20has%20increased. (accessed on 13 August 2024).
- Davies, J.; Webb, V. Antibiotic resistance in bacteria. In Biomedical Research Reports; Krause, R.M., Ed.; Academic Press: Cambridge, MD, USA, 1998; Volume 1, pp. 239–273. [Google Scholar]
- Tierney, A.R.; Rather, P.N. Roles of two-component regulatory systems in antibiotic resistance. Future Microbiol. 2019, 14, 533–552. [Google Scholar] [CrossRef]
- Liao, X.; Ma, Y.; Daliri, E.B.-M.; Koseki, S.; Wei, S.; Liu, D.; Ye, X.; Chen, S.; Ding, T. Interplay of antibiotic resistance and food-associated stress tolerance in foodborne pathogens. Trends Food Sci. Technol. 2020, 95, 97–106. [Google Scholar] [CrossRef]
- Meckes, M.C. Effect of UV light disinfection on antibiotic-resistant coliforms in wastewater effluents. Appl. Environ. Microbiol. 1982, 43, 371–377. [Google Scholar] [CrossRef]
- Gardner, A.; Soni, A.; Cookson, A.; Brightwell, G. Light tolerance of extended spectrum β-lactamase producing Escherichia coli strains after repetitive exposure to far-UVC and blue LED light. J. Appl. Microbiol. 2023, 134, lxad124. [Google Scholar] [CrossRef]
- Jagger, J. Solar-UV Actions on Living Cells; Praeger: Westport, CT, USA, 1985. [Google Scholar]
- Knobling, B.; Franke, G.; Belmar Campos, C.; Büttner, H.; Christner, M.; Klupp, E.M.; Maurer, P.M.; Knobloch, J.K. Tolerance of clinical vancomycin-resistant Enterococcus faecium isolates against UV-C light from a mobile source. Antimicrob. Resist. Infect. Control 2023, 12, 63. [Google Scholar] [CrossRef]
- Ozawa, Y.; Tanimoto, K.; Fujimoto, S.; Tomita, H.; Ike, Y. Cloning and genetic analysis of the UV resistance determinant (uvr) encoded on the Enterococcus faecalis pheromone-responsive conjugative plasmid pAD1. J. Bacteriol. 1997, 179, 7468–7475. [Google Scholar] [CrossRef] [PubMed]
- Oni, R.A.; Sharma, M.; Buchanan, R.L. Survival of Salmonella enterica in dried turkey manure and persistence on spinach leaves. J. Food Prot. 2015, 78, 1791–1799. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Cheng, Y.; Moraru, C.I. Blue 405 nm LED light effectively inactivates bacterial pathogens on substrates and packaging materials used in food processing. Sci. Rep. 2023, 13, 15472. [Google Scholar] [CrossRef] [PubMed]
- Guffey, J.S.; Wilborn, J. In vitro bactericidal effects of 405-nm and 470-nm blue light. Photomed. Laser Surg. 2006, 24, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Hadi, J.; Wu, S.; Brightwell, G. Antimicrobial blue light versus pathogenic bacteria: Mechanism, application in the food industry, hurdle technologies and potential resistance. Foods 2020, 9, 1895. [Google Scholar] [CrossRef]
- García, A.H. Anhydrobiosis in bacteria: From physiology to applications. J. Biosci. 2011, 36, 939–950. [Google Scholar] [CrossRef]
- Gonzalez, J.M.; Aranda, B. Microbial growth under limiting conditions-future perspectives. Microorganisms 2023, 11, 1641. [Google Scholar] [CrossRef]
- Demain, A.L.; Fang, A. The natural functions of secondary metabolites. Adv. Biochem. Eng. Biotechnol. 2000, 69, 1–39. [Google Scholar] [CrossRef]
- Weiland-Bräuer, N. Friends or foes-microbial interactions in nature. Biology 2021, 10, 496. [Google Scholar] [CrossRef]
- Heejeong, L.; Dong Gun, L. Programmed cell death in bacterial community: Mechanisms of action, causes and consequences. J. Microbiol. Biotechnol. 2019, 29, 1014–1021. [Google Scholar] [CrossRef]
- Marteyn, B.; Scorza, F.B.; Sansonetti, P.J.; Tang, C. Breathing life into pathogens: The influence of oxygen on bacterial virulence and host responses in the gastrointestinal tract. Cell. Microbiol. 2011, 13, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Greffe, V.R.G.; Michiels, J. Desiccation-induced cell damage in bacteria and the relevance for inoculant production. Appl. Microbiol. Biotechnol. 2020, 104, 3757–3770. [Google Scholar] [CrossRef] [PubMed]
- Xian-Zhi, L.; Plésiat, P.; Nikaido, H. The challenge of efflux-mediated antibiotic resistance in gram-negative bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef]
- Buckhouse, J.C.; Gifford, G.F. Water quality implications of cattle grazing on a semiarid watershed in southeastern Utah. J. Range Manag. 1976, 29, 109–113. [Google Scholar] [CrossRef]
- Strøm, A.R.; Kaasen, I. Trehalose metabolism in Escherichia coli: Stress protection and stress regulation of gene expression. Mol. Microbiol. 1993, 8, 205–210. [Google Scholar] [CrossRef]
- Welsh, D.T.; Herbert, R.A. Osmotically induced intracellular trehalose, but not glycine betaine accumulation promotes desiccation tolerance in Escherichia coli. FEMS Microbiol. Lett. 1999, 174, 57–63. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, H.; Ionita, C.; Luo, F.; Jiang, X. Effects of chicken litter storage time and ammonia content on thermal resistance of desiccation-adapted Salmonella spp. Appl. Environ. Microbiol. 2015, 81, 6883–6889. [Google Scholar] [CrossRef]
- Lebreton, F.; Manson, A.L.; Saavedra, J.T.; Straub, T.J.; Earl, A.M.; Gilmore, M.S. Tracing the Enterococci from paleozoic origins to the hospital. Cell 2017, 169, 849–861.e813. [Google Scholar] [CrossRef]
- Bale, M.J.; Bennett, P.M.; Beringer, J.E.; Hinton, M. The survival of bacteria exposed to desiccation on surfaces associated with farm buildings. J. Appl. Bacteriol. 1993, 75, 519–528. [Google Scholar] [CrossRef]
- Whitman, W.B.; Rainey, F.; Kämpfer, P.; Trujillo, M.; Chun, J.; DeVos, P.; Hedlund, B.; Dedysh, S. Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley Online Library: Hoboken, NJ, USA, 2015; Volume 410. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure Time * | Time (Min) | Time (h) | UV-B Dose (J/m2) |
|---|---|---|---|
| 0 | 0 | 0 | 0 |
| 1 | 60 | 1 | 16,560 |
| 2 | 125 | 2.08 | 33,120 |
| 3 | 207 | 3.45 | 49,680 |
| Type of Sample | E. coli (%) | TET 1 Resistant E. coli (%) | AXO 2 Resistant E. coli (%) | Enterococcus spp. (%) | TET 1 Resistant Enterococcus spp. (%) | ERY 3 Resistant Enterococcus spp. (%) |
|---|---|---|---|---|---|---|
| UV-B | ||||||
| Dry manure (control) | 33.3 (3/9) | 11.1 (1/9) | 0 (0/9) | 100 (9/9) | 66.6 (6/9) | 88.9 (8/9) |
| Dry manure (treatment) | 77.8 (7/9) | 55.5 (5/9) | 0 (0/9) | 77.8 (7/9) | 66.7 (6/9) | 77.8 (7/9) |
| Fresh manure (control) | 100 (9/9) | 100 (9/9) | 100 (9/9) | 100 (9/9) | 100 (9/9) | 100 (9/9) |
| Fresh manure (treatment) | 100 (9/9) | 100 (9/9) | 100 (9/9) | 100 (9/9) | 100 (9/9) | 100 (9/9) |
| Relative Humidity | ||||||
| Control | 100 (12/12) | 100 (12/12) | 91.7(11/12) | 100 (12/12) | 100 (12/12) | 100 (12/12) |
| Treatment | 100 (12/12) | 100 (12/12) | 83.3 (10/12) | 100 (12/12) | 100 (12/12) | 100 (12/12) |
| Type of Sample | E. coli (Mean Log10) | TET 1 Resistant E. coli (Mean Log10) | AXO 2 Resistant E. coli (Mean Log10) | Enterococcus spp. (Mean Log10) | TET 1 Resistant Enterococcus spp. (Mean Log10) | ERY 3 Resistant Enterococcus spp. (Mean Log10) |
|---|---|---|---|---|---|---|
| UV-B | ||||||
| Dry manure (control) | 0.8 | 0.4 | 0 | 3.4 | 2.5 | 2.9 |
| Dry manure (treatment) | 2.1 | 1.6 | 0 | 2.9 | 1.9 | 2.1 |
| Fresh manure (control) | 5.7 | 4.9 | 4.3 | 5.3 | 3.5 | 3.5 |
| Fresh manure (treatment) | 5.9 | 5.4 | 4.2 | 5 | 3.2 | 3.2 |
| Relative Humidity | ||||||
| Control | 8.6 | 8.2 | 3.6 | 5.5 | 5.3 | 5.3 |
| Treatment | 8.5 | 7.9 | 3.1 | 5.9 | 5.2 | 5.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leon, I.M.; Auvermann, B.W.; Bush, K.J.; Casey, K.; Pinchak, W.E.; Vinasco, J.; Lawhon, S.D.; Smith, J.K.; Scott, H.M.; Norman, K.N. Assessing the Resilience of Enteric Bacteria in Manure in Response to Changes in Relative Humidity and UV-B Light. Environments 2024, 11, 197. https://doi.org/10.3390/environments11090197
Leon IM, Auvermann BW, Bush KJ, Casey K, Pinchak WE, Vinasco J, Lawhon SD, Smith JK, Scott HM, Norman KN. Assessing the Resilience of Enteric Bacteria in Manure in Response to Changes in Relative Humidity and UV-B Light. Environments. 2024; 11(9):197. https://doi.org/10.3390/environments11090197
Chicago/Turabian StyleLeon, Ingrid M., Brent W. Auvermann, Kevin Jack Bush, Kenneth Casey, William E. Pinchak, Javier Vinasco, Sara D. Lawhon, Jason K. Smith, Harvey Morgan Scott, and Keri N. Norman. 2024. "Assessing the Resilience of Enteric Bacteria in Manure in Response to Changes in Relative Humidity and UV-B Light" Environments 11, no. 9: 197. https://doi.org/10.3390/environments11090197